Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Blood and Organ Sampling
2.3. Biochemical Plasma Analyses
2.4. Fatty Acid Composition Analyses
2.5. Hepatic Lipid Measurements
2.6. Statistical Analysis
3. Results
3.1. Effects of P. tricornutum on Body and Organ Weight
3.1.1. Food and Water Intake
3.1.2. Energy Intake
3.1.3. Body and Organ Weights
3.2. Effects of P. tricornutum on Fatty Acid Composition of Plasma, RBC and Liver Lipids
3.2.1. SFA Contents
3.2.2. n-6 LC-PUFA Contents
3.2.3. n-3 LC-PUFA contents
3.2.4. MUFA Levels
3.2.5. Δ9-Desaturase Index
3.3. Effects of P. tricornutum Supplementation on Physiological and Metabolic Disorders in Wistar Rats Fed a High-Fat Diet
3.3.1. Plasma Lipid Levels and AIP Index
3.3.2. Transaminase and Hepatic Lipid Levels
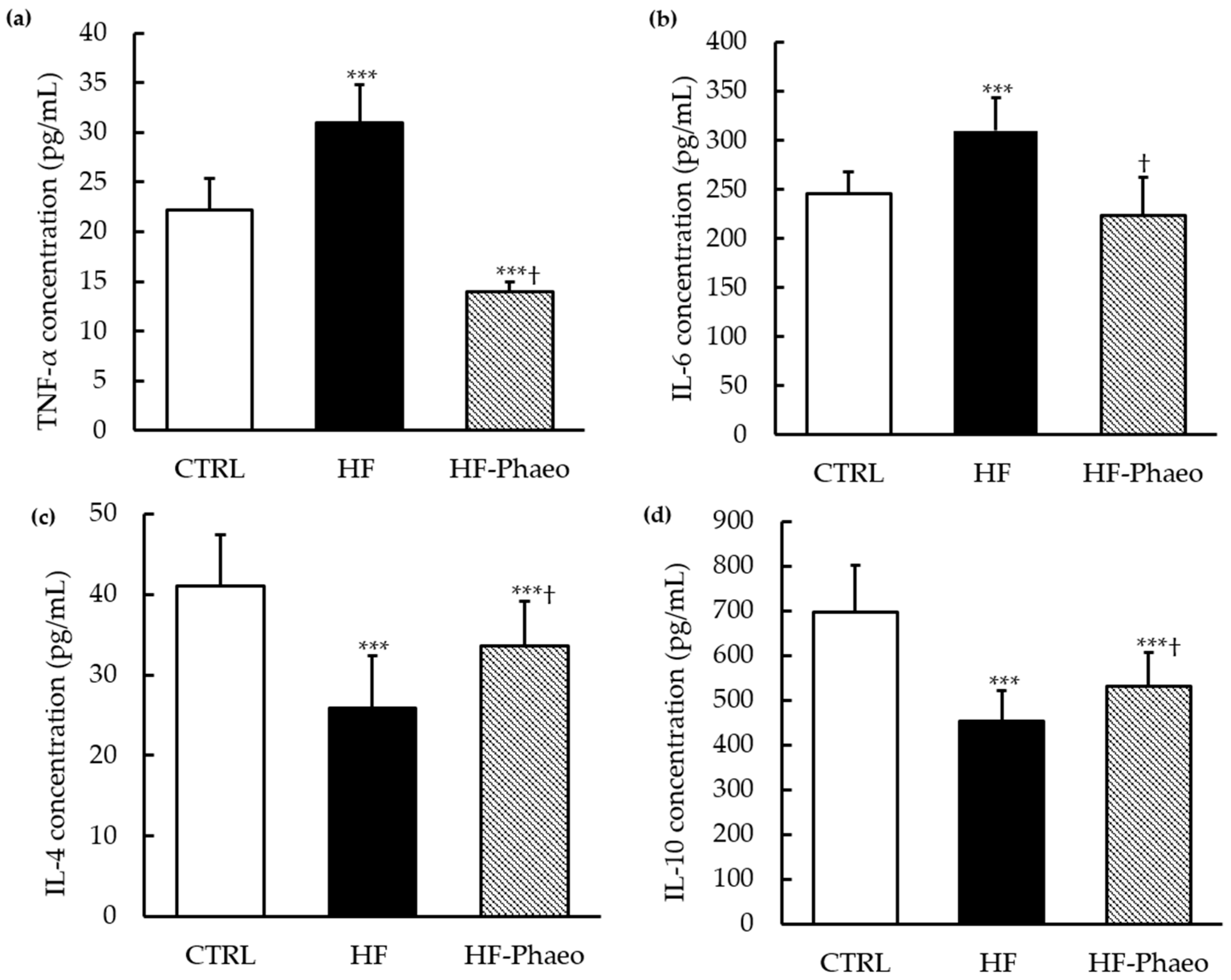
3.4. Effects of P. tricornutum on Inflammatory Status
3.4.1. Pro-Inflammatory Cytokines
3.4.2. Anti-Inflammatory Cytokines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Punthakee, Z.; Goldenberg, R.; Katz, P.; Diabetes Canada Clinical Practice Guidelines Expert Committee. Definition, Classification and Diagnosis of Diabetes, Prediabetes and Metabolic Syndrome. Can. J. Diabetes 2018, 42, S10–S15. [Google Scholar] [CrossRef]
- Bessone, F.; Razori, M.V.; Roma, M.G. Molecular pathways of nonalcoholic fatty liver disease development and progression. Cel. Mol. Life Sci. 2009, 76, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Soares, R. Chronic Inflammation in the Metabolic Syndrome: Emphasis on Adipose tissue. In Oxidative Stress, Inflammation and Angiogenesis in the Metabolic Syndrome; Soares, R., Costa, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 65–84. [Google Scholar] [CrossRef]
- Yanai, H.; Masui, Y.; Katsuyama, H.; Adachi, H.; Kawaguchi, A.; Hakoshima, M.; Waragai, Y.; Harigae, T.; Sako, A. An Improvement of Cardiovascular Risk Factors by Omega-3 Polyunsaturated Fatty Acids. J. Clin. Med. Res. 2018, 10, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Adkins, Y.; Kelley, D.S. Mechanisms underlying the cardioprotective effects of omega-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2010, 21, S781–S792. [Google Scholar] [CrossRef]
- De la Iglesia, R.; Loria-Kohen, V.; Zulet, M.; Martinez, J.; Reglero, G.; Ramirez de Molina, A. Dietary Strategies Implicated in the Prevention and Treatment of Metabolic Syndrome. Int. J. Mol. Sci. 2016, 17, 1877. [Google Scholar] [CrossRef]
- Martins, D.; Custódio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K. Alternative Sources of n-3 Long-Chain Polyunsaturated Fatty Acids in Marine Microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.P.; Morant-Manceau, A.; Schoefs, B. The potential of microalgae for the production of bioactive molecules of pharmaceutical interest. Curr. Pharm. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Goiris, K.; Muylaert, K.; Foubert, I. Nutritional evaluation of microalgae oils rich in omega-3 long chain polyunsaturated fatty acids as an alternative for fish oil. Food Chem. 2014, 160, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazih, H.; Bard, J.M. Microalgae in Human Health: Interest as a Functional Food. In Microalgae in Health and Disease Prevention; Levine, I.A., Fleurence, J., Eds.; Academic Press: Cambridge, UK, 2018; pp. 211–226. [Google Scholar] [CrossRef]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion 500 nutraceuticals. Microb Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef]
