Hypoglycemic Effect of Prolamin from Cooked Foxtail Millet (Setaria italic) on Streptozotocin-Induced Diabetic Mice

 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of PCFM
2.2. Animal Experimental Design
2.3. Biochemical Analysis
2.4. Gut Microbiota Analysis
2.5. Unbiased Metabolomics Analysis
2.6. Data Analysis
3. Results
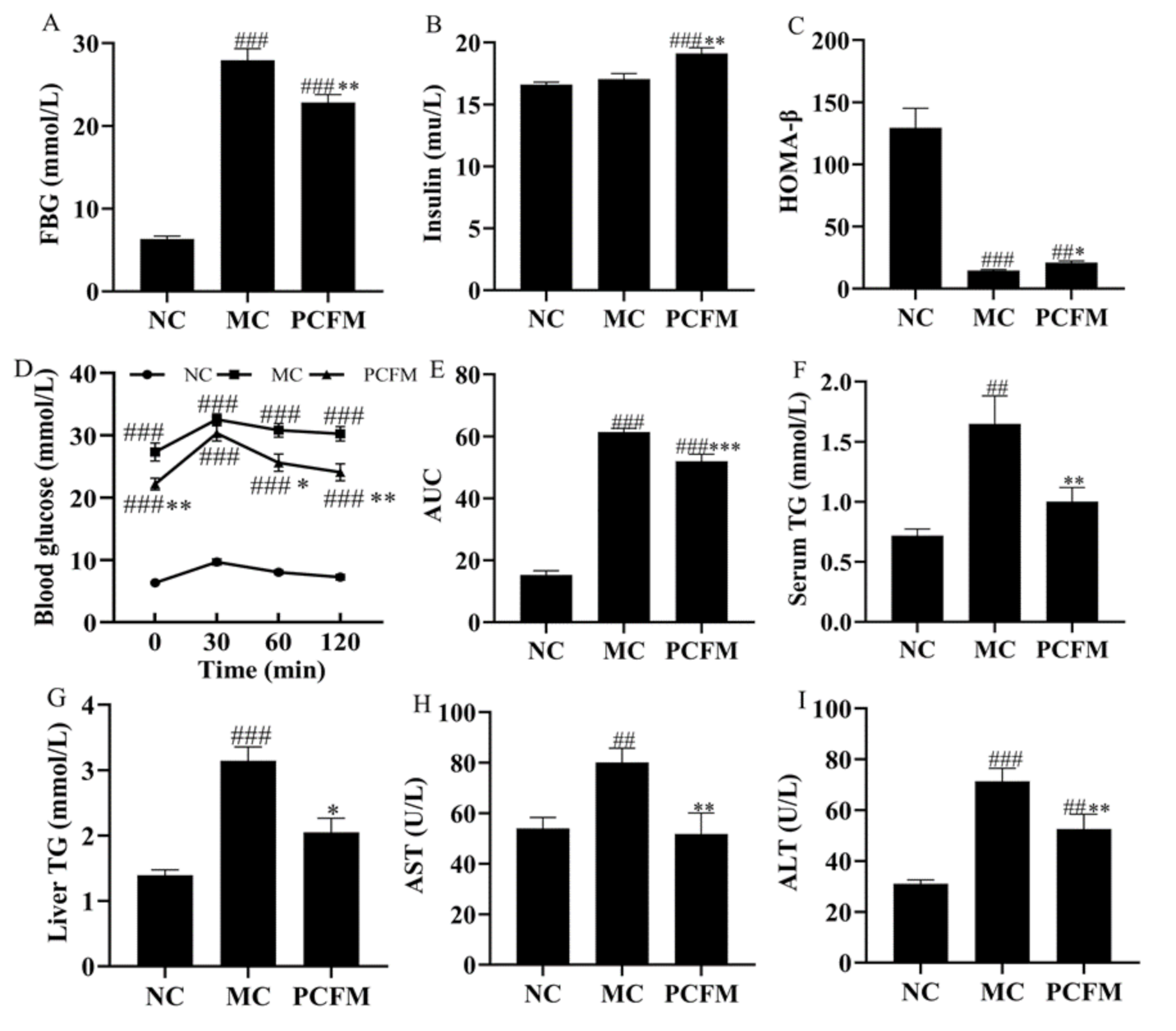
3.1. Effect of PCFM on Diabetes-Related Biomarkers in Diabetic Mice
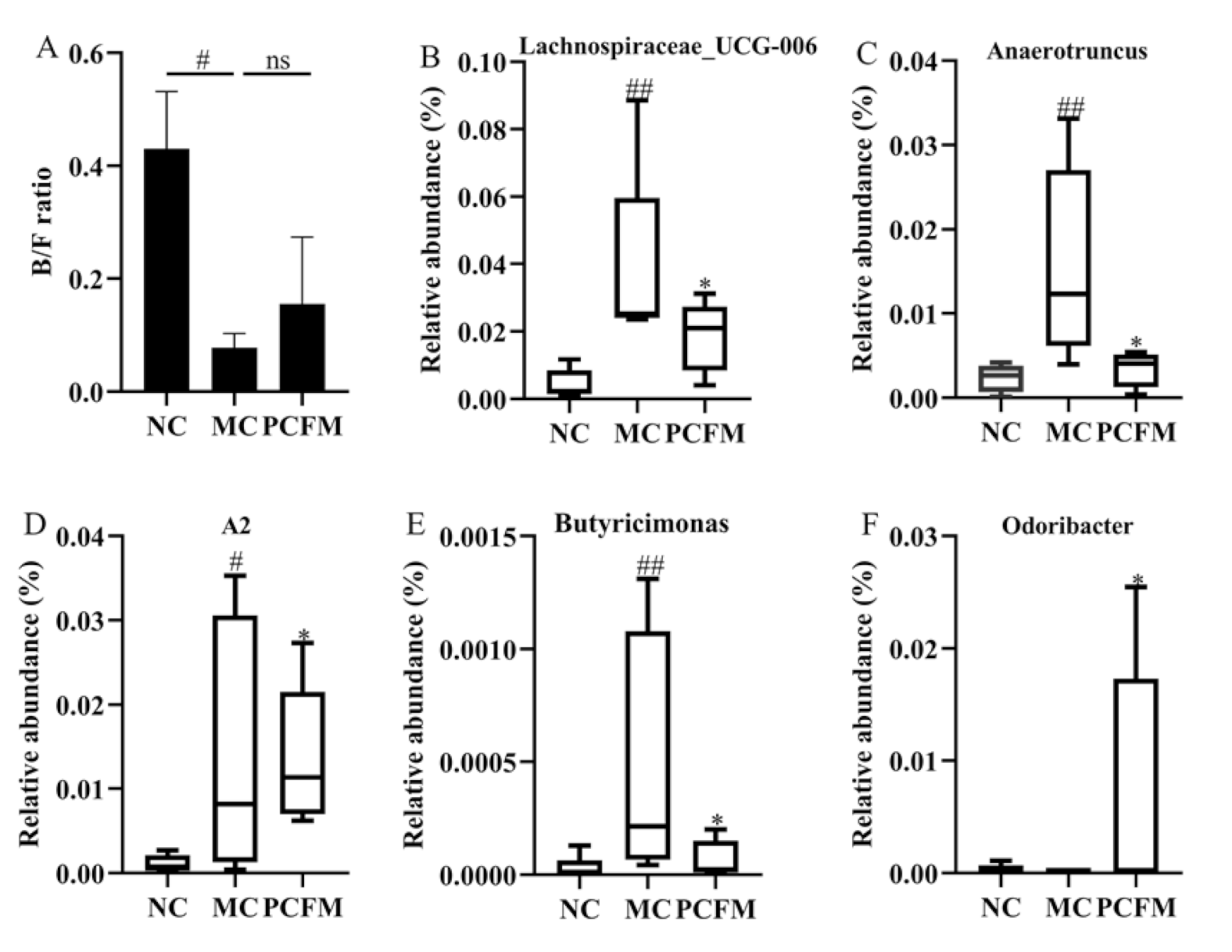
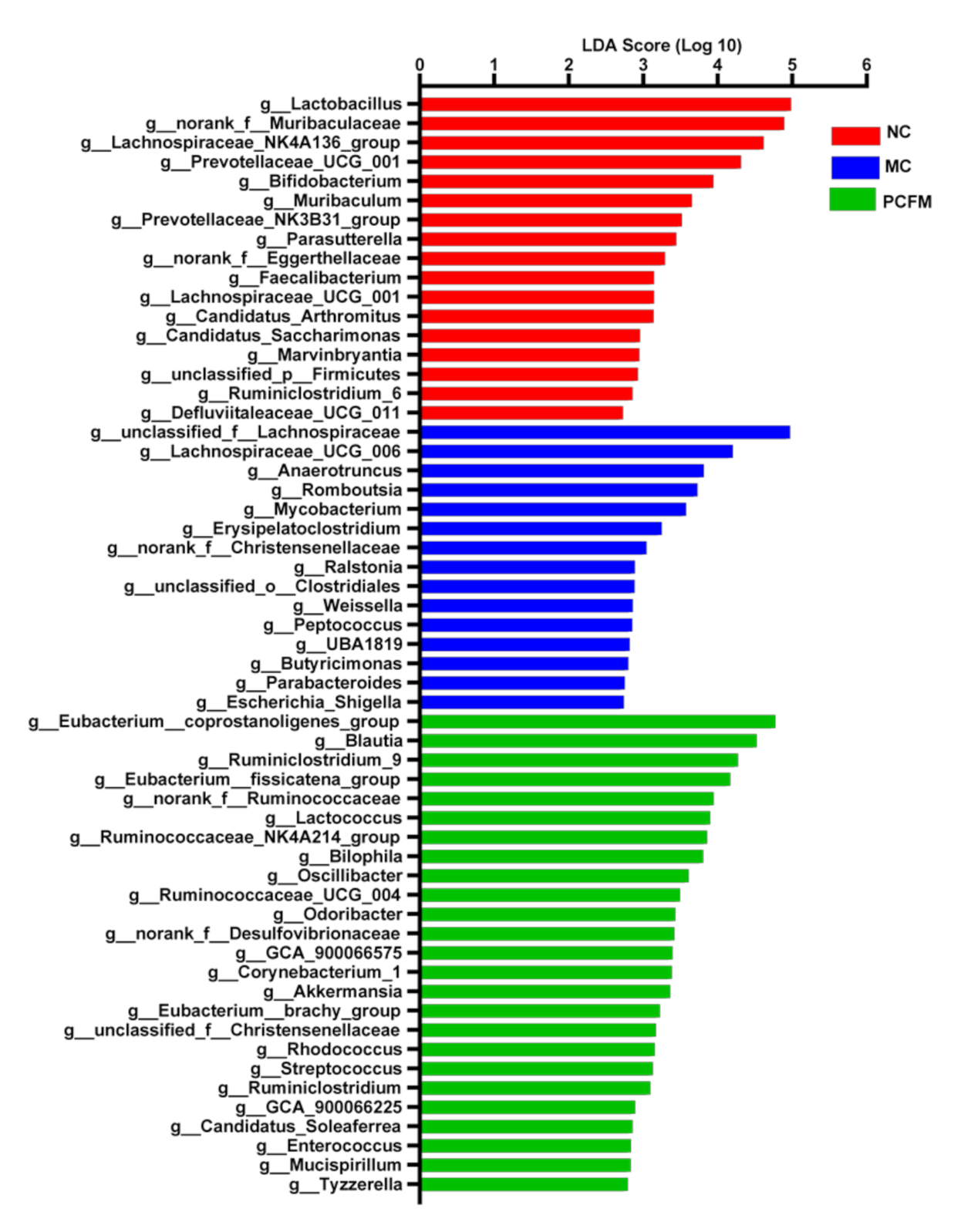
3.2. Effect of Oral PCFM on Gut Microbiota Composition of Mice
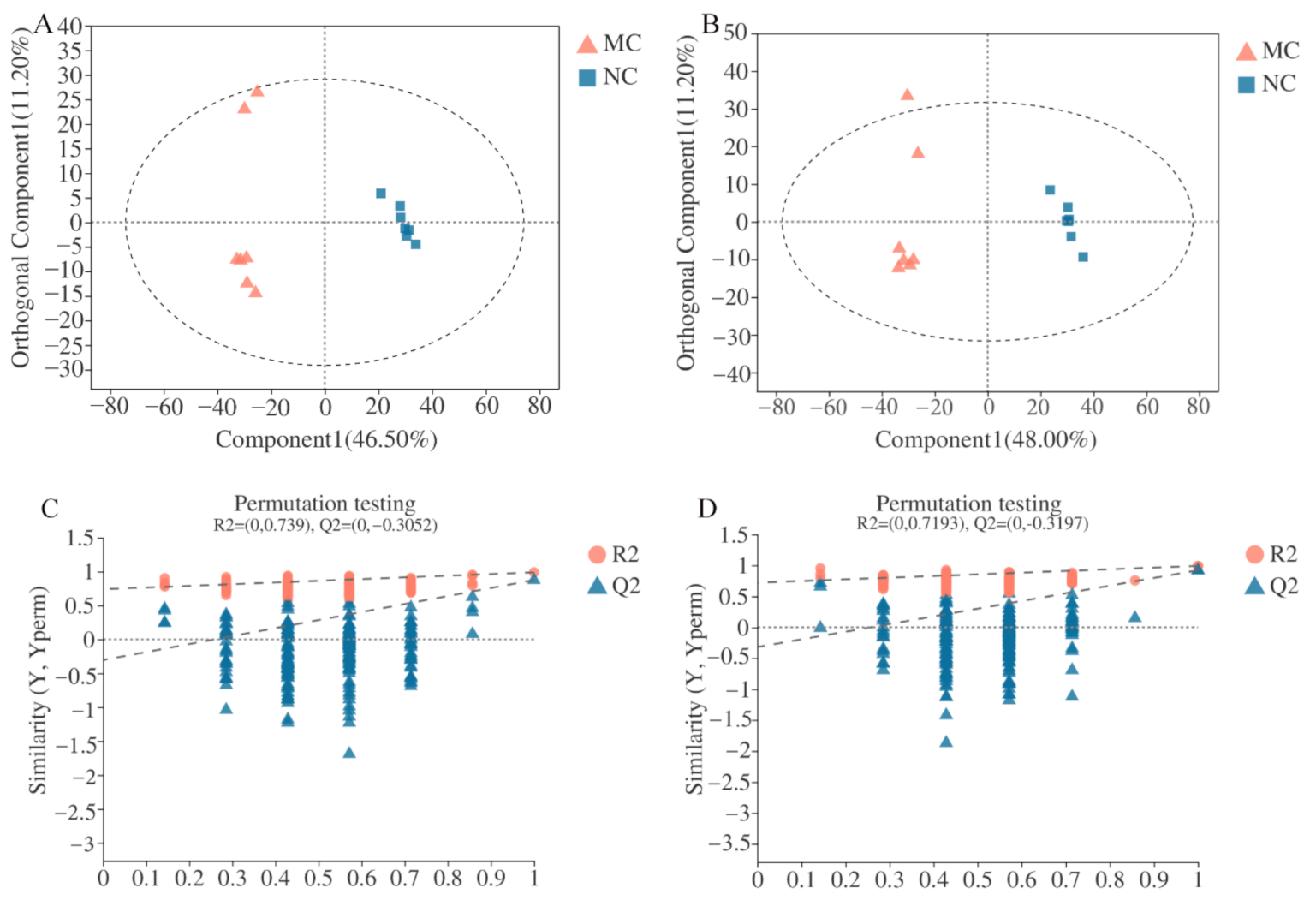
3.3. Serum Metabolites Profiling by UPLC-Q-TOF/MS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Upadhyay, J.; Polyzos, S.A.; Perakakis, N.; Thakkar, B.; Paschou, S.A.; Katsiki, N.; Underwood, P.; Park, K.H.; Seufert, J.; Kang, E.S.; et al. Pharmacotherapy of type 2 diabetes: An update. Metabolism 2018, 78, 13–42. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Snehalatha, C.; Kapur, A.; Vijay, V.; Mohan, V.; Das, A.K.; Rao, P.V.; Yajnik, C.S.; Kumar, P.K.M.; Nair, J.D. High Prevalence of Diabetes and Impaired Glucose Tolerance in India: National Urban Diabetes Survey. Diabetologia 2001, 44, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Tutino, G.E.; Tam, W.H.; Yang, X.; Chan, J.C.N.; Lao, T.T.H.; Ma, R.C.W. Diabetes and pregnancy: Perspectives from Asia. Diabetic Med. 2014, 31, 302–318. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Yang, C.; Wai, S.T.C.; Zhang, Y.; Portillo, M.P.; Paoli, P.; Wu, Y.; Cheang, S.W.; Liu, B.; Carpéné, C.; et al. Regulation of glucose metabolism by bioactive phytochemicals for the management of type 2 diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2019, 59, 830–847. [Google Scholar] [CrossRef]
- Sami, W.; Ansari, T.; Butt, N.S.; Rashid, M.; Hamid, A. Effect of Diet Counseling On Type 2 Diabetes Mellitus. Int. J. Sci. Technol. Res. 2015, 4, 112–118. [Google Scholar]
- Palacios, O.M.; Kramer, M.; Maki, K.C. Diet and prevention of type 2 diabetes mellitus: Beyond weight loss and exercise. Expert Rev. Endocrinol. Metab. 2019, 14, 1–12. [Google Scholar] [CrossRef]
- Kam, J.; Puranik, S.; Yadav, R.; Manwaring, H.R.; Pierre, S.; Srivastava, R.K.; Yadav, R.S. Dietary interventions for type 2 diabetes: How millet comes to help. Front. Plant Sci. 2016, 7, 1454. [Google Scholar] [CrossRef]
- Ren, X.; Yin, R.; Hou, D.; Xue, Y.; Zhang, M.; Diao, X.; Zhang, Y.; Wu, J.; Hu, J.; Hu, X.; et al. The glucose-lowering effect of foxtail millet in subjects with impaired glucose tolerance: A self-controlled clinical trial. Nutrients 2018, 10, 1509. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yu, W.; Guan, X.; Huang, K.; Liu, J.; Liu, D.; Duan, R. Effects of millet whole grain supplementation on the lipid profile and gut bacteria in rats fed with high-fat diet. J. Funct. Foods 2019, 59, 49–59. [Google Scholar] [CrossRef]
