
The Marine Microalga, Tisochrysis lutea, Protects against Metabolic Disorders Associated with Metabolic Syndrome and Obesity

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Diets
2.2. Blood and Organ Sampling
2.3. Biochemical Analyses
2.4. Hepatic Lipid Measurements
2.5. Statistical Analysis
3. Results
3.1. T. lutea Supplementation Decreases Body Weight and Fat Mass in Wistar Rats Fed a High-Fat Diet
3.1.1. Nutritional Monitoring: Food and Water Intake
3.1.2. Energy Intake and Food Efficiency
3.1.3. Body Weight and Fat Mass
3.2. Effects of T. lutea Supplementation on Physiological and Metabolic Disorders in Wistar Rats Fed a High-Fat Diet
3.2.1. Plasma Biochemical Parameters and HOMA-IR Index
3.2.2. Plasma Lipids, HDL/LDL Ratio and AIP Index
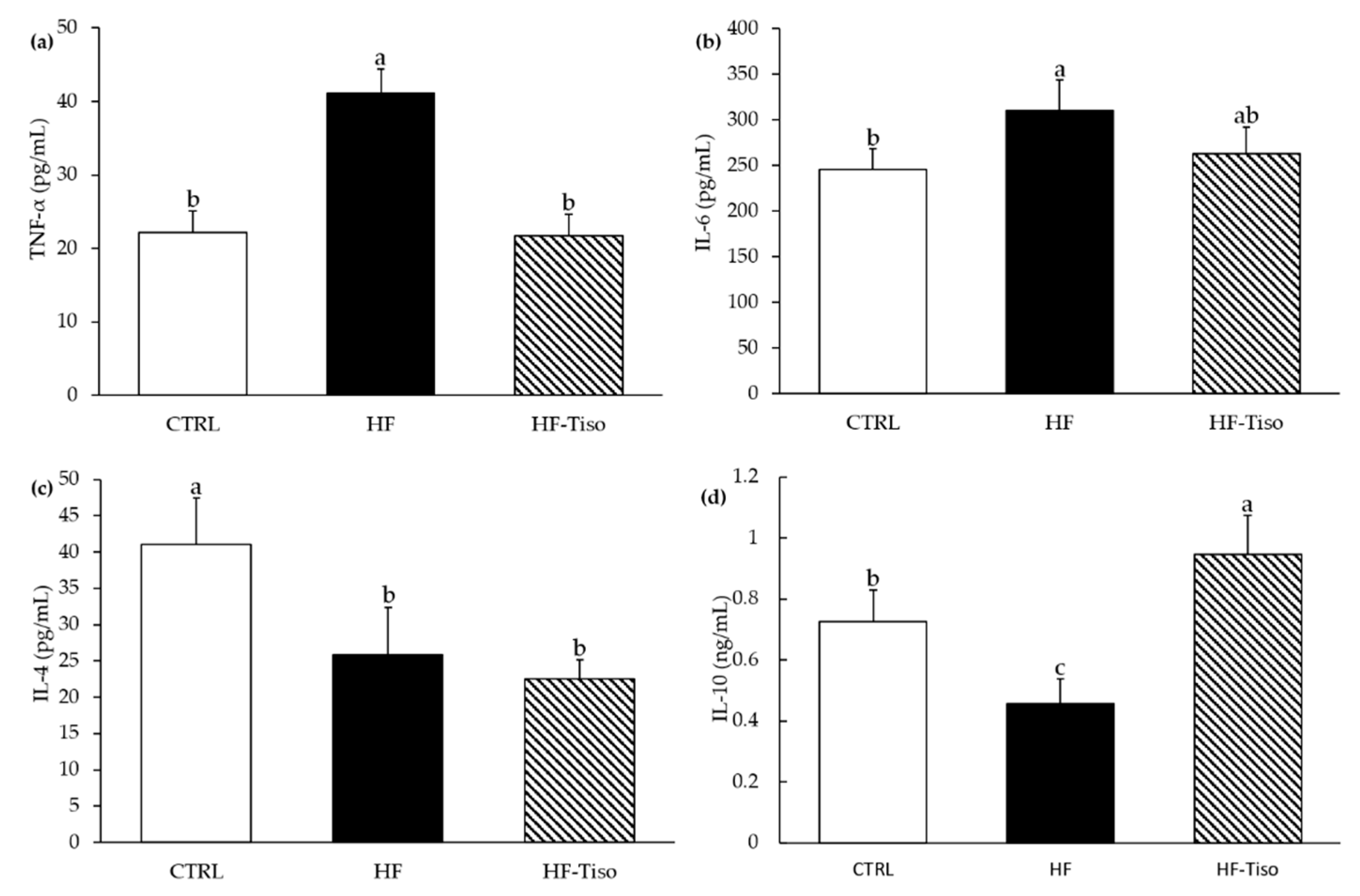
3.3. Effects of T. lutea on Inflammatory Status
3.3.1. Pro-Inflammatory and Anti-Inflammatory Cytokines
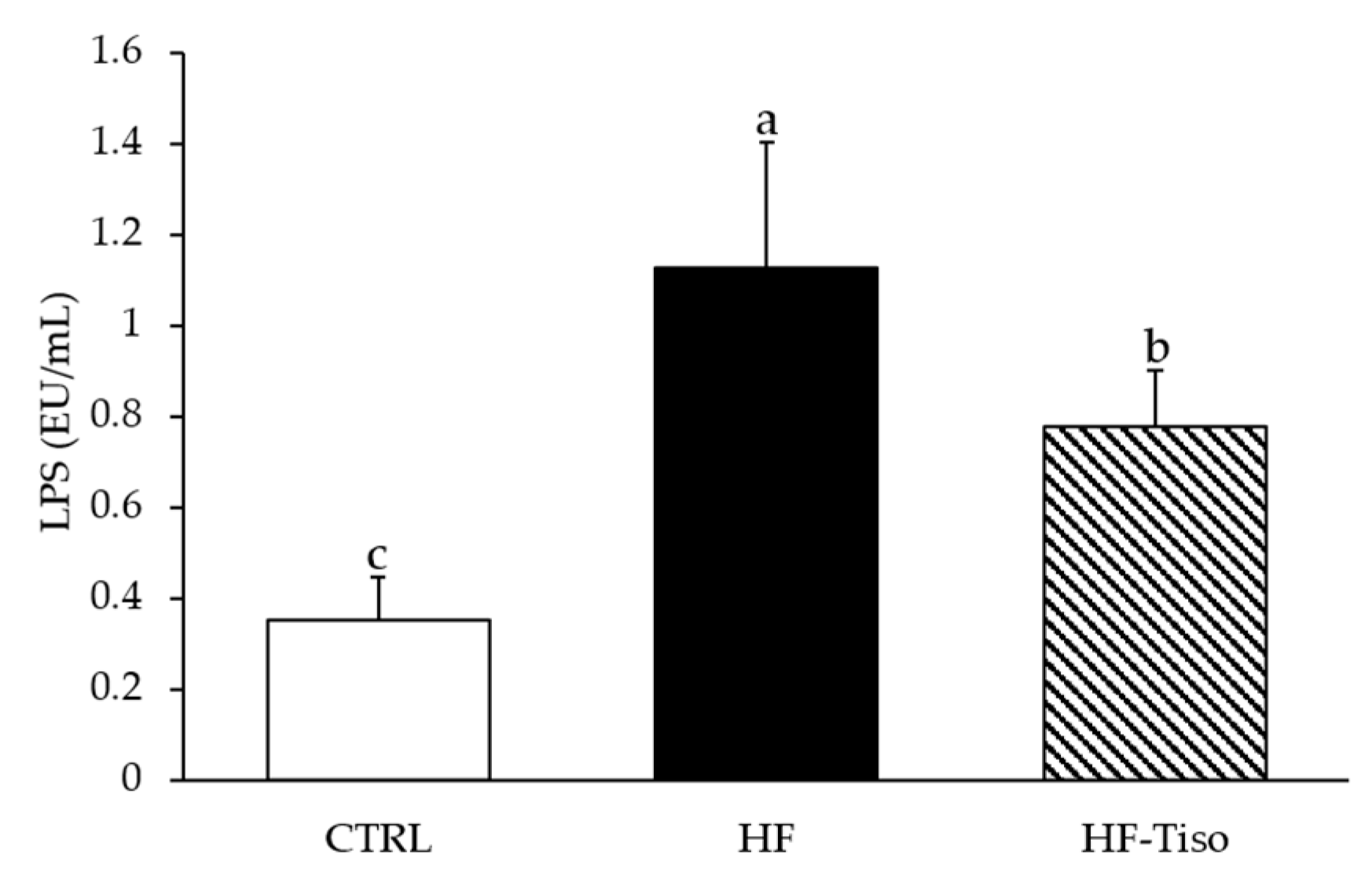
3.3.2. Serum LPS Levels
3.4. Effects of T. lutea on Liver Triglyceride and Total Cholesterol Levels
4. Discussion
4.1. T. lutea Supplementation Reduced Body Weight, Abdominal and Epididymal Adipose Weights in HF-Fed Wistar Rats
4.2. Effects of T. lutea, as a Dietary Supplement, in the Prevention of Dyslipidemia and Atherosclerosis
4.3. Effects of T. lutea Supplementation on Inflammatory Status in HF-Fed Wistar Rats
4.4. Hypoglycemic and Hypoinsulinemic Effects of T. lutea Supplementation in HF-Fed Wistar Rats
4.5. Effects of T. lutea on Non-Alcoholic Fatty Liver Disease
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Global Health Estimates 2016: Deaths by Cause, Age, Sex, by Country and by Region, 2000–2016; World Health Organization: Genève, Switzerland, 2018. [Google Scholar]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [PubMed] [Green Version]
- Després, J.-P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nat. Cell Biol. 2006, 444, 881–887. [Google Scholar] [CrossRef]
- Browning, J.D.; Szczepaniak, L.S.; Dobbins, R.; Nuremberg, P.; Horton, J.D.; Cohen, J.C.; Grundy, S.M.; Hobbs, H.H. Prevalence of hepatic steatosis in an urban population in the United States: Impact of ethnicity. Hepatology 2004, 40, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Luyendyk, J.P.; Guo, G.L. Steatosis DeLIVERs High-Sensitivity C-Reactive Protein. Arter. Thromb. Vasc. Biol. 2011, 31, 1714–1715. [Google Scholar] [CrossRef] [Green Version]
- Marhuenda, J.; Villaño, D.; Cerdá, B.; Zafrilla, P. Cardiovascular disease and nutrition. In Nutrition in Health and Disease; Mogzik, G., Fliger, M., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Manson, J.E.; Cook, N.R.; Lee, I.-M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Albert, C.M.; Gordon, D.; Copeland, T.; et al. Marine n−3 Fatty Acids and Prevention of Cardiovascular Disease and Cancer. N. Engl. J. Med. 2019, 380, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Albracht-Schulte, K.; Kalupahana, N.S.; Ramalingam, L.; Wang, S.; Rahman, S.M.; Robert-McComb, J.; Moustaid-Moussa, N. Omega-3 fatty acids in obesity and metabolic syndrome: A mechanistic update. J. Nutr. Biochem. 2018, 58, 1–16. [Google Scholar] [CrossRef]
