Combined Treatment with Omega-3 Fatty Acid and Cholecalciferol Increases 1,25-Dihydroxyvitamin D Levels by Modulating Dysregulation of Vitamin D Metabolism in 5/6 Nephrectomy Rats



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Histopathological Evaluation
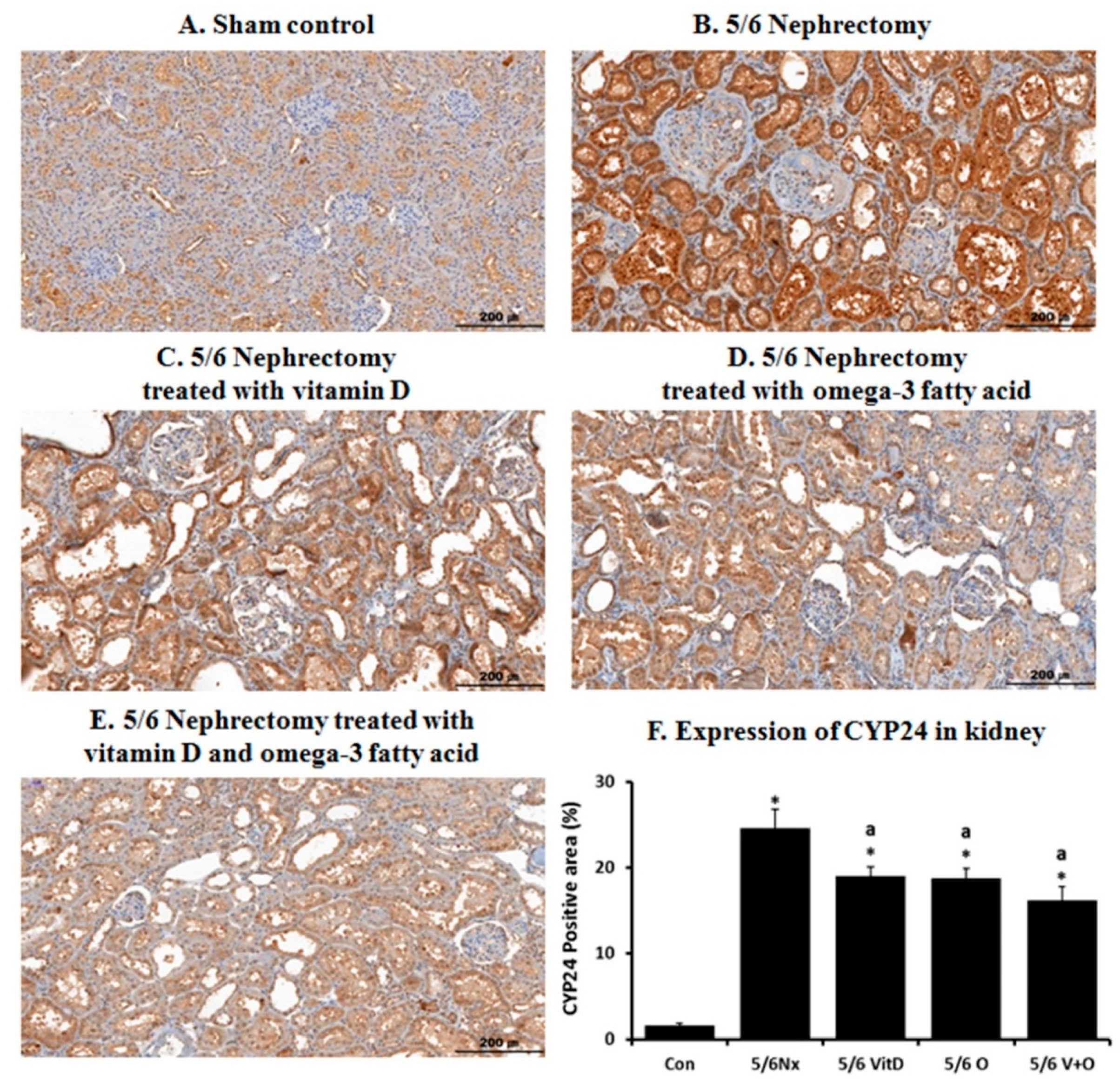
2.3. Immunohistochemistry
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
3.1. Characteristics of Animals
3.2. CYP24 Expression in the Kidney and Liver
3.3. CYP27B1 Expression in the Kidney and Liver
3.4. Renal Pathology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pilz, S.; Tomaschitz, A.; Friedl, C.; Amrein, K.; Drechsler, C.; Ritz, E.; Boehm, B.O.; Grammer, T.B.; Marz, W. Vitamin D status and mortality in chronic kidney disease. Nephrol. Dial. Transplant. 2011, 26, 3603–3609. [Google Scholar] [CrossRef] [PubMed]
- Ravani, P.; Malberti, F.; Tripepi, G.; Pecchini, P.; Cutrupi, S.; Pizzini, P.; Mallamaci, F.; Zoccali, C. Vitamin D levels and patient outcome in chronic kidney disease. Kidney Int. 2009, 75, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Helvig, C.F.; Cuerrier, D.; Hosfield, C.M.; Ireland, B.; Kharebov, A.Z.; Kim, J.W.; Ramjit, N.J.; Ryder, K.; Tabash, S.P.; Herzenberg, A.M.; et al. Dysregulation of renal vitamin D metabolism in the uremic rat. Kidney Int. 2010, 78, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Strugnell, S.A.; DeLuca, H.F. Current understanding of the molecular actions of vitamin D. Physiol. Rev. 1998, 78, 1193–1231. [Google Scholar] [CrossRef] [PubMed]
- Vuica, A.; Ferhatovic Hamzic, L.; Vukojevic, K.; Jeric, M.; Puljak, L.; Grkovic, I.; Filipovic, N. Aging and a long-term diabetes mellitus increase expression of 1 alpha-hydroxylase and vitamin D receptors in the rat liver. Exp. Gerontol. 2015, 72, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Gottfried, E.; Rehli, M.; Hahn, J.; Holler, E.; Andreesen, R.; Kreutz, M. Monocyte-derived cells express CYP27A1 and convert vitamin D3 into its active metabolite. Biochem. Biophys. Res. Commun. 2006, 349, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Vuica, A.; Vukojevic, K.; Ferhatovic Hamzic, L.; Jeric, M.; Puljak, L.; Grkovic, I.; Filipovic, N. Expression pattern of CYP24 in liver during ageing in long-term diabetes. Acta Histochem. 2016, 118, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Lambert, P.W.; Stern, P.H.; Avioli, R.C.; Brackett, N.C.; Turner, R.T.; Greene, A.; Fu, I.Y.; Bell, N.H. Evidence for extrarenal production of 1 alpha, 25-dihydroxyvitamin D in man. J. Clin. Investig. 1982, 69, 722–725. [Google Scholar] [CrossRef] [PubMed]