- Park, K.O.; Ito, Y.; Nagasawa, T.; Choi, M.R.; Nishizawa, N. Effects of dietary Korean proso-millet protein on plasma adiponectin, HDL cholesterol, insulin levels, and gene expression in obese type 2 diabetic mice. Biosci. Biotechnol. Biochem. 2008, 72, 2918–2925. [Google Scholar] [CrossRef]
- Shahidi, F.; Chandrasekara, A. Millet grain phenolics and their role in disease risk reduction and health promotion: A review. J. Funct. Foods 2013, 5, 570–581. [Google Scholar] [CrossRef]
- Devi, P.B.; Vijayabharathi, R.; Sathyabama, S.; Malleshi, N.G.; Priyadarisini, V.B. Health benefits of finger millet (Eleusine coracana L.) polyphenols and dietary fiber: A review. J. Food Sci. Technol. 2014, 51, 1021–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, N.; Togawa, T.; Park, K.O.; Sato, D.; Miyakoshi, Y.; Inagaki, K.; Ohmori, N.; Ito, Y.; Nagasawa, T. Dietary Japanese millet protein ameliorates plasma levels of adiponectin, glucose, and lipids in type 2 diabetic mice. Biosci. Biotechnol. Biochem. 2009, 3, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubczyk, A.; Szymanowska, U.; Karaś, M.; Złotek, U.; Kowalczyk, D. Potential anti-inflammatory and lipase inhibitory peptides generated by in vitro gastrointestinal hydrolysis of heat treated millet grains. CyTA J. Food 2019, 17, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Feng, R.; Mao, J. Separation and identification of antioxidant peptides from foxtail millet (Setaria italica) prolamins enzymatic hydrolysate. Cereal Chem. 2019, 96, 981–993. [Google Scholar] [CrossRef]
- Duodu, K.G.; Nunes, A.; Delgadillo, I.; Parker, M.L.; Mills, E.N.C.; Belton, P.S.; Taylor, J.R.N. Effect of grain structure and cooking on sorghum and maize in vitro protein digestibility. J. Cereal Sci. 2002, 35, 161–174. [Google Scholar] [CrossRef]
- Duodu, K.G.; Taylor, J.R.N.; Belton, P.S.; Hamaker, B.R. Factors affecting sorghum protein digestibility. J. Cereal Sci. 2003, 35, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Gulati, P.; Li, A.; Holding, D.; Santra, D.; Zhang, Y.; Rose, D.J. Heating Reduces Proso Millet Protein Digestibility via Formation of Hydrophobic Aggregates. J. Agric. Food Chem. 2017, 65, 1952–1959. [Google Scholar] [CrossRef]
- Sharma, N.; Niranjan, K. Foxtail millet: Properties, processing, health benefits, and uses. Food Rev. Int. 2018, 34, 329–363. [Google Scholar] [CrossRef]
- Sitrin, M.D. Digestion and Absorption of Carbohydrates and Proteins. In The Gastrointestinal System, 6th ed.; Leung, P.S., Ed.; Springer: Dordrecht, Germany, 2014; pp. 137–157. [Google Scholar]