- Yashodhara, B.M.; Umakanth, S.; Pappachan, J.M.; Bhat, S.K.; Kamath, R.; Choo, B.H. Omega-3 fatty acids: A comprehensive review of their role in health and disease. Postgrad. Med. J. 2009, 85, 84–90. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Kim, H.-J.; Takahashi, M.; Ezaki, O. Fish oil feeding decreases mature sterol regulatory element-binding protein 1 (SREBP-1) by down-regulation of SREBP-1c mRNA in mouse liver: A possible mechanism for down regulation of lipogenic enzyme mRNAs. J. Biol. Chem. 1999, 274, 25892–25898. [Google Scholar] [CrossRef] [Green Version]
- Chadli, F.K.; Andre, A.; Prieur, X.; Loirand, G.; Meynier, A.; Krempf, M.; Nguyen, P.; Ouguerram, K. n-3 PUFA prevent metabolic disturbances associated with obesity and improve endothelial function in golden Syrian hamsters fed with a high-fat diet. Br. J. Nutr. 2011, 107, 1305–1315. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.A.; Custódio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.C.; Abu-Salah, K.M. Alternative Sources of n-3 Long-Chain Polyunsaturated Fatty Acids in Marine Microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef]
- Lakra, N.; Mahmood, S.; Marwal, A.; Sudheep, N.M.; Anwar, K. Bioengineered plants can be an alternative source of omega-3 fatty acids for human health. In Plant and Human Health; Ozturk, M., Hakeem, K.R., Eds.; Springer International Publishing: Cham, Switzerland, 2019; Volume 2, pp. 361–382. [Google Scholar]
- Hamed, I. The Evolution and Versatility of Microalgal Biotechnology: A Review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1104–1123. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wu, Y.; Yang, C.; Liu, B.; Huang, Y. Hypotensive, hypoglycaemic and hypolipidaemic effects of bioactive compounds from microalgae and marine micro-organisms. Int. J. Food Sci. Technol. 2015, 50, 1705–1717. [Google Scholar] [CrossRef]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-P.; Lin, L.-P. Ultrastructural study and lipid formation of Isochrysis sp. CCMP1324. Bot. Bull. Acad. Sin. 2001, 42, 207–214. [Google Scholar]
- O’Shea, S.K.; Holland, F.; Bilodeau, A. Modeling the Effects of Salinity and pH on the Cadmium Bioabsorptive Properties of the MicroalgaeIsochrysis galbana(T-Iso) in Coastal Waters. J. Coast. Res. 2010, 261, 59–66. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, R.M.S.C.; de Morais, A.M. Health applications of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef]
- Alkhamis, Y.; Qin, J.G. Comparison of pigment and proximate compositions of Tisochrysis lutea in phototrophic and mixotrophic cultures. J. Appl. Phycol. 2016, 28, 35–42. [Google Scholar] [CrossRef]
- Mimouni, V.; Couzinet-Mossion, A.; Ulmann, L.; Wielgosz-Collin, G. Lipids from microalgae. In Microalgae in Health and Disease Prevention; Levine, I., Fleurence, J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 109–131. [Google Scholar]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Fairus, A.; Soelaiman, I.-N. Animal models of metabolic syndrome: A review. Nutr. Metab. 2016, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.L.R.P.; Leite, L.H.R.; Gioda, C.R.; Leme, F.O.P.; Couto, C.A.; Coimbra, C.C.; Leite, V.H.R.; Ferrari, T.C.A. A Novel Wistar Rat Model of Obesity-Related Nonalcoholic Fatty Liver Disease Induced by Sucrose-Rich Diet. J. Diabetes Res. 2015, 2016, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buettner, R.; Schölmerich, J.; Bollheimer, L.C. High-fat Diets: Modeling the Metabolic Disorders of Human Obesity in Rodents. Obesity 2007, 15, 798–808. [Google Scholar] [CrossRef]