- Rebolloso-Fuentes, M.M.; Navarro-Pérez, A.; Ramos-Miras, J.J.; Uil-Guerrero, J.L. Biomass nutrient profiles of the microalga phaeodactylum tricornutum. J. Food Biochem. 2007, 25, 57–76. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.M.; Cha, K.H.; Mok, I.K.; Koo, S.Y.; Pan, C.H.; Lee, J.K. Evaluation of the anti-obesity effect of the microalga Phaeodactylum tricornutum. Appl. Biol. Chem. 2016, 59, 283–290. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Fairus, A.; Ima Nirwana, S. Animal models of metabolic syndrome: A review. Nutr. Metab. 2016, 13, 65. [Google Scholar] [CrossRef]
- Aydin, S.; Aksoy, A.; Aydin, S.; Kalayci, M.; Yilmaz, M.; Kuloglu, T.; Citil, C.; Catak, Z. Today’s and yesterday’s of pathophysiology: Biochemistry of metabolic syndrome and animal models. Nutrients 2014, 30, 1–9. [Google Scholar] [CrossRef]
- Moreno-Fernández, S.; Garcés-Rimón, M.; Vera, G.; Astier, J.; Landrier, J.; Miguel, M. High Fat/High Glucose Diet Induces Metabolic Syndrome in an Experimental Rat Model. Nutrients 2018, 10, 1502. [Google Scholar] [CrossRef]
- Panchal, S.K.; Brown, L. Rodent Models for Metabolic Syndrome Research. J. Biomed. Biotechnol. 2011, 2011, 1–14. [Google Scholar] [CrossRef]
- Jensen, T.; Abdelmalek, M.F.; Sullivan, S.; Nadeau, K.J.; Green, M.; Roncal, C.; Nakagawa, T.; Kuwabara, M.; Sato, Y.; Kang, D.H.; et al. Fructose and sugar: A major mediator of non-alcoholic fatty liver disease. J. Hepatol. 2018, 68, 1063–1075. [Google Scholar] [CrossRef]
- Haimeur, A.; Mimouni, V.; Ulmann, L.; Martineau, A.S.; Messaouri, H.; Pineau-Vincent, F.; Tremblin, G.; Meskini, N. Fish Oil and Microalga Omega-3 as Dietary Supplements: A Comparative Study on Cardiovascular Risk Factors in High fat Fed Rats. Lipids 2016, 51, 1037–1049. [Google Scholar] [CrossRef]
- Mimouni, V.; Ulmann, L.; Haimeur, A.; Guéno, F.; Meskini, N.; Tremblin, G. Marine microalgae used as food supplements and their implication in preventing cardiovascular diseases. OCL 2015, 22, D409. [Google Scholar] [CrossRef]
- Toop, C.; Gentili, S. Fructose Beverage Consumption Induces a Metabolic Syndrome Phenotype in the Rat: A Systematic Review and Meta-Analysis. Nutrients 2016, 8, 577. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Marsh, J.B.; Weinstein, D.B. Simple charring method for determination of lipids. J. Lipid Res. 1966, 4, 574–576. [Google Scholar]
- Simionato, J.I.; Garcia, J.C.; dos Santos, G.T.; Oliveira, C.C.; Visentainer, J.V.; de Souza, N.E. Validation of the determination of fatty acids in milk by gas chromatography. J. Braz. Chem. Soc. 2010, 21, 520–524. [Google Scholar] [CrossRef] [Green Version]