- Portune, K.J.; Beaumont, M.; Davila, A.M.; Tomé, D.; Blachier, F.; Sanz, Y. Gut microbiota role in dietary protein metabolism and health-related outcomes: The two sides of the coin. Trends Food Sci. Technol. 2016, 57, 213–232. [Google Scholar] [CrossRef] [Green Version]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Tapia, M.; Tovar, A.R.; Torres, N. Diet as Regulator of Gut Microbiota and its Role in Health and Disease. Arch. Med. Res. 2019, 50, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, A.; Li, X.; Miyamoto, J.; Igarashi, M.; Watanabe, H.; Sutou, A.; Watanabe, K.; Motoyama, T.; Tachibana, N.; Kohno, M.; et al. Dietary mung bean protein reduces high-fat diet-induced weight gain by modulating host bile acid metabolism in a gut microbiota-dependent manner. Biochem. Biophys. Res. Commun. 2018, 501, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, Y.; Hosojima, M.; Kabasawa, H.; Kuwahara, S. Rice Endosperm Protein Administration to Juvenile Mice Regulates Gut Microbiota and Suppresses the Development of High-Fat Diet-Induced Obesity and Related Disorders in Adulthood. Nutrients 2019, 11, 2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sas, K.M.; Karnovsky, A.; Michailidis, G.; Pennathur, S. Metabolomics and diabetes: Analytical and computational approaches. Diabetes 2015, 64, 718–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esko, T.; Hirschhorn, J.N.; Feldman, H.A.; Hsu, Y.H.H.; Deik, A.A.; Clish, C.B.; Ebbeling, C.B.; Ludwig, D.S. Metabolomic profiles as reliable biomarkers of dietary composition. Am. J. Clin. Nutr. 2017, 105, 547–554. [Google Scholar] [CrossRef]
- Shi, L. Untargeted Metabolomics and Novel Data Analysis Strategies to Identify Biomarkers of Diet and Type 2 Diabetes. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Swedish, November 2017. [Google Scholar]
- Li, H.; Li, Y.; Xiang, L.; Zhang, J.J.; Zhu, B.; Xiang, L.; Dong, J.; Liu, M.; Xiang, G. GDF11 attenuates development of type 2 diabetes via improvement of islet β-cells function and survival. Diabetes 2017, 66, 1914–1927. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Yuan, Q.; Yu, R.; Zhang, J.; Ma, H.; Chen, S. Ameliorative Effects of Probiotic Lactobacillus paracasei NL41 on Insulin Sensitivity, Oxidative Stress, and Beta-Cell Function in a Type 2 Diabetes Mellitus Rat Model. Mol. Nutr. Food Res. 2019, 63, 1900457. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhan, B.; Chang, R.; Du, M.; Mao, X. Antidiabetic effect of casein glycomacropeptide hydrolysates on high-fat diet and STZ-induced diabetic mice via regulating insulin signaling in skeletal muscle and modulating gut microbiota. Nutrients 2020, 12, 220. [Google Scholar] [CrossRef] [Green Version]
