
In Vivo Evaluation of Dendropanax morbifera Leaf Extract for Anti-Obesity and Cholesterol-Lowering Activity in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of D. morbifera Leaf Extracts
2.2. Animal Experiment
2.3. Histological Examination
2.4. Plasma Biochemistry
2.5. Determination of SOD Activity
2.6. Determination of Lipid Peroxidation
2.7. Measurement of Nitric Oxide Activity
2.8. Limulus Amebocyte Lysate Assays
2.9. RNA Extraction and Real-Time Quantitative PCR Analyses
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
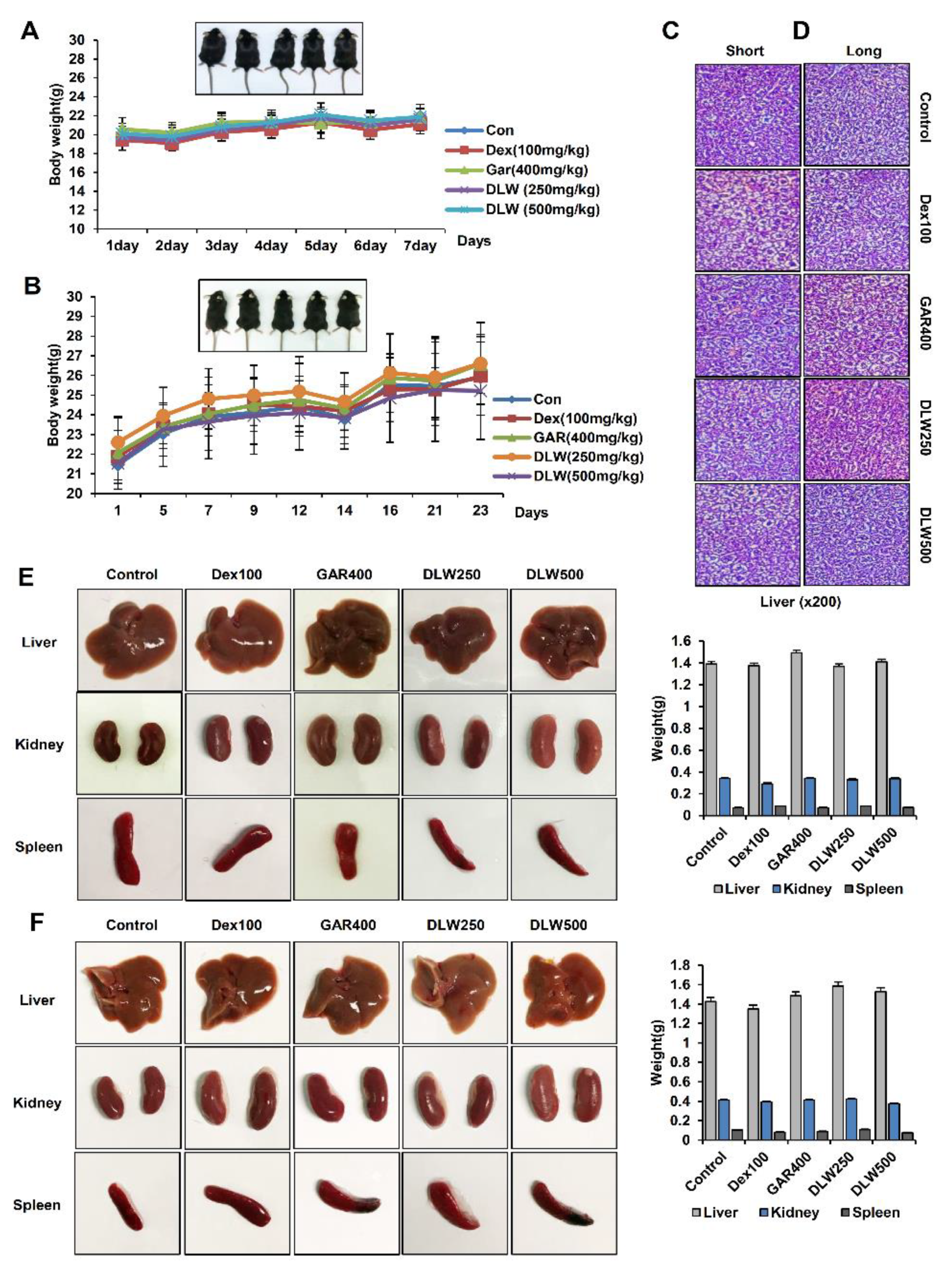
3.1. In Vivo Determine the Dosage and Toxicity of DLW Extracts
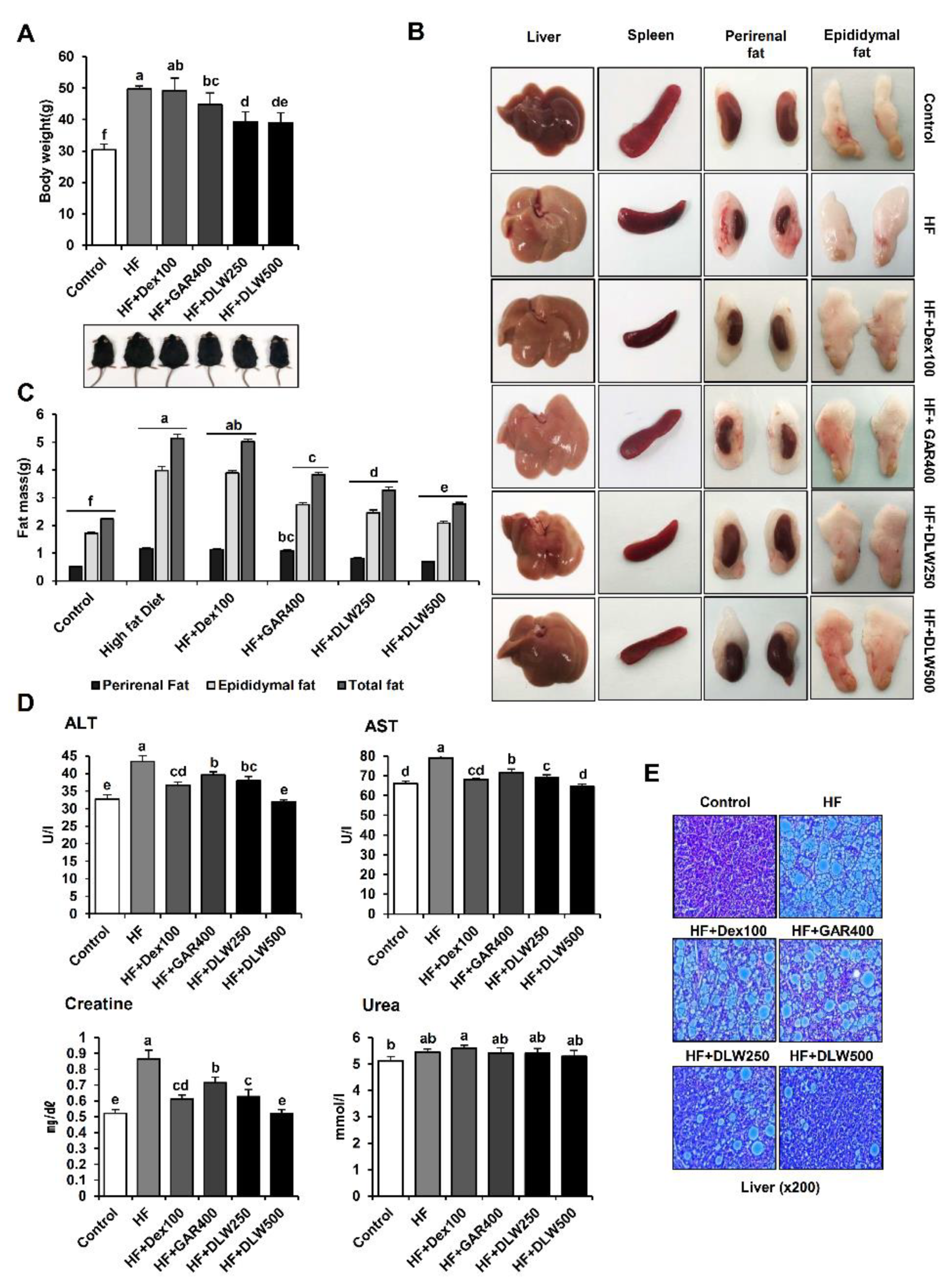
3.2. Anti-Obesity Function of DLW Extracts In Vivo
3.3. DLW Extract Affects Oxidative Stress and Cholesterol Level in the Tissue, Plasma, Urine
3.4. DLW Extract Regulates AMPK, ACC-1, CPT1, ATGL, and DGAT1 Expression Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zoccali, C.; Mallamaci, F.; Tripepi, G. Adipose tissue as a source of inflammatory cytokines in health and disease: Focus on end-stage renal disease. Kidney Int. Suppl. 2003, 63, S65–S68. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPalpha induces adipogenesis through PPARgamma: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Payne, V.A.; Au, W.S.; Lowe, C.E.; Rahman, S.M.; Friedman, J.E.; O’Rahilly, S.; Rochford, J.J. C/EBP transcription factors regulate SREBP1c gene expression during adipogenesis. Biochem. J. 2009, 425, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Nguyen, V.B.; Dong, J.; Wang, Y.; Park, J.Y.; Lee, S.C.; Yang, T.J. Evolution of the Araliaceae family inferred from complete chloroplast genomes and 45S nrDNAs of 10 Panax-related species. Sci. Rep. 2017, 7, 4917. [Google Scholar] [CrossRef]
- Kregiel, D.; Berlowska, J.; Witonska, I.; Antolak, H.; Proestos, C.; Babic, M.; Babic, L.; Zhang, B. Saponin-Based, Biological-Active Surfactants from Plants; SW7 2QJ, UNITED KINGDOM; InTech: London, UK, 2017; pp. 183–204. [Google Scholar]