- Vohra, D.F. Determination of photosynthetic pigments in sea-water. In Monographs Onocéanographie Methodology; UNESCO, Ed.; UNESCO: Paris, France, 1966; p. 66. [Google Scholar]
- Van Heukelem, L.; Thomas, C.S. Computer-assisted high-performance liquid chromatography method development with applications to the isolation and analysis of phytoplankton pigments. J. Chromatogr. A 2001, 910, 31–49. [Google Scholar] [CrossRef]
- Boisen, S.; Fernández, J.A. Prediction of the total tract digestibility of energy in feedstuffs and pig diets by in vitro analyses. Anim. Feed Sci. Technol. 1997, 68, 277–286. [Google Scholar] [CrossRef]
- Batista, A.P.; Niccolai, A.; Fradinho, P.; Fragoso, S.; Bursic, I.; Rodolfi, L.; Biondi, N.; Tredici, M.R.; Sousa, I.; Raymundo, A. Microalgae biomass as an alternative ingredient in cookies: Sensory, physical and chemical properties, antioxidant activity and in vitro digestibility. Algal Res. 2017, 26, 161–171. [Google Scholar] [CrossRef]
- Bondet, V.; Brand Williams, W.; Berset, C. Kinetics and Mechanisms of Antioxidant Activity using the DPPH. Free Radical Method. LWT Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Frohlich, J. Fractional Esterification Rate of Cholesterol and Ratio of Triglycerides to HDL-Cholesterol Are Powerful Predictors of Positive Findings on Coronary Angiography. Clin. Chem. 2003, 49, 1873–1880. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Delsal, J.L.; Lees, M.; Sloane Stanley, G. New method of extraction of serum lipids by methylal. Application to micro-estimation of total cholesterol, phospho-aminolipins and proteins. Bull. Soc. Chim Biol. 1944, 26, 99. [Google Scholar]
- Slover, H.T.; Lanza, E. Quantitative analysis of food fatty acids by capillary gas chromatography. J. Am. Oil Chem. Soc. 1979, 56, 933. [Google Scholar] [CrossRef]
- Buettner, R.; Schölmerich, J.; Bollheimer, L.C. High fat Diets: Modeling the Metabolic Disorders of Human Obesity in Rodents. Obesity 2012, 15, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Go, R.E.; Hwang, K.A.; Park, G.T.; Lee, H.M.; Lee, G.A.; Kim, C.W.; Jeon, S.Y.; Seo, J.W.; Hong, W.K.; Choi, K.C. Effects of microalgal polyunsaturated fatty acid oil on body weight and lipid accumulation in the liver of C57BL/6 mice fed a high fat diet. J. Biomed. Res. 2016, 30, 234–242. [Google Scholar] [CrossRef]
- Bertrand, C.; Pignalosa, A.; Wanecq, E.; Rancoule, C.; Batut, A.; Deleruyelle, S.; Lionetti, L.; Valet, P.; Castan-Laurell, I. Effects of Dietary Eicosapentaenoic Acid (EPA) Supplementation in High fat Fed Mice on Lipid Metabolism and Apelin/APJ System in Skeletal Muscle. PLoS ONE 2013, 8, e78874. [Google Scholar] [CrossRef] [PubMed]
- Ruzickova, J.; Rossmeisl, M.; Prazak, T.; Flachs, P.; Sponarova, J.; Veck, M.; Tvrzicka, E.; Bryhn, M.; Kopecky, J. Omega-3 PUFA of marine origin limit diet-induced obesity in mice by reducing cellularity of adipose tissue. Lipids 2004, 39, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Pérez Matute, P.; Pérez Echarri, N.; Martínez, J.A.; Marti, A.; Moreno Aliaga, M.J. Eicosapentaenoic acid actions on adiposity and insulin resistance in control and high-fat-fed rats: Role of apoptosis, adiponectin and tumour necrosis factor-α. Br. J. Nutr. 2007, 97, 389–398. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [Green Version]