- Hui, S.; Liu, Y.; Chen, M.; Wang, X.; Lang, H.; Zhou, M.; Yi, L.; Mi, M. Capsaicin Improves Glucose Tolerance and Insulin Sensitivity Through Modulation of the Gut Microbiota-Bile Acid-FXR Axis in Type 2 Diabetic db/db Mice. Mol. Nutr. Food Res. 2019, 63, 1900608. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Huang, M.; You, X.; Zhao, J.; Chen, L.; Wang, L.; Luo, Y.; Chen, Y. Gut microbiota mediates the anti-obesity effect of calorie restriction in mice. Sci. Rep. 2018, 8, 13037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, D.; Zhao, Q.; Yousaf, L.; Khan, J.; Xue, Y.; Qun, S. Consumption of mung bean (Vigna radiata L.) attenuates obesity, ameliorates lipid metabolic disorders and modifies the gut microbiota composition in mice fed a high-fat diet. J. Funct. Foods 2019, 64, 103687. [Google Scholar] [CrossRef]
- Mu, J.; Woods, J.; Zhou, Y.P.; Roy, R.S.; Li, Z.; Zycband, E.; Feng, Y.; Zhu, L.; Li, C.; Howard, A.D.; et al. Chronic inhibition of dipeptidyl peptidase-4 with a sitagliptin analog preserves pancreatic β-cell mass and function in a rodent model of type 2 diabetes. Diabetes 2006, 55, 1695–1704. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, J.; Nazratun Nafizah, A.H.; Zariyantey, A.H.; Budin, S.B. Mechanisms of diabetes-induced liver damage: The role of oxidative stress and inflammation. Sultan Qaboos Univ. Med. J. 2016, 16, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Vozarova, B.; Stefan, N.; Lindsay, R.S.; Saremi, A.; Pratley, R.E.; Bogardus, C.; Antonio Tataranni, P. High alanine aminotransferase is associated with decreased hepatic insulin sensitivity and predicts the development of type 2 diabetes. Diabetes 2002, 51, 1889–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, N.; Vogensen, F.K.; Van Den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE. 2010, 5, 9085. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Long, Y.; Lu, M. Exploration of variations in proteome and metabolome for predictive diagnostics and personalized treatment algorithms: Innovative approach and examples for potential clinical application. J. Proteom. 2018, 188, 30–40. [Google Scholar] [CrossRef]
- Torre-Villalvazo, I.; Tovar, A.R.; Ramos-Barragán, V.E.; Cerbón-Cervantes, M.A.; Torres, N. Soy protein ameliorates metabolic abnormalities in liver and adipose tissue of rats fed a high fat diet. J. Nutr. 2008, 138, 462–468. [Google Scholar] [CrossRef]
- Mourad, H.H.; El-kassaby, M.I.; El-hussieny, E.A.; Esmail, R.S.; Mannaa, F.A.; Khaled, G. Role of soy protein concentrate on oxidative stress and DNA fragmentation in streptozotocin-induced diabetic rats. J. Innov. Pharm. Biol. Sci. 2017, 4, 16–25. [Google Scholar]