- Kakimoto, P.A.; Kowaltowski, A.J. Effects of high fat diets on rodent liver bioenergetics and oxidative imbalance. Redox Biol. 2016, 8, 216–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.; Meireles, M.; Norberto, S.; Leite, J.; Freitas, J.; Pestana, D.; De Faria, A.M.C.; Calhau, C. High-fat diet-induced obesity Rat model: A comparison between Wistar and Sprague-Dawley Rat. Adipocyte 2015, 5, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, High-fat Diet–induced Metabolic Syndrome and Cardiovascular Remodeling in Rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Haimeur, A.; Mimouni, V.; Ulmann, L.; Martineau, A.-S.; Messaouri, H.; Pineau-Vincent, F.; Tremblin, G.; Meskini, N. Fish Oil and Microalga Omega-3 as Dietary Supplements: A Comparative Study on Cardiovascular Risk Factors in High-Fat Fed Rats. Lipids 2016, 51, 1037–1049. [Google Scholar] [CrossRef]
- Mayer, C.; Côme, M.; Ulmann, L.; Zittelli, G.C.; Faraloni, C.; Nazih, H.; Ouguerram, K.; Chénais, B.; Mimouni, V. Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats. Nutrients 2019, 11, 1069. [Google Scholar] [CrossRef] [Green Version]
- Toop, C.R.; Gentili, S. Fructose Beverage Consumption Induces a Metabolic Syndrome Phenotype in the Rat: A Systematic Review and Meta-Analysis. Nutrients 2016, 8, 577. [Google Scholar] [CrossRef] [Green Version]
- Novelli, E.; Diniz, Y.S.; Galhardi, C.M.; Ebaid, G.M.X.; Rodrigues, H.G.; Mani, F.; Fernandes, A.A.H.; Cicogna, A.C.; Filho, J.L.V.B.N. Anthropometrical parameters and markers of obesity in rats. Lab. Anim. 2007, 41, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Marsh, J.B.; Weinstein, D.B. Simple charring method for determination of lipids. J. Lipid Res. 1966, 7, 574–576. [Google Scholar] [CrossRef]
- Simionato, J.I.; Garcia, J.C.; Santos, G.T.; Oliveira, C.C.; Visentainer, J.V.; Souza, N.E. Validation of the determination of fatty acids in milk by gas chromatography. J. Braz. Chem. Soc. 2010, 21, 520–524. [Google Scholar] [CrossRef]
- Vohra, D.F. Determination of photosynthetic pigments in sea-water. In Monographs Onocéanographie Methodology; UNESCO, Ed.; UNESCO: Paris, France, 1966; p. 66. [Google Scholar]
- Van Heukelem, L.; Thomas, C.S. Computer-assisted high-performance liquid chromatography method development with applications to the isolation and analysis of phytoplankton pigments. J. Chromatogr. A 2001, 910, 31–49. [Google Scholar] [CrossRef]
- Boisen, S.; Fernández, J. Prediction of the total tract digestibility of energy in feedstuffs and pig diets by in vitro analyses. Anim. Feed. Sci. Technol. 1997, 68, 277–286. [Google Scholar] [CrossRef]