- An, W.S.; Lee, S.M.; Son, Y.K.; Kim, S.E.; Kim, K.H.; Han, J.Y.; Bae, H.R.; Rha, S.H.; Park, Y. Omega-3 fatty acid supplementation increases 1,25-dihydroxyvitamin D and fetuin-A levels in dialysis patients. Nutr. Res. 2012, 32, 495–502. [Google Scholar] [CrossRef] [PubMed]
- An, W.S.; Kim, H.J.; Cho, K.H.; Vaziri, N.D. Omega-3 fatty acid supplementation attenuates oxidative stress, inflammation, and tubulointerstitial fibrosis in the remnant kidney. Am. J. Physiol. Renal. Physiol. 2009, 297, F895–F903. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, D.; Bland, R.; Williams, M.C.; McNinch, R.W.; Howie, A.J.; Stewart, P.M.; Hewison, M. Extrarenal expression of 25-hydroxyvitamin d(3)-1 alpha-hydroxylase. J. Clin. Endocrinol. Metab. 2001, 86, 888–894. [Google Scholar] [PubMed]
- Hewison, M. Vitamin D and the immune system: New perspectives on an old theme. Rheum. Dis. Clin. 2012, 38, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Hewison, M. Extrarenal expression of the 25-hydroxyvitamin D-1-hydroxylase. Arch. Biochem. Biophys. 2012, 523, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Andress, D.L. Vitamin D in chronic kidney disease: A systemic role for selective vitamin D receptor activation. Kidney Int. 2006, 69, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Dusso, A.S.; Tokumoto, M. Defective renal maintenance of the vitamin D endocrine system impairs vitamin D renoprotection: A downward spiral in kidney disease. Kidney Int. 2011, 79, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, D.; Quinkler, M.; Eardley, K.S.; Bland, R.; Lepenies, J.; Hughes, S.V.; Raymond, N.T.; Howie, A.J.; Cockwell, P.; Stewart, P.M.; et al. Reduction of the vitamin D hormonal system in kidney disease is associated with increased renal inflammation. Kidney Int. 2008, 74, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Jean, G.; Terrat, J.C.; Vanel, T.; Hurot, J.M.; Lorriaux, C.; Mayor, B.; Chazot, C. Evidence for persistent vitamin D 1-alpha-hydroxylation in hemodialysis patients: Evolution of serum 1,25-dihydroxycholecalciferol after 6 months of 25-hydroxycholecalciferol treatment. Nephron Clin. Pract. 2008, 110, c58–c65. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Byford, V.; Arabian, A.; Sakai, Y.; Demay, M.B.; St-Arnaud, R.; Jones, G. Altered pharmacokinetics of 1alpha,25-dihydroxyvitamin D3 and 25-hydroxyvitamin D3 in the blood and tissues of the 25-hydroxyvitamin D-24-hydroxylase (Cyp24a1) null mouse. Endocrinology 2005, 146, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 6) | 5/6 Nephrectomy (n = 6) | 5/6 Nephrectomy with Cholecalciferol (n = 6) | 5/6 Nephrectomy with Omega-3 FA (n = 6) | 5/6 Nephrectomy with Cholecalciferol and Omega-3 FA (n = 6) | p Value | |
|---|---|---|---|---|---|---|
| Final body weight (g) | 464 ± 9 | 385 ± 11 * | 420 ± 33 * | 422 ± 10 *a | 422 ± 12 *a | <0.001 |
| Body weight gain (g) | 91 ± 3 | 46 ± 13 * | 71 ± 26 | 75 ± 4 a | 73 ± 6 a | <0.001 |
| Blood urea nitrogen (mg/dL) | 18 ± 0.7 | 77 ± 35 * | 72 ± 23 * | 68 ± 19 | 55 ± 13 ab | 0.003 |
| Creatinine (mg/dL) | 0.4 ± 0.0 | 1.3 ± 0.6 * | 1.2 ± 0.3 * | 1.1 ± 0.3 | 0.9 ± 0.2 abc | 0.002 |
| Calcium (mg/dL) | 7 ± 0.4 | 7 ± 1 | 6 ± 0.4 | 8 ± 0.6 | 7 ± 1 | 0.50 |
| Phosphorus (mg/dL) | 8 ± 0.5 | 11 ± 5 | 8 ± 1 | 8 ± 0.6 | 8 ± 0.6 | 0.26 |
| 25(OH)D (ng/mL) | 98 ± 6 | 28 ± 17 * | 50 ± 42 | 62 ± 39 a | 111 ± 37 abc | 0.003 |
| 1,25(OH)2D (pg/mL) | 171 ± 41 | 45 ± 17* | 58 ± 41 * | 83 ± 42 * | 108 ± 43 *ab | 0.002 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.M.; Lee, M.H.; Son, Y.K.; Kim, S.E.; An, W.S. Combined Treatment with Omega-3 Fatty Acid and Cholecalciferol Increases 1,25-Dihydroxyvitamin D Levels by Modulating Dysregulation of Vitamin D Metabolism in 5/6 Nephrectomy Rats. Nutrients 2019, 11, 2903. https://doi.org/10.3390/nu11122903
Lee SM, Lee MH, Son YK, Kim SE, An WS. Combined Treatment with Omega-3 Fatty Acid and Cholecalciferol Increases 1,25-Dihydroxyvitamin D Levels by Modulating Dysregulation of Vitamin D Metabolism in 5/6 Nephrectomy Rats. Nutrients. 2019; 11(12):2903. https://doi.org/10.3390/nu11122903
Chicago/Turabian StyleLee, Su Mi, Mi Hwa Lee, Young Ki Son, Seong Eun Kim, and Won Suk An. 2019. "Combined Treatment with Omega-3 Fatty Acid and Cholecalciferol Increases 1,25-Dihydroxyvitamin D Levels by Modulating Dysregulation of Vitamin D Metabolism in 5/6 Nephrectomy Rats" Nutrients 11, no. 12: 2903. https://doi.org/10.3390/nu11122903
APA StyleLee, S. M., Lee, M. H., Son, Y. K., Kim, S. E., & An, W. S. (2019). Combined Treatment with Omega-3 Fatty Acid and Cholecalciferol Increases 1,25-Dihydroxyvitamin D Levels by Modulating Dysregulation of Vitamin D Metabolism in 5/6 Nephrectomy Rats. Nutrients, 11(12), 2903. https://doi.org/10.3390/nu11122903