- Boonloh, K.; Lee, E.S.; Kim, H.M.; Kwon, M.H.; Kim, Y.M.; Pannangpetch, P.; Kongyingyoes, B.; Kukongviriyapan, U.; Thawornchinsombut, S.; Lee, E.Y. Rice bran protein hydrolysates attenuate diabetic nephropathy in diabetic animal model. Eur. J. Nutr. 2018, 57, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Hakkak, R.; Gauss, C.H.; Bell, A.; Korourian, S. Short-term soy protein isolate feeding prevents liver steatosis and reduces serum ALT and AST levels in obese female zucker rats. Biomedicines 2018, 6, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asokan, S.M.; Wang, T.; Su, W.T.; Lin, W.T. Antidiabetic effects of a short peptide of potato protein hydrolysate in STZ-induced diabetic mice. Nutrients 2019, 11, 779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Yuan, X.; Cheng, G.; Jiao, S.; Feng, C.; Zhao, X.; Yin, H.; Du, Y.; Liu, H. Chitosan oligosaccharides improve the disturbance in glucose metabolism and reverse the dysbiosis of gut microbiota in diabetic mice. Carbohydr. Polym. 2018, 190, 77–86. [Google Scholar] [CrossRef]
- Song, Y.; Wu, M.; Tao, G.; Lu, M.; Lin, J.; Huang, J. Feruloylated oligosaccharides and ferulic acid alter gut microbiome to alleviate diabetic syndrome. Food Res. Int. 2020, 137, 109410. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, A.; Alkhalidy, H.; Luo, J.; Moomaw, E.; Neilson, A.P.; Liu, D. Flavone Hispidulin Stimulates Glucagon-Like Peptide-1 Secretion and Ameliorates Hyperglycemia in Streptozotocin-Induced Diabetic Mice. Mol. Nutr. Food Res. 2020, 64, 1900978. [Google Scholar] [CrossRef]
- Zheng, J.; Zhu, L.; Hu, B.; Zou, X.; Hu, H.; Zhang, Z.; Jiang, N.; Ma, J.; Yang, H.; Liu, H. 1-Deoxynojirimycin improves high fat diet-induced nonalcoholic steatohepatitis by restoring gut dysbiosis. J. Nutr. Biochem. 2019, 71, 16–26. [Google Scholar] [CrossRef]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.M.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; De Vos, W.M.; et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Xiao, X.; Zheng, J.; Li, M.; Yu, M.; Ping, F.; Wang, T.; Wang, X. Featured article: Structure moderation of gut microbiota in liraglutide-treated diabetic male rats. Exp. Biol. Med. 2018, 243, 34–44. [Google Scholar] [CrossRef]
- Xu, J.; Lian, F.; Zhao, L.; Zhao, Y.; Chen, X.; Zhang, X.; Guo, Y.; Zhang, C.; Zhou, Q.; Xue, Z.; et al. Structural modulation of gut microbiota during alleviation of type 2 diabetes with a Chinese herbal formula. ISME J. 2015, 9, 552–562. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Q.; Li, H.; Wen, C.; He, Z. Alterations of the gut microbiome associated with the treatment of hyperuricaemia in male rats. Front. Microbiol. 2018, 9, 2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, L.M.; Creasy, H.H.; Fettweis, J.M.; Lloyd-Price, J.; Mahurkar, A.; Zhou, W.; Buck, G.A.; Snyder, M.P.; Strauss, J.F.; Weinstock, G.M.; et al. The Integrative Human Microbiome Project. Nature 2019, 569, 641–648. [Google Scholar]