- Batista, A.P.; Niccolai, A.; Fradinho, P.; Fragoso, S.; Bursic, I.; Rodolfi, L.; Biondi, N.; Tredici, M.R.; De Sousa, I.M.N.; Raymundo, A. Microalgae biomass as an alternative ingredient in cookies: Sensory, physical and chemical properties, antioxidant activity and in vitro digestibility. Algal Res. 2017, 26, 161–171. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and Mechanisms of Antioxidant Activity using the DPPH. Free Radical Method. LWT 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Frohlich, J.; Dobiásová, M. Fractional Esterification Rate of Cholesterol and Ratio of Triglycerides to HDL-Cholesterol Are Powerful Predictors of Positive Findings on Coronary Angiography. Clin. Chem. 2003, 49, 1873–1880. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Parrish, C.; Pathy, D.; Angel, A. Dietary fish oils limit adipose tissue hypertrophy in rats. Metabolism 1990, 39, 217–219. [Google Scholar] [CrossRef]
- Ruzickova, J.; Rossmeisl, M.; Prazak, T.; Flachs, P.; Sponarova, J.; Vecka, M.; Tvrzicka, E.; Bryhn, M.; Kopecký, J. Omega-3 PUFA of marine origin limit diet-induced obesity in mice by reducing cellularity of adipose tissue. Lipids 2004, 39, 1177–1185. [Google Scholar] [CrossRef]
- Murali, G.; DeSouza, C.V.; Clevenger, M.E.; Ramalingam, R.; Saraswathi, V. Differential effects of eicosapentaenoic acid and docosahexaenoic acid in promoting the differentiation of 3T3-L1 preadipocytes. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-K.; Della-Fera, M.A.; Lin, J.; Baile, C.A. Docosahexaenoic Acid Inhibits Adipocyte Differentiation and Induces Apoptosis in 3T3-L1 Preadipocytes. J. Nutr. 2006, 136, 2965–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Grasa-López, A.; Miliar-García, Á.; Quevedo-Corona, L.; Paniagua-Castro, N.; Escalona-Cardoso, G.; Reyes-Maldonado, E.; Jaramillo-Flores, M.-E. Undaria pinnatifida and Fucoxanthin Ameliorate Lipogenesis and Markers of Both Inflammation and Cardiovascular Dysfunction in an Animal Model of Diet-Induced Obesity. Mar. Drugs 2016, 14, 148. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cui, Y.; Hu, X.; Liao, X.; Zhang, Y. Chlorophyll supplementation in early life prevents diet-induced obesity and modulates gut microbiota in mice. Mol. Nutr. Food Res. 2019, 63, 1801219. [Google Scholar] [CrossRef]
- Jung, H.A.; Jung, H.J.; Jeong, H.Y.; Kwon, H.J.; Kim, M.S.; Choi, J.S. Anti-adipogenic activity of the edible brown alga Ecklonia stolonifera and its constituent fucosterol in 3T3-L1 adipocytes. Arch Pharm. Res. 2014, 37, 713–720. [Google Scholar] [CrossRef]
- Slavin, J.L. Dietary fibers and body weight. Nutrition 2005, 21, 411–418. [Google Scholar] [CrossRef]
- Nuño, K.; Villarruel-López, A.; Pueblaperez, A.M.; Romerovelarde, E.; Puebla-Mora, A.; Ascencio, F. Effects of the marine microalgae Isochrysis galbana and Nannochloropsis oculata in diabetic rats. J. Funct. Foods 2013, 5, 106–115. [Google Scholar] [CrossRef]
- Anderson, J.W.; Jones, A.E.; Riddell-Mason, S. Ten Different Dietary Fibers Have Significantly Different Effects on Serum and Liver Lipids of Cholesterol-Fed Rats. J. Nutr. 1994, 124, 78–83. [Google Scholar] [CrossRef]
- Ostlund, R.E. Phytosterols in human nutrition. Annu. Rev. Nutr. 2002, 22, 533–549. [Google Scholar] [CrossRef]