- Grasa López, A.; Miliar García, Á.; Quevedo Corona, L.; Paniagua Castro, N.; Escalona Cardoso, G.; Reyes Maldonado, E.; Jaramillo-Flores, M.E. Undaria pinnatifida and Fucoxanthin Ameliorate Lipogenesis and Markers of Both Inflammation and Cardiovascular Dysfunction in an Animal Model of Diet-Induced Obesity. Mar. Drugs 2007, 14, 148. [Google Scholar] [CrossRef]
- Slavin, J.L. Dietary fiber and body weight. Nutrition 2005, 21, 411–418. [Google Scholar] [CrossRef]
- Nomura, K.; Yamanouchi, T. The role of fructose-enriched diets in mechanisms of nonalcoholic fatty liver disease. J. Nutr. Biochem. 2012, 23, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Comte, C.; Bellenger, S.; Bellenger, J.; Tessier, C.; Poisson, J.P.; Narce, M. Effects of streptozotocin and dietary fructose on delta-6 desaturation in spontaneously hypertensive rat liver. Biochimie 2004, 86, 799–806. [Google Scholar] [CrossRef]
- Mašek, T.; Filipović, N.; Vuica, A.; Starčević, K. Effects of treatment with sucrose in drinking water on liver histology, lipogenesis and lipogenic gene expression in rats fed high-fiber diet. Prostaglandins Leukot. Essent. Fat. Acids 2017, 116, 1–8. [Google Scholar] [CrossRef]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n- 3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, S1467–S1476. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Sinclair, A.J.; Kaur, G.; Li, D. Differential effects of EPA, DPA and DHA on cardio-metabolic risk factors in high fat diet fed mice. Prostaglandins Leukot. Essent. Fat. Acids PLEFA 2018, 136, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Lamaziere, A.; Wolf, C.; Barbe, U.; Bausero, P.; Visioli, F. Lipidomics of hepatic lipogenesis inhibition by omega 3 fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the Metabolic Syndrome: A Joint Interim Statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Ostlund, R.E. Phytosterols in human nutrition. Annu. Rev. Nutr. 2002, 22, 533–549. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W.; Jones, A.E.; Riddell Mason, S. Ten Different Dietary Fibers Have Significantly Different Effects on Serum and Liver Lipids of Cholesterol-Fed Rats. J. Nutr. 2002, 124, 78–83. [Google Scholar] [CrossRef]
- Egert, S.; Kannenberg, F.; Somoza, V.; Erbersdobler, H.F.; Wahrburg, U. Dietary α-Linolenic Acid, EPA, and DHA Have Differential Effects on LDL Fatty Acid Composition but Similar Effects on Serum Lipid Profiles in Normolipidemic Humans. J. Nutr. 2009, 139, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S.M. Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats. Nutrients 2009, 1, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Threapleton, D.E.; Greenwood, D.C.; Evans, C.E.L.; Cleghorn, C.L.; Nykjaer, C.; Woodhead, C.; Cade, J.E.; Gale, C.P.; Burley, V.J. Dietary fibre intake and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2013, 347, F6879. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Sano, K.; Horiuchi, A.; Tanaka, E.; Kiyosawa, K.; Aoyama, T. Highly purified eicosapentaenoic acid treatment improves nonalcoholic steatohepatitis. J. Clin. Gastroenterol. 2008, 42, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Adhyaru, B.B.; Jacobson, T.A. New Cholesterol Guidelines for the Management of Atherosclerotic Cardiovascular Disease Risk. Endocrinol. Metab. Clin. 2016, 45, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.W.; Lu, Y.; Li, F.; Yang, C.J.; Feng, Y.B.; Li, H.W.; Yao, W.F.; Shen, Z.H. Atherogenic index of plasma is an effective index for estimating abdominal obesity. Lipids Health Dis. 2018, 17, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Liu, J.R.; Gao, J.M.; Parry, J.W.; Wei, Y.M. Antioxidant Activity of Tartary Buckwheat Bran Extract and Its Effect on the Lipid Profile of Hyperlipidemic Rats. J. Agric. Food Chem. 2009, 57, 5106–5112. [Google Scholar] [CrossRef]
- Luo, X.; Su, P.; Zhang, W. Advances in Microalgae-Derived Phytosterols for Functional Food and Pharmaceutical Applications. Mar. Drugs 2015, 13, 4231–4254. [Google Scholar] [CrossRef] [Green Version]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of polysaccharides in food, digestion, and health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef]
- Cao, J.; Schwichtenberg, K.A.; Hanson, N.Q.; Tsai, M.Y. Incorporation and Clearance of Omega-3 Fatty Acids in Erythrocyte Membranes and Plasma Phospholipids. Clin. Chem. 2006, 52, 2265–2272. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.J.; Meyer, B.J.; Mori, T.A.; Burke, V.; Mansour, J.; Patch, C.S.; Tapsell, L.C.; Noakes, M.; Clifton, P.A.; Barden, A.; et al. Impact of foods enriched with n-3 long-chain polyunsaturated fatty acids on erythrocyte n-3 levels and cardiovascular risk factors. Br. J. Nutr. 2007, 97, 598–749. [Google Scholar] [CrossRef]
- Howe, P.; Buckley, J.; Murphy, K.; Pettman, T.; Milte, C.; Coates, A. Relationship between Erythrocyte Omega-3 Content and Obesity Is Gender Dependent. Nutrients 2014, 6, 1850–1860. [Google Scholar] [CrossRef] [Green Version]
- De Mello-Sampayo, C.; Paterna, A.; Polizzi, A.; Duarte, D.; Batista, I.; Pinto, R.; Gonçalves, P.; Raymundo, A.; Batista, A.; Gouveia, L.; et al. Evaluation of marine microalga Diacronema vlkianum biomass fatty acid assimilation in Wistar rats. Molecules 2017, 22, 1097. [Google Scholar] [CrossRef]
- Levy, J.R.; Clore, J.N.; Stevens, W. Dietary n-3 polyunsaturated fatty acids decrease hepatic triglycerides in Fischer 344 rats. Hepatology 2004, 39, 608–616. [Google Scholar] [CrossRef] [Green Version]
- Dupas, J.; Goanvec, C.; Feray, A.; Guernec, A.; Alain, C.; Guerrero, F.; Mansourati, J. Progressive Induction of Type 2 Diabetes: Effects of a Reality-Like Fructose Enriched Diet in Young Wistar Rats. PLoS ONE 2016, 11, e0146821. [Google Scholar] [CrossRef]