- Zhou, W.; Sailani, M.R.; Contrepois, K.; Zhou, Y.; Ahadi, S.; Leopold, S.R.; Zhang, M.J.; Rao, V.; Avina, M.; Mishra, T.; et al. Longitudinal multi-omics of host–microbe dynamics in prediabetes. Nature 2019, 569, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, Y.; Xu, J.; Xue, Z.; Zhang, M.; Pang, X.; Zhang, X.; Zhao, L. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci. Rep. 2015, 5, 14405. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.; Xu, J.; Lian, F.; Yu, X.; Zhao, Y.; Xu, L.; Zhang, M.; Zhao, X.; Shen, J.; Wu, S.; et al. Structural alteration of gut microbiota during the amelioration of human type 2 diabetes with hyperlipidemia by metformin and a traditional chinese herbal formula: A multicenter, randomized, open label clinical trial. MBio 2018, 9, 2392–2417. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Qin, Q.; Liu, M.; Zhang, X.; He, F.; Wang, G. Akkermansia muciniphila can reduce the damage of gluco/lipotoxicity, oxidative stress and inflammation, and normalize intestine microbiota in streptozotocin-induced diabetic rats. Pathog. Dis. 2018, 76, fty028. [Google Scholar] [CrossRef] [Green Version]
- De La Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin is associated with higher relative abundance of mucin-degrading akkermansia muciniphila and several short-chain fatty acid-producing microbiota in the gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Mukherjee, S.; Ayele, B.T. Molecular aspects of sucrose transport and its metabolism to starch during seed development in wheat: A comprehensive review. Biotechnol. Adv. 2018, 36, 954–967. [Google Scholar] [CrossRef]
- Nordlie, R.C.; Foster, J.D.; Lange, A.J. Regulation of glucose production by the liver. Annu. Rev. Nutr. 1999, 19, 379–406. [Google Scholar] [CrossRef]
- Berry, G.T. Disorders of Galactose Metabolism. In Rosenberg’s Molecular and Genetic Basis of Neurological and Psychiatric Disease (Fifth Edition), 3rd ed.; Rosenberg, R.N., Pascual, J.M., Eds.; Academic Press: Dallas, TX, USA, 2015; pp. 615–626. [Google Scholar]
- Wang, J. Glycolysis. In Biochemistry, 22nd ed.; Wang, J., Zhu, S., Xu, C., Eds.; Higher Education Press: Beijing, China, 2002; pp. 63–91. [Google Scholar]