- Egert, S.; Kannenberg, F.; Somoza, V.; Erbersdobler, H.F.; Wahrburg, U. Dietary α-Linolenic Acid, EPA, and DHA Have Differential Effects on LDL Fatty Acid Composition but Similar Effects on Serum Lipid Profiles in Normolipidemic Humans. J. Nutr. 2009, 139, 861–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, M.-H.; Jia, Y.; Jun, H.-J.; Lee, J.H.; Lee, B.Y.; Lee, S.-J. Fucosterol Is a Selective Liver X Receptor Modulator That Regulates the Expression of Key Genes in Cholesterol Homeostasis in Macrophages, Hepatocytes, and Intestinal Cells. J. Agric. Food Chem. 2012, 60, 11567–11575. [Google Scholar] [CrossRef] [PubMed]
- Threapleton, D.E.; Greenwood, D.C.; Evans, C.E.; Cleghorn, C.L.; Nykjaer, C.; Woodhead, C.; Cade, J.E.; Gale, C.P.; Burley, V.J. Dietary fiber intake and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2013, 347, f6879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRorie, J.; McKeown, N.M. Understanding the Physics of Functional Fibers in the Gastrointestinal Tract: An Evidence-Based Approach to Resolving Enduring Misconceptions about Insoluble and Soluble Fiber. J. Acad. Nutr. Diet. 2017, 117, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, N.; Sano, K.; Horiuchi, A.; Tanaka, E.; Kiyosawa, K.; Aoyama, T. Highly Purified Eicosapentaenoic Acid Treatment Improves Nonalcoholic Steatohepatitis. J. Clin. Gastroenterol. 2008, 42, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Eliasson, B.; Gudbjörnsdottir, S.; Zethelius, B.; Eeg-Olofsson, K.; Cederholm, J. LDL-cholesterol versus non-HDL-to-HDL-cholesterol ratio and risk for coronary heart disease in type 2 diabetes. Eur. J. Prev. Cardiol. 2014, 21, 1420–1428. [Google Scholar] [CrossRef]
- Adhyaru, B.B.; Jacobson, T.A. New Cholesterol Guidelines for the Management of Atherosclerotic Cardiovascular Disease Risk. Endocrinol. Metab. Clin. N. Am. 2016, 45, 17–37. [Google Scholar] [CrossRef]
- Shen, S.-W.; Lu, Y.; Li, F.; Yang, C.-J.; Feng, Y.-B.; Li, H.-W.; Yao, W.-F.; Shen, Z.-H. Atherogenic index of plasma is an effective index for estimating abdominal obesity. Lipids Heal. Dis. 2018, 17, 1–6. [Google Scholar] [CrossRef] [Green Version]
- De Jesus Raposo, M.; de Morais, A.; De Morais, R.M. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef]
- Luo, X.; Su, P.; Zhang, W. Advances in Microalgae-Derived Phytosterols for Functional Food and Pharmaceutical Applications. Mar. Drugs 2015, 13, 4231–4254. [Google Scholar] [CrossRef]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of polysaccharides in food, digestion, and health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanai, H.; Masui, Y.; Katsuyama, H.; Adachi, H.; Kawaguchi, A.; Hakoshima, M.; Waragai, Y.; Harigae, T.; Sako, A. An Improvement of Cardiovascular Risk Factors by Omega-3 Polyunsaturated Fatty Acids. J. Clin. Med. Res. 2018, 10, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Dietary modification of inflammation with lipids. Proc. Nutr. Soc. 2002, 61, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ma, Y.; Sun, J.; Ran, L.; Li, Y.; Wang, N.; Yu, T.; Gao, W.; Jia, W.; Jiang, R.; et al. Microalgal Oil fromSchizochytriumsp. Prevents HFD-Induced Abdominal Fat Accumulation in Mice. J. Am. Coll. Nutr. 2017, 36, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Abdul, Q.A.; Choi, R.J.; Jung, H.A.; Choi, J.S. Health benefit of fucosterol from marine algae: A review: Health benefit of fucosterol. J. Sci. Food Agric. 2016, 96, 1856–1866. [Google Scholar] [CrossRef]