- Song, J.H.; Kang, H.B.; Kim, J.H.; Kwak, S.; Sung, G.J.; Park, S.H.; Jeong, J.H.; Kim, H.; Lee, J.; Jun, W.; et al. Antiobesity and Cholesterol-Lowering Effects of Dendropanax morbifera Water Extracts in Mouse 3T3-L1 Cells. J. Med. Food 2018, 21, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, R.; Cho, D.Y.; Su-Kim, I.; Choi, D.K. Dendropanax Morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology. Antioxidants 2020, 9, 962. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Kim, S.H.; Kwon, C.; Kim, S.Y.; Yang, Y.J.; Kim, J.S.; Ali, M.; Ahmad, A. New Chemical Constituents from the Bark of Dendropanax morbifera Leveille and their Evaluation of Antioxidant Activities. Molecules 2019, 24, 3967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choo, G.S.; Lim, D.P.; Kim, S.M.; Yoo, E.S.; Kim, S.H.; Kim, C.H.; Woo, J.S.; Kim, H.J.; Jung, J.Y. Antiinflammatory effects of Dendropanax morbifera in lipopolysaccharidestimulated RAW264.7 macrophages and in an animal model of atopic dermatitis. Mol. Med. Rep. 2019, 19, 2087–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Zhao, X.; Xu, H.; Chen, F.; Xu, Y.; Li, Z.; Sanchis, D.; Jin, L.; Zhang, Y.; Ye, J. AKT2 Blocks Nucleus Translocation of Apoptosis-Inducing Factor (AIF) and Endonuclease G (EndoG) While Promoting Caspase Activation during Cardiac Ischemia. Int. J. Mol. Sci. 2017, 18, 565. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Ryu, H.K. Effects of Dendropanax morbifera Leaf Extracts on Lipid Profiles in Mice Fed a High-Fat and High-Cholesterol Diet. J. Korean Soc. Food Sci. Nutr. 2015, 44, 641–648. [Google Scholar] [CrossRef]
- O’Rahilly, S.; Farooqi, I.S. Human obesity: A heritable neurobehavioral disorder that is highly sensitive to environmental conditions. Diabetes 2008, 57, 2905–2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Kim, M.-S.; Chun, S.-S.; Choi, J.-H. Effects of Turmeric (Curcuma longa L.) on Antioxidative Systems and Oxidative Damage in Rats Fed a High Fat and Cholesterol Diet. J. Korean Soc. Food Sci. Nutr. 2013, 42, 570–576. [Google Scholar] [CrossRef] [Green Version]
- Song, W.-Y.; Yang, J.-A.; Ku, K.-H.; Choi, J.-H. Effect of Red Pepper Seeds Powder on Antioxidative System and Oxidative Damage in Rats Fed High-Fat.High-Cholesterol Diet. J. Korean Soc. Food Sci. Nutr. 2009, 38, 1161–1166. [Google Scholar] [CrossRef]
- Patlevic, P.; Vaskova, J.; Svorc, P., Jr.; Vasko, L.; Svorc, P. Reactive oxygen species and antioxidant defense in human gastrointestinal diseases. Integr. Med. Res. 2016, 5, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd Esa, N.; Abdul Kadir, K.K.; Amom, Z.; Azlan, A. Antioxidant activity of white rice, brown rice and germinated brown rice (in vivo and in vitro) and the effects on lipid peroxidation and liver enzymes in hyperlipidaemic rabbits. Food Chem. 2013, 141, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Gotoh, T. Regulation of nitric oxide production by arginine metabolic enzymes. Biochem. Biophys. Res. Commun. 2000, 275, 715–719. [Google Scholar] [CrossRef]
- Jequier, E.; Constant, F. Water as an essential nutrient: The physiological basis of hydration. Eur. J. Clin. Nutr. 2010, 64, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Afonso, M.S.; Machado, R.M.; Lavrador, M.S.; Quintao, E.C.R.; Moore, K.J.; Lottenberg, A.M. Molecular Pathways Underlying Cholesterol Homeostasis. Nutrients 2018, 10, 760. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, K.; Murakami, T.; Sakurai, T.; Miyoshi, M.; Kurahashi, K.; Kishi, S.; Tamaki, M.; Tominaga, T.; Yoshida, S.; Nagai, K.; et al. Circulating Apolipoprotein L1 is associated with insulin resistance-induced abnormal lipid metabolism. Sci. Rep. 2019, 9, 14869. [Google Scholar] [CrossRef]
- Zhou, L.; Li, C.; Gao, L.; Wang, A. High-density lipoprotein synthesis and metabolism (Review). Mol. Med. Rep. 2015, 12, 4015–4021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss. Arter. Thromb. Vasc. Biol. 2006, 26, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lai, X.; Sun, L.; Cao, J.; Ling, C.; Zhang, W.; Xiang, L.; Chen, R.; Li, D.; Sun, S. Antiobesity and anti-inflammation effects of Hakka stir-fried tea of different storage years on high-fat diet-induced obese mice model via activating the AMPK/ACC/CPT1 pathway. Food Nutr. Res. 2020, 64, 1681. [Google Scholar] [CrossRef] [PubMed]
- Bruce, C.R.; Hoy, A.J.; Turner, N.; Watt, M.J.; Allen, T.L.; Carpenter, K.; Cooney, G.J.; Febbraio, M.A.; Kraegen, E.W. Overexpression of carnitine palmitoyltransferase-1 in skeletal muscle is sufficient to enhance fatty acid oxidation and improve high-fat diet-induced insulin resistance. Diabetes 2009, 58, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Haemmerle, G.; Lass, A.; Zimmermann, R.; Gorkiewicz, G.; Meyer, C.; Rozman, J.; Heldmaier, G.; Maier, R.; Theussl, C.; Eder, S.; et al. Defective lipolysis and altered energy metabolism in mice lacking adipose triglyceride lipase. Science 2006, 312, 734–737. [Google Scholar] [CrossRef]
- Zechner, R.; Zimmermann, R.; Eichmann, T.O.; Kohlwein, S.D.; Haemmerle, G.; Lass, A.; Madeo, F. FAT SIGNALS--lipases and lipolysis in lipid metabolism and signaling. Cell Metab. 2012, 15, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.C. Enhancing energy and glucose metabolism by disrupting triglyceride synthesis: Lessons from mice lacking DGAT1. Nutr. Metab. (Lond.) 2006, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr Hypertens Rep 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [Green Version]
- Childs, B.G.; Durik, M.; Baker, D.J.; van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef]
- Foretz, M.; Even, P.C.; Viollet, B. AMPK Activation Reduces Hepatic Lipid Content by Increasing Fat Oxidation In Vivo. Int. J. Mol. Sci. 2018, 19, 2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.F.; Infante, J.R. Molecular Pathways: Fatty Acid Synthase. Clin. Cancer Res. 2015, 21, 5434–5438. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.-H.; Kim, H.; Jeong, M.; Kong, M.J.; Choi, H.-K.; Jun, W.; Kim, Y.; Choi, K.-C. In Vivo Evaluation of Dendropanax morbifera Leaf Extract for Anti-Obesity and Cholesterol-Lowering Activity in Mice. Nutrients 2021, 13, 1424. https://doi.org/10.3390/nu13051424
Song J-H, Kim H, Jeong M, Kong MJ, Choi H-K, Jun W, Kim Y, Choi K-C. In Vivo Evaluation of Dendropanax morbifera Leaf Extract for Anti-Obesity and Cholesterol-Lowering Activity in Mice. Nutrients. 2021; 13(5):1424. https://doi.org/10.3390/nu13051424
Chicago/Turabian StyleSong, Ji-Hye, Hyunhee Kim, Minseok Jeong, Min Jung Kong, Hyo-Kyoung Choi, Woojin Jun, Yongjae Kim, and Kyung-Chul Choi. 2021. "In Vivo Evaluation of Dendropanax morbifera Leaf Extract for Anti-Obesity and Cholesterol-Lowering Activity in Mice" Nutrients 13, no. 5: 1424. https://doi.org/10.3390/nu13051424
APA StyleSong, J. -H., Kim, H., Jeong, M., Kong, M. J., Choi, H. -K., Jun, W., Kim, Y., & Choi, K. -C. (2021). In Vivo Evaluation of Dendropanax morbifera Leaf Extract for Anti-Obesity and Cholesterol-Lowering Activity in Mice. Nutrients, 13(5), 1424. https://doi.org/10.3390/nu13051424