- Holden, H.M.; Rayment, I.; Thoden, J.B. Structure and function of enzymes of the Leloir pathway for galactose metabolism. J. Biol. Chem. 2003, 278, 43885–43888. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Wang, Y.; Ong, C.N.; Subramaniam, T.; Choi, H.W.; Yuan, J.M.; Koh, W.P.; Pan, A. Metabolic signatures and risk of type 2 diabetes in a Chinese population: An untargeted metabolomics study using both LC-MS and GC-MS. Diabetologia 2016, 59, 2349–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulmann, N.; Grohmann, M.; Voigt, J.P.; Bert, B.; Vowinckel, J.; Bader, M.; Skelin, M.; Jevšek, M.; Fink, H.; Rupnik, M.; et al. Intracellular serotonin modulates insulin secretion from pancreatic β-cells by protein serotonylation. PLoS Biol. 2009, 7, 1000229. [Google Scholar] [CrossRef] [Green Version]
- Lam, D.D.; Heisler, L.K. Serotonin and energy balance: Molecular mechanisms and implications for type 2 diabetes. Expert Rev. Mol. Med. 2007, 9, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Tavintharan, S.; Sum, C.F.; Woon, K.; Lim, S.C.; Ong, C.N. Metabolic signature shift in type 2 diabetes mellitus revealed by mass spectrometry-based metabolomics. J. Clin. Endocrinol. Metab. 2013, 98, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Ugarte, M.; Brown, M.; Hollywood, K.A.; Cooper, G.J.; Bishop, P.N.; Dunn, W.B. Metabolomic analysis of rat serum in streptozotocin-induced diabetes and after treatment with oral triethylenetetramine (TETA). Genome Med. 2012, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drogan, D.; Dunn, W.B.; Lin, W.; Buijsse, B.; Schulze, M.B.; Langenberg, C.; Brown, M.; Floegel, A.; Dietrich, S.; Rolandsson, O.; et al. Untargeted metabolic profiling identifies altered serum metabolites of type 2 diabetes mellitus in a prospective, nested case control study. Clin. Chem. 2015, 61, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.I.; Jang, H.D.; Shetty, K. Evaluation of Rhodiola crenulata and Rhodiola rosea for management of Type II diabetes and hypertension. Asia Pac. J. Clin. Nutr. 2006, 15, 425–432. [Google Scholar]
- Chandramohan, R.; Pari, L. Anti-inflammatory effects of tyrosol in streptozotocin-induced diabetic Wistar rats. J. Funct. Foods 2016, 27, 17–28. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | MC/NC | PCFM/MC | PCFM/NC | KEGG Pathway |
|---|---|---|---|---|
| Lysophosphatidylcholine (LysoPC) (16:1(9Z)/0:0) | ↓ *** | ↑ # | ↓ &&& | Glycerophospholipid metabolism |
| Serotonin | ↓ *** | ↑ ## | ↓ && | Tryptophan metabolism |
| Uridine diphosphogalactose (UDP)-L-rhamnose | ↓ * | ↑ # | ↓ | Amino sugar and nucleotide sugar metabolism |
| LysoPC (22:1(13Z)) | ↓ *** | ↑ # | ↓ &&& | Glycerophospholipid metabolism |
| LysoPC (14:1(9Z)) | ↓ ** | ↑ ### | ↑ | Glycerophospholipid metabolism |
| M-Coumaric acid | ↓ ** | ↑ ## | ↓ | Phenylalanine metabolism |
| Tyrosol | ↓ ** | ↑ ## | ↓ | Tyrosine metabolism |
| Lactosylceramide (d18:1/12:0) | ↑ ** | ↓ # | ↑ | Sphingolipid metabolism |
| Prostaglandin J2 | ↑ *** | ↓ ## | ↑ | Arachidonic acid metabolism |