- Amine, H.; Benomar, Y.; Haimeur, A.; Messaouri, H.; Meskini, N.; Taouis, M. Odontella aurita-enriched diet prevents high fat diet induced liver insulin resistance. J. Endocrinol. JOE 2015, 228, 1–12. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Nah, J.W.; Jeon, Y.J. Potential anti-inflammatory natural products from marine algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef]
- Tan, C.; Hou, Y. First Evidence for the Anti-inflammatory Activity of Fucoxanthin in High fat-Diet-Induced Obesity in Mice and the Antioxidant Functions in PC12 Cells. Inflammation 2014, 37, 443–450. [Google Scholar] [CrossRef]
- Calder, P.C. Dietary modification of inflammation with lipids. Proc. Nutr. Soc. 2002, 61, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Robertson, R.; Guihéneuf, F.; Bahar, B.; Schmid, M.; Stengel, D.; Fitzgerald, G.; Ross, R.; Stanton, C. The Anti-Inflammatory Effect of Algae-Derived Lipid Extracts on Lipopolysaccharide (LPS)-Stimulated Human THP-1 Macrophages. Mar. Drugs 2015, 13, 5402–5424. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos Moraes, A.; Pisani, L.P.; Corgosinho, F.C.; Testa Carvalho, L.O.; Masquio, D.C.L.; Jamar, G.; Sanches, R.B.; Oyama, L.M.; Dâmaso, A.R.; Belote, C.; et al. The Role of Leptinemia State as a Mediator of Inflammation in Obese Adults. Horm. Metab. Res. 2013, 45, 605–610. [Google Scholar] [CrossRef]
- Yook, J.S.; Kim, K.A.; Park, J.E.; Lee, S.H.; Cha, Y.S. Microalgal Oil Supplementation Has an Anti-Obesity Effect in C57BL/6J Mice Fed a High Fat Diet. Prev. Nutr. Food Sci. 2015, 20, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.J.; Kim, S.M.; Jeong, S.M.; Choi, H.N.; Jang, Y.H.; Kim, J.I. Antioxidant effect of Phaeodactylum tricornutum in mice fed high fat diet. Food Sci. Biotechnol. 2013, 22, 107–113. [Google Scholar] [CrossRef]
- Nicoletti, M. Microalgae Nutraceuticals. Foods 2016, 5, 54. [Google Scholar] [CrossRef]
- Gwaltney Brant, S.M. Nutraceuticals in Hepatic Diseases. In Nutraceuticals; Ramesh, C., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 87–99. [Google Scholar] [CrossRef]
- Ibitoye, O.B.; Ajiboye, T.O. Dietary phenolic acids reverse insulin resistance, hyperglycaemia, dyslipidaemia, inflammation and oxidative stress in high-fructose diet-induced metabolic syndrome rats. Arch. Physiol. Biochem. 2018, 124, 410–417. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CTRL | HF | HF-Phaeo | ||||
|---|---|---|---|---|---|---|
| Parameter | Mean | SD | Mean | SD | Mean | SD |
| Final BW at day 56 of treatment (g) | 457.86 | 25.32 | 512.75 ** | 28.82 | 468.75 † | 38.37 |
| LW/BW (%) | 2.57 | 0.20 | 2.76 * | 0.16 | 2.62 | 0.13 |
| AAT/BW (%) | 1.61 | 0.27 | 4.12 *** | 0.32 | 2.26 ***† | 0.51 |
| EAT/BW (%) | 1.63 | 0.20 | 2.76 *** | 0.14 | 1.86 | 0.36 |
| CTRL | HF | HF-Phaeo | ||||
|---|---|---|---|---|---|---|
| Fatty Acids (mol %) | Mean | SD | Mean | SD | Mean | SD |
| SFA | ||||||
| 14:0 | 0.47 | 0.05 | 1.35 ** | 0.31 | 0.88 **† | 0.18 |
| 16:0 | 22.07 | 2.95 | 22.21 | 1.84 | 17.51 *† | 2.27 |
| 18:0 | 9.52 | 1.76 | 11.51 | 0.60 | 10.68 | 1.80 |