- Fernando, I.S.; Nah, J.-W.; Jeon, Y.-J. Potential anti-inflammatory natural products from marine algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef]
- Tan, C.-P.; Hou, Y.-H. First Evidence for the Anti-inflammatory Activity of Fucoxanthin in High-Fat-Diet-Induced Obesity in Mice and the Antioxidant Functions in PC12 Cells. Inflammation 2013, 37, 443–450. [Google Scholar] [CrossRef]
- Bellenger, J.; Bellenger, S.; Escoula, Q.; Bidu, C.; Narce, M. N-3 polyunsaturated fatty acids: An innovative strategy against obesity and related metabolic disorders, intestinal alteration and gut microbiota dysbiosis. Biochimie 2019, 159, 66–71. [Google Scholar] [CrossRef]
- Reilly, S.M.; Saltiel, A.R. Adapting to obesity with adipose tissue inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Moraes, A.D.S.; Pisani, L.; Corgosinho, F.; Carvalho, L.T.; Masquio, D.; Jamar, G.; Sanches, R.B.; Oyama, L.; Dâmaso, A.; Belote, C.; et al. The Role of Leptinemia State as a Mediator of Inflammation in Obese Adults. Horm. Metab. Res. 2013, 45, 605–610. [Google Scholar] [CrossRef]
- Yook, J.-S.; Kim, K.-A.; Park, J.E.; Lee, S.-H.; Cha, Y.-S. Microalgal Oil Supplementation Has an Anti-Obesity Effect in C57BL/6J Mice Fed a High Fat Diet. Prev. Nutr. Food Sci. 2015, 20, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.-J.; Kim, S.M.; Jeong, S.-M.; Choi, H.-N.; Jang, Y.-H.; Kim, J.-I. Antioxidant effect of Phaeodactylum tricornutum in mice fed high-fat diet. Food Sci. Biotechnol. 2013, 22, 107–113. [Google Scholar] [CrossRef]
- Noguchi, N.; Konishi, F.; Kumamoto, S.; Maruyama, I.; Ando, Y.; Yanagita, T. Beneficial effects of Chlorella on glucose and lipid metabolism in obese rodents on a high-fat diet. Obes. Res. Clin. Pr. 2013, 7, e95–e105. [Google Scholar] [CrossRef] [PubMed]
- Vecina, J.F.; Oliveira, A.G.; Araujo, T.G.; Baggio, S.R.; Torello, C.O.; Saad, M.J.A.; Queiroz, M.L.D.S. Chlorella modulates insulin signaling pathway and prevents high-fat diet-induced insulin resistance in mice. Life Sci. 2014, 95, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Nasirian, F.; Sarir, H.; Moradi-Kor, N. Antihyperglycemic and antihyperlipidemic activities of Nannochloropsis oculata microalgae in Streptozotocin-induced diabetic rats. Biomol. Concepts 2019, 10, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Chen, L.; Wang, Z.; Zheng, X.; Hou, Z.; Zhao, D.; Long, J.; Liu, J. Omega-3 polyunsaturated fatty acids prevent obesity by improving tricarboxylic acid cycle homeostasis. J. Nutr. Biochem. 2021, 88, 108503. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary Combination of Fucoxanthin and Fish Oil Attenuates the Weight Gain of White Adipose Tissue and Decreases Blood Glucose in Obese/Diabetic KK-AyMice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Dupas, J.; Goanvec, C.; Feray, A.; Guernec, A.; Alain, C.; Guerrero, F.; Mansourati, J. Progressive Induction of Type 2 Diabetes: Effects of a Reality–Like Fructose Enriched Diet in Young Wistar Rats. PLoS ONE 2016, 11, e0146821. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, E.; Bandy, A.; VanWagner, L.B. Diagnostic challenges of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Clin. Liver Dis. 2018, 11, 98–104. [Google Scholar] [CrossRef] [Green Version]
- De Mello-Sampayo, C.; Paterna, A.; Polizzi, A.; Duarte, D.; Batista, I.; Pinto, R.; Gonçalves, P.; Raymundo, A.; Batista, A.P.; Gouveia, L.; et al. Evaluation of marine microalga Diacronema vlkianum biomass fatty acid assimilation in Wistar Rats. Molecules 2017, 22, 1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.; Huang, J.; Lee, B.-K.; Jung, Y.-S.; Im, E.; Koh, J.-M.; Im, D.-S. Omega-3 polyunsaturated fatty acids protect human hepatoma cells from developing steatosis through FFA4 (GPR120). Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2018, 1863, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.-N.; Jeon, S.-M.; Kim, H.-J.; Lee, M.-K.; Shin, S.-K.; Shin, Y.C.; Park, Y.-B.; Choi, M.-S. Fucoxanthin supplementation improves plasma and hepatic lipid metabolism and blood glucose concentration in high-fat fed C57BL/6N mice. Chem. Interact. 2010, 186, 316–322. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.; Borggreve, S.E.; Dullaart, R.P. Role of lipases, lecithin:cholesterol acyltransferase and cholesteryl ester transfer protein in abnormal high density lipoprotein metabolism in insulin resistance and type 2 diabetes mellitus. Clin. Lab. 2003, 49, 601–613. [Google Scholar]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochime 2004, 86, 839–848. [Google Scholar] [CrossRef]
- Tong, L. Acetyl-coenzyme A carboxylase: Crucial metabolic enzyme and attractive target for drug discovery. Cell. Mol. Life Sci. 2005, 62, 1784–1803. [Google Scholar] [CrossRef]
- Gammone, M.; D’Orazio, N. Anti-obesity activity of the marine carote-noid fucoxanthin. Mar. Drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Kanthimathi, M.S.; Tan, A.T.-B.; Nesaretnam, K.; Teng, K.-T. The amount and types of fatty acids acutely affect insulin, glycemic and gastrointestinal peptide responses but not satiety in metabolic syndrome subjects. Eur. J. Nutr. 2016, 57, 179–190. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Chen, Y.-L.; Huang, W.-C.; Liou, C.-J. Fucoxanthin attenuates fatty acid-induced lipid accumulation in FL83B hepatocytes through regulated Sirt1/AMPK signaling pathway. Biochem. Biophys. Res. Commun. 2018, 495, 197–203. [Google Scholar] [CrossRef]
 ), control group; HF (
), control group; HF (  ), high-fat group; HF-Tiso (
), high-fat group; HF-Tiso (  ), the high-fat group supplemented with T. lutea. Values are means (n = 6), with standard deviations represented by vertical bars. Statistical significance was determined using ANOVA with post hoc Fisher’s test, and means associated with letters indicate significative difference at p < 0.05 (week 1, Figure 1c) and p < 0.001 with a > b > c.