| 9S,11R,15S-trihydroxy-2,3-dinor-13E-prostaenoic acid-cyclo [8 S,12R] | ↑ *** | ↓ # | ↑ | Arachidonic acid metabolism |
| Estriol | ↑ *** | ↓ ### | ↑ | Steroid hormone biosynthesis |
| 9,10,13-Trihydroxyoctadecenoic acid (TriHOME) | ↑ *** | ↓ ### | ↑ | Linoleic acid metabolism |
| 9(S)-Hydroperoxyoctadecatrienoic acid (HpOTrE) | ↑ *** | ↓ ### | ↑ | alpha-Linolenic acid metabolism |
| 19-Hydroxyandrost-4-ene-3,17-dione | ↑ ** | ↓ ### | ↑ | Steroid hormone biosynthesis |
| D-Glucose | ↑ *** | ↓ # | ↑ &&& | Starch and sucrose metabolism; Glycolysis / Gluconeogenesis; Galactose metabolism |
| LysoPC (20:1(11Z)) | ↓ *** | ↑ | ↓ &&& | Glycerophospholipid metabolism |
| LysoPC (20:0/0:0) | ↓ *** | ↑ | ↓ & | Glycerophospholipid metabolism |
| Retinol | ↓ *** | ↑ | ↓ & | Retinol metabolism |
| Glycocholic Acid | ↓ *** | ↑ | ↓ && | Primary bile acid biosynthesis |
| 4-O-alpha-D-Galactopyranuronosyl-D-galacturonic acid | ↓ * | ↑ | ↓ & | Pentose and glucuronate interconversions |
| Sucrose | ↑ ** | ↓ | ↑ && | Galactose metabolism; Starch and sucrose metabolism |
| N6-Acetyl-L-lysine | ↑ ** | ↓ | ↑ | Lysine degradation |
| 5’-Deoxy-5-fluorouridine | ↑ ** | ↓ | ↑ | Xenobiotics biodegradation and metabolism |
| Thiamine | ↑ * | ↑ | ↑ | Thiamine metabolism |
| Acetylcholine | ↓ *** | ↓ | ↓ &&& | Bile secretion; Glycerophospholipid metabolism |
| Phosphoserine | ↑ *** | ↑ | ↑ &&& | Glycine, serine and threonine metabolism; Cysteine and methionine metabolism |
| Phosphatidyl choline (PC) (22:5 (4Z,7Z,10Z,13Z,16Z)/P-18:0) | ↑ * | ↑ # | ↑ && | Linoleic acid metabolism; Arachidonic acid metabolism; alpha-Linolenic acid metabolism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Yin, R.; Liu, Z.; Niu, Y.; Guo, E.; Cheng, R.; Diao, X.; Xue, Y.; Shen, Q. Hypoglycemic Effect of Prolamin from Cooked Foxtail Millet (Setaria italic) on Streptozotocin-Induced Diabetic Mice. Nutrients 2020, 12, 3452. https://doi.org/10.3390/nu12113452
Fu Y, Yin R, Liu Z, Niu Y, Guo E, Cheng R, Diao X, Xue Y, Shen Q. Hypoglycemic Effect of Prolamin from Cooked Foxtail Millet (Setaria italic) on Streptozotocin-Induced Diabetic Mice. Nutrients. 2020; 12(11):3452. https://doi.org/10.3390/nu12113452
Chicago/Turabian StyleFu, Yongxia, Ruiyang Yin, Zhenyu Liu, Yan Niu, Erhu Guo, Ruhong Cheng, Xianmin Diao, Yong Xue, and Qun Shen. 2020. "Hypoglycemic Effect of Prolamin from Cooked Foxtail Millet (Setaria italic) on Streptozotocin-Induced Diabetic Mice" Nutrients 12, no. 11: 3452. https://doi.org/10.3390/nu12113452
APA StyleFu, Y., Yin, R., Liu, Z., Niu, Y., Guo, E., Cheng, R., Diao, X., Xue, Y., & Shen, Q. (2020). Hypoglycemic Effect of Prolamin from Cooked Foxtail Millet (Setaria italic) on Streptozotocin-Induced Diabetic Mice. Nutrients, 12(11), 3452. https://doi.org/10.3390/nu12113452