| Total SFA | 28.97 | 1.89 | 35.07 | 3.90 | 29.07 | 4.21 |
| MUFA | ||||||
| 16:1 | 2.27 | 0.28 | 2.10 | 0.60 | 1.86 | 0.24 |
| 18:1 (n-7 + n-9) | 13.33 | 1.75 | 18.80 ** | 3.41 | 12.91 † | 0.30 |
| 20:1n-9 | ND | - | ND | - | ND | - |
| 22:1 | ND | - | ND | - | ND | - |
| 24:1n-9 | ND | - | ND | - | ND | - |
| Total MUFA | 15.60 | 2.01 | 20.90 * | 3.96 | 14.90 † | 0.52 |
| PUFA | ||||||
| 18:2n-6 | 16.44 | 1.32 | 10.69 *** | 1.42 | 10.38 *** | 0.91 |
| 20:2n-6 | 0.44 | 0.11 | 1.26 ** | 0.18 | 0.74 † | 0.23 |
| 20:4n-6 | 22.18 | 1.56 | 17.20 ** | 2.89 | 14.69 ** | 3.26 |
| n-6 PUFA | 38.70 | 0.45 | 26.84 ** | 2.86 | 25.81 ** | 4.03 |
| 18:3n-3 | 0.47 | 0.12 | 0.31 ** | 0.03 | 0.26 ** | 0.03 |
| 20:5n-3 | 0.49 | 0.15 | 0.48 | 0.18 | 3.49 *** † | 1.03 |
| 22:5n-3 | 0.72 | 0.23 | 0.60 | 0.17 | 1.66 **† | 0.38 |
| 22:6n-3 | 3.93 | 0.62 | 4.39 | 0.32 | 4.64 | 1.24 |
| n-3 PUFA | 5.89 | 0.69 | 5.87 | 0.69 | 9.18 *† | 1.98 |
| n-6 PUFA/n-3 PUFA | 6.26 | 0.37 | 4.86 *** | 0.02 | 2.74 ***† | 0.43 |
| Total PUFA | 44.89 | 0.66 | 35.35 * | 3.39 | 40.22 † | 3.29 |
| MUFA/SFA | 0.55 | 0.10 | 0.61 | 0.17 | 0.50 | 0.08 |
| Δ9-Desaturase index ‡ | 0.58 | 0.07 | 0.62 | 0.09 | 0.54 | 0.05 |
| CTRL | HF | HF-Phaeo | ||||
|---|---|---|---|---|---|---|
| Fatty Acids (mol %) | Mean | SD | Mean | SD | Mean | SD |
| SFA | ||||||
| 14:0 | 0.80 | 0.22 | 0.90 | 0.27 | 1.13 | 0.26 |
| 16:0 | 40.24 | 2.68 | 28.79 ** | 2.61 | 34.12 **† | 5.04 |
| 18:0 | 14.86 | 2.64 | 13.90 | 1.58 | 15.03 | 1.69 |
| Total SFA | 55.91 | 3.90 | 43.41 ** | 2.02 | 50.27 † | 6.19 |
| MUFA | ||||||
| 16:1 | 1.03 | 0.23 | 0.96 | 0.28 | 1.09 | 0.17 |
| 18:1 (n-7 + n-9) | 11.64 | 1.30 | 14.01 * | 2.03 | 11.94 | 0.59 |
| 20:1n-9 | 0.30 | 0.015 | 0.19 *** | 0.04 | 0.12 ***† | 0.02 |
| 22:1 | 0.76 | 0.06 | 0.97 | 0.25 | 0.24 ***† | 0.09 |
| 24:1n-9 | ND | - | ND | - | ND- | |
| Total MUFA | 13.76 | 2.08 | 15.61 | 2.42 | 12.88 | 0.15 |
| PUFA | ||||||
| 18:2n-6 | 6.69 | 0.55 | 7.69 | 1.04 | 6.05 † | 1.00 |
| 20:2n-6 | 0.37 | 0.02 | 0.84 ** | 0.17 | 0.48 † | 0.15 |
| 20:4n-6 | 10.48 | 1.83 | 17.05 ** | 3.45 | 8.81 | 3.09 |
| n-6 PUFA | 17.54 | 1.51 | 25.63 *** | 3.50 | 13.74 ***† | 1.92 |
| 18:3n-3 | ND | - | ND | - | ND- | |
| 20:5n-3 | 0.19 | 0.01 | 0.35 | 0.13 | 1.69 ***† | 0.61 |
| 22:5n-3 | 3.28 | 0.87 | 2.83 | 0.46 | 11.39 ***† | 2.21 |
| 22:6n-3 | 1.17 | 0.12 | 2.43 *** | 0.41 | 1.59 † | 0.68 |
| n-3 PUFA | 10.38 | 1.91 | 5.79 *** | 0.48 | 14.68 ***† | 2.44 |
| n-6 PUFA/n-3 PUFA | 1.62 | 0.30 | 4.50 *** | 0.62 | 1.12 † | 0.08 |
| Total PUFA | 26.85 | 2.21 | 31.81 | 4.44 | 30.02 | 5.84 |
| MUFA/SFA | 0.25 | 0.05 | 0.36 * | 0.04 | 0.25 † | 0.00 |
| Δ9-Desaturase index ‡ | 0.45 | 0.06 | 0.50 | 0.06 | 0.44 | 0.03 |
| CTRL | HF | HF-Phaeo | ||||
|---|---|---|---|---|---|---|
| Fatty Acid (mol %) | Mean | SD | Mean | SD | Mean | SD |
| SFA | ||||||
| 14:0 | 0.18 | 0.05 | 0.41 *** | 0.05 | 0.32 ***† | 0.04 |
| 16:0 | 20.94 | 1.08 | 18.91 * | 1.48 | 18.45 * | 1.16 |
| 18:0 | 17.41 | 2.21 | 19.58 | 1.01 | 21.95 * | 2.47 |
| Total SFA | 38.53 | 1.83 | 38.90 | 1.74 | 40.72 | 1.55 |