), control group; HF ( ), high-fat group; HF-Tiso ( ), the high-fat group supplemented with T. lutea. Values are means (n = 6), with standard deviations represented by vertical bars. Statistical significance was determined using ANOVA with post hoc Fisher’s test, and means associated with letters indicate significative difference at p < 0.05 (week 1, Figure 1c) and p < 0.001 with a > b > c.
), the high-fat group supplemented with T. lutea. Values are means (n = 6), with standard deviations represented by vertical bars. Statistical significance was determined using ANOVA with post hoc Fisher’s test, and means associated with letters indicate significative difference at p < 0.05 (week 1, Figure 1c) and p < 0.001 with a > b > c.
), control group; HF ( ), high-fat group; HF-Tiso ( ), the high-fat group supplemented with T. lutea. Values are means (n = 6), with standard deviations represented by vertical bars. Statistical significance was determined using ANOVA with post hoc Fisher’s test, and means associated with letters indicate significative difference at p < 0.05 (week 1, Figure 1c) and p < 0.001 with a > b > c.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTRL | CTRL | HF | HF-Tiso |
|---|---|---|---|
| Plasma biochemical parameters | |||
| ASAT (UI/L) | 58.72 ± 6.22 | 51.80 *** ± 7.83 | 45.09 ***† ± 5.39 |
| ALAT (UI/L) | 44.46 ± 4.57 | 52.10 *** ± 4.10 | 52.00 *** ± 6.79 |
| ASAT/ALAT | 1.36 ± 0.11 | 1.24 *** ± 0.17 | 0.87 ***† ± 0.09 |
| Glucose (mmol/L) | 9.17 ± 1.48 | 10.01 ***† ± 0.58 | 7.73 ***† ± 0.10 |
| Insulin (µUI/L) | 37.68 ± 8.26 | 89.17 *** ± 12.95 | 64.41 ***† ± 11.69 |
| Leptin (ng/mL) | 2.02 ± 0.52 | 3.87 *** ± 0.53 | 2.17 † ± 0.30 |
| HOMA-IR | 0.97 ± 0.34 | 2.80 *** ± 0.57 | 1.30 ***† ± 0.33 |
| TAG (mmol/L) | 3.07 ± 0.84 | 6.85 *** ± 1.16 | 3.53 † ± 1.02 |
| TC (mmol/L) | 2.14 ± 0.36 | 2.72 *** ± 0.45 | 3.64 ***† ± 0.35 |
| HDL-C (mmol/L) | 2.03 ± 0.27 | 1.72 *** ± 0.20 | 2.83 ***† ± 0.26 |
| LDL-C (mmol/L) | 0.44 ± 0.12 | 0.96 *** ± 0.21 | 0.86 *** ± 0.19 |
| HDL/LDL | 2.65 ± 0.31 | 1.62 *** ± 0.39 | 2.73 † ± 0.44 |
| AIP | 0.32 ± 0.08 | 0.51 *** ± 0.13 | 0.18 ***† ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayer, C.; Richard, L.; Côme, M.; Ulmann, L.; Nazih, H.; Chénais, B.; Ouguerram, K.; Mimouni, V. The Marine Microalga, Tisochrysis lutea, Protects against Metabolic Disorders Associated with Metabolic Syndrome and Obesity. Nutrients 2021, 13, 430. https://doi.org/10.3390/nu13020430
Mayer C, Richard L, Côme M, Ulmann L, Nazih H, Chénais B, Ouguerram K, Mimouni V. The Marine Microalga, Tisochrysis lutea, Protects against Metabolic Disorders Associated with Metabolic Syndrome and Obesity. Nutrients. 2021; 13(2):430. https://doi.org/10.3390/nu13020430
Chicago/Turabian StyleMayer, Claire, Léo Richard, Martine Côme, Lionel Ulmann, Hassan Nazih, Benoît Chénais, Khadija Ouguerram, and Virginie Mimouni. 2021. "The Marine Microalga, Tisochrysis lutea, Protects against Metabolic Disorders Associated with Metabolic Syndrome and Obesity" Nutrients 13, no. 2: 430. https://doi.org/10.3390/nu13020430
APA StyleMayer, C., Richard, L., Côme, M., Ulmann, L., Nazih, H., Chénais, B., Ouguerram, K., & Mimouni, V. (2021). The Marine Microalga, Tisochrysis lutea, Protects against Metabolic Disorders Associated with Metabolic Syndrome and Obesity. Nutrients, 13(2), 430. https://doi.org/10.3390/nu13020430