
Metabolism and Bioavailability of Olive Bioactive Constituents Based on In Vitro, In Vivo and Human Studies



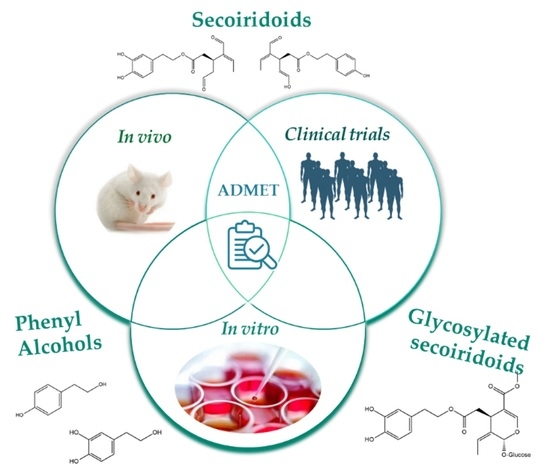
Abstract
:
1. Introduction
2. Materials and Methods
3. Results
3.1. Bibliography Overview
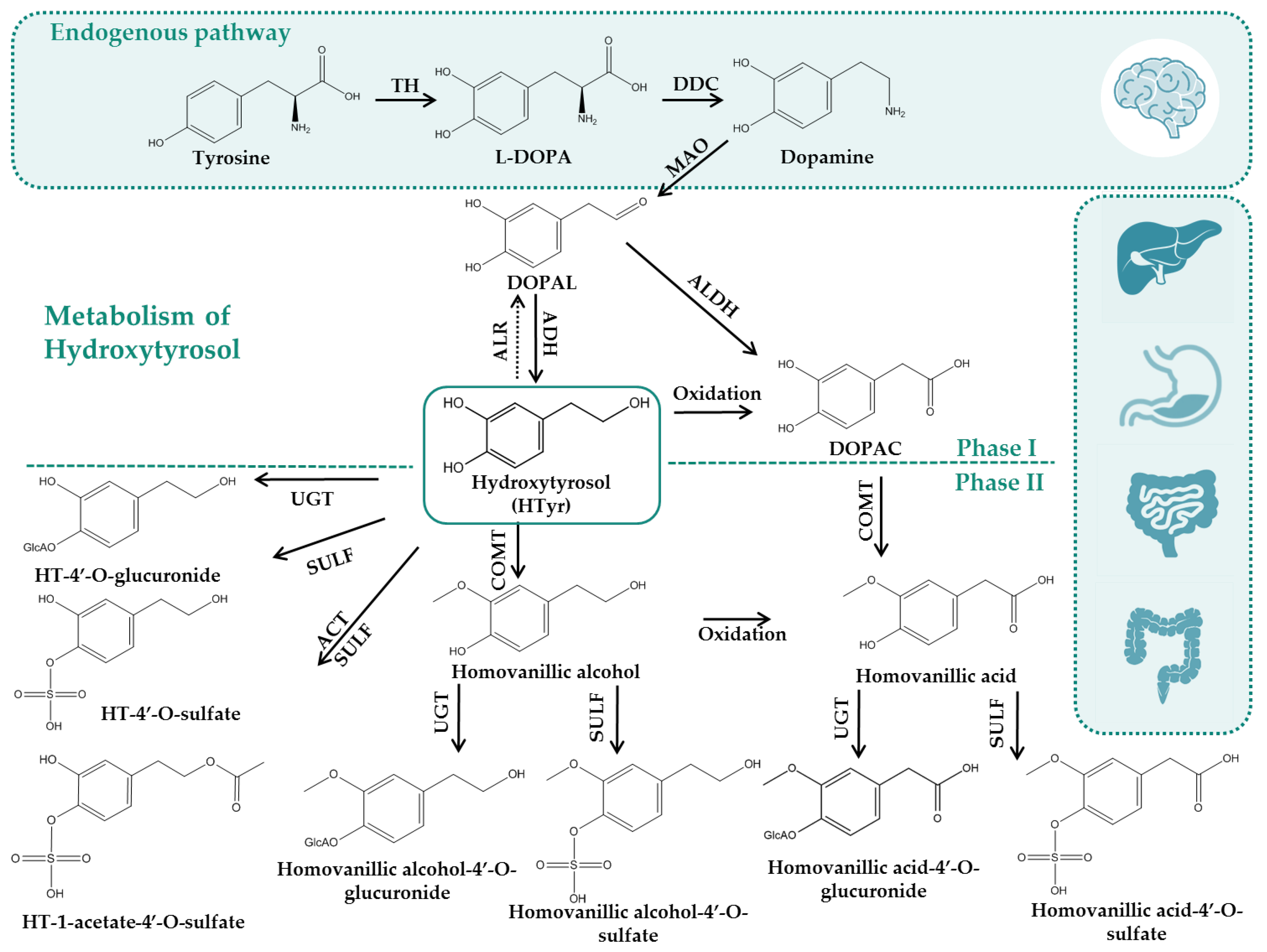
3.2. ADMET of Hydroxytyrosol (HTyr) and Tyrosol (Tyr)
3.2.1. In Vitro Bioavailabilty and Metabolism Assays of Hydroxytyrosol (Htyr)
3.2.2. In Vitro Bioavailability and Metabolism Assays of Tyrosol (Tyr)
3.2.3. In Vivo Assays for ADMET Properties of Hydroxytyrosol (HTyr)
3.2.4. In Vivo Assays for ADMET Properties of Tyrosol (Tyr)
3.3. ADMET of Secoiridoids Oleacein (Olea), Oleocanthal (Oleo) and Oleuropein (Oleu)
3.3.1. In Vitro Bioavailability and Metabolism Assays of Oleacein (Olea), Oleocanthal (Oleo) and Oleuropein (Oleu)
3.3.2. In Vivo Assays of Oleacein (Olea), Oleocanthal (Oleo) and Oleuropein (Oleu)
3.4. Human Trials for Phenylalcohols and Secoiridoids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Visioli, F.; Davalos, A.; de las Hazas, M.C.L.; Crespo, M.C.; Tomé-Carneiro, J. An overview of the pharmacology of olive oil and its active ingredients. Br. J. Pharmacol. 2020, 177, 1316–1330. [Google Scholar] [CrossRef] [PubMed]
- Nikou, T.; Witt, M.; Stathopoulos, P.; Barsch, A.; Halabalaki, M. Olive Oil Quality and Authenticity Assessment Aspects Employing FIA-MRMS and LC-Orbitrap MS Metabolomic Approaches. Front. Public Health 2020, 8, 534. [Google Scholar] [CrossRef] [PubMed]
- Angelis, A.; Hamzaoui, M.; Aligiannis, N.; Nikou, T.; Michailidis, D.; Gerolimatos, P.; Termentzi, A.; Hubert, J.; Halabalaki, M.; Renault, J.-H.; et al. An integrated process for the recovery of high added-value compounds from olive oil using solid support free liquid-liquid extraction and chromatography techniques. J. Chromatogr. A 2017, 1491, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-López, P.; Lozano-Sanchez, J.; Borrás-Linares, I.; Emanuelli, T.; Menéndez, J.A.; Segura-Carretero, A. Structure–biological activity relationships of extra-virgin olive oil phenolic compounds: Health properties and bioavailability. Antioxidants 2020, 9, 685. [Google Scholar] [CrossRef] [PubMed]
- Acar-Tek, N.; Ağagündüz, D. Olive Leaf (Olea europaea L. folium): Potential Effects on Glycemia and Lipidemia. Ann. Nutr. Metab. 2020, 76, 10–15. [Google Scholar] [CrossRef]
- Silenzi, A.; Giovannini, C.; Scazzocchio, B.; Varì, R.; D’Archivio, M.; Santangelo, C.; Masella, R. Extra virgin olive oil polyphenols: Biological properties and antioxidant activity. In Pathology; Academic Press: Cambridge, MA, USA, 2020; pp. 225–233. [Google Scholar] [CrossRef]
- Gorzynik-Debicka, M.; Przychodzen, P.; Cappello, F.; Kuban-Jankowska, A.; Gammazza, A.M.; Knap, N.; Wozniak, M.; Gorska-Ponikowska, M. Potential health benefits of olive oil and plant polyphenols. Int. J. Mol. Sci. 2018, 19, 686. [Google Scholar] [CrossRef]
- Bulotta, S.; Celano, M.; Lepore, S.M.; Montalcini, T.; Pujia, A.; Russo, D. Beneficial effects of the olive oil phenolic components oleuropein and hydroxytyrosol: Focus on protection against cardiovascular and metabolic diseases. J. Transl. Med. 2014, 12, 219. [Google Scholar] [CrossRef]
- Carrera-González, M.P.; Ramírez-Expósito, M.J.; Mayas, M.D.; Martínez-Martos, J.M. Protective role of oleuropein and its metabolite hydroxytyrosol on cancer. Trends Food Sci. Technol. 2013, 31, 92–99. [Google Scholar] [CrossRef]
- Parkinson, L.; Cicerale, S. The health benefiting mechanisms of virgin olive oil phenolic compounds. Molecules 2016, 21, 1734. [Google Scholar] [CrossRef] [Green Version]
- Soldevila-Domenech, N.; Boronat, A.; Mateus, J.; Diaz-Pellicer, P.; Matilla, I.; Pérez-Otero, M.; Aldea-Perona, A.; De La Torre, R. Generation of the antioxidant hydroxytyrosol from tyrosol present in beer and red wine in a randomized clinical trial. Nutrients 2019, 11, 2241. [Google Scholar] [CrossRef]
- Rodríguez-Morató, J.; Boronat, A.; Kotronoulas, A.; Pujadas, M.; Pastor, A.; Olesti, E.; Pérez-Mañá, C.; Khymenets, O.; Fitó, M.; Farré, M.; et al. Metabolic disposition and biological significance of simple phenols of dietary origin: Hydroxytyrosol and tyrosol. Drug Metab. Rev. 2016, 48, 218–236. [Google Scholar] [CrossRef] [PubMed]
- Castañer, O.; Covas, M.-I.; Khymenets, O.; Nyyssonen, K.; Konstantinidou, V.; Zunft, H.-F.; de la Torre, R.; Muñoz-Aguayo, D.; Vila, J.; Fitó, M. Protection of LDL from oxidation by olive oil polyphenols is associated with a downregulation of CD40-ligand expression and its downstream products in vivo in humans. Am. J. Clin. Nutr. 2012, 95, 1238–1244. [Google Scholar] [CrossRef]
- Moreno-Luna, R.; Muñoz-Hernandez, R.; Miranda, M.L.; Costa, A.F.; Jimenez-Jimenez, L.; Vallejo-Vaz, A.J.; Muriana, F.J.G.; Villar, J.; Stiefel, P. Olive Oil Polyphenols Decrease Blood Pressure and Improve Endothelial Function in Young Women with Mild Hypertension. Am. J. Hypertens. 2012, 25, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, N.; Ma, Y.; Wen, D. Hydroxytyrosol improves obesity and insulin resistance by modulating gut microbiota in high-fat diet-induced obese mice. Front. Microbiol. 2019, 10, 390. [Google Scholar] [CrossRef]
- Hernáez, Á.; Fernández-Castillejo, S.; Farràs, M.; Catalán, Ú.; Subirana, I.; Montes, R.; Solà, R.; Muñoz-Aguayo, D.; Gelabert-Gorgues, A.; Díaz-Gil, Ó.; et al. Olive oil polyphenols enhance high-density lipoprotein function in humans: A randomized controlled trial. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2115–2119. [Google Scholar] [CrossRef]
- Susalit, E.; Agus, N.; Effendi, I.; Tjandrawinata, R.R.; Nofiarny, D.; Perrinjaquet-