| MUFA | ||||||
| 16:1 | 1.09 | 0.39 | 0.84 | 0.19 | 0.78 | 0.23 |
| 18:1(n-7 + n-9) | 7.74 | 1.43 | 9.42 * | 0.68 | 7.29 † | 0.72 |
| 20:1n-9 | 0.22 | 0.06 | 0.14 | 0.07 | 0.12 * | 0.06 |
| 22:1 | 0.26 | 0.05 | 0.39 | 0.10 | 0.20 † | 0.16 |
| 24:1n-9 | ND | - | ND | - | ND | - |
| Total MUFA | 9.27 | 1.88 | 10.72 | 0.99 | 8.38 † | 1.02 |
| PUFA | ||||||
| 18:2n-6 | 11.15 | 1.08 | 7.76 ** | 1.49 | 7.89 ** | 1.12 |
| 20:2n-6 | ND | - | ND | - | ND | - |
| 20:4n-6 | 28.07 | 2.29 | 26.44 | 2.04 | 24.16 * | 1.38 |
| n-6 PUFA | 39.22 | 2.06 | 34.21 *** | 1.47 | 32.05 *** | 1.65 |
| 18:3n-3 | ND | - | ND | - | ND | - |
| 20:5n-3 | 0.19 | 0.05 | 0.42 | 0.10 | 2.29 ***† | 0.23 |
| 22:5n-3 | 0.90 | 0.11 | 0.98 | 0.17 | 2.43 ***† | 0.28 |
| 22:6n-3 | 8.78 | 1.11 | 10.00 | 1.01 | 9.99 | 0.72 |
| n-3 PUFA | 9.88 | 1.25 | 11.31 | 0.82 | 14.72 ***† | 1.03 |
| n-6 PUFA/n-3 PUFA | 4.04 | 0.72 | 3.04 *** | 0.28 | 2.19 ***† | 0.25 |
| Total PUFA | 49.10 | 1.17 | 45.52 ** | 1.46 | 46.76 ** | 1.09 |
| MUFA/SFA | 0.24 | 0.06 | 0.28 | 0.04 | 0.21 † | 0.03 |
| Δ9-Desaturase index ‡ | 0.31 | 0.07 | 0.32 | 0.02 | 0.25 † | 0.04 |
| CTRL | HF | HF-Phaeo | ||||
|---|---|---|---|---|---|---|
| Parameter | Mean | SD | Mean | SD | Mean | SD |
| Plasma biochemical parameters | ||||||
| ASAT (UI/L) | 61.74 | 4.25 | 50.97 *** | 5.71 | 52.75 *** | 4.27 |
| ALAT (UI/L) | 41.52 | 5.16 | 50.30 *** | 4.91 | 47.07 *** | 6.49 |
| ASAT/ALAT ratio | 1.55 | 0.20 | 1.12 *** | 0.11 | 1.09 *** | 0.17 |
| Glucose (mmol/L) | 8.79 | 1.48 | 9.81 * | 0.84 | 9.76 * | 0.84 |
| Insulin (μUI/mL) | 37.68 | 8.26 | 89.17 *** | 12.95 | 48.60 ***† | 12.99 |
| Leptin (ng/mL) | 2.02 | 0.52 | 3.87 *** | 0.53 | 2.25 | 0.53 |
| AIP | 0.44 | 0.09 | 0.72 *** | 0.10 | 0.20 ***† | 0.05 |
| HOMA-IR | 0.97 | 0.34 | 2.80 *** | 0.57 | 1.09 | 0.22 |
| Liver lipids | ||||||
| TAG (mg/g) | 65.99 | 11.74 | 153.58 *** | 15.65 | 71.05 † | 13.34 |
| TC (mg/g) | 10.74 | 1.20 | 46.74 *** | 2.99 | 11.70 † | 3.20 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayer, C.; Côme, M.; Ulmann, L.; Chini Zittelli, G.; Faraloni, C.; Nazih, H.; Ouguerram, K.; Chénais, B.; Mimouni, V. Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats. Nutrients 2019, 11, 1069. https://doi.org/10.3390/nu11051069
Mayer C, Côme M, Ulmann L, Chini Zittelli G, Faraloni C, Nazih H, Ouguerram K, Chénais B, Mimouni V. Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats. Nutrients. 2019; 11(5):1069. https://doi.org/10.3390/nu11051069
Chicago/Turabian StyleMayer, Claire, Martine Côme, Lionel Ulmann, Graziella Chini Zittelli, Cecilia Faraloni, Hassan Nazih, Khadija Ouguerram, Benoît Chénais, and Virginie Mimouni. 2019. "Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats" Nutrients 11, no. 5: 1069. https://doi.org/10.3390/nu11051069
APA StyleMayer, C., Côme, M., Ulmann, L., Chini Zittelli, G., Faraloni, C., Nazih, H., Ouguerram, K., Chénais, B., & Mimouni, V. (2019). Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats. Nutrients, 11(5), 1069. https://doi.org/10.3390/nu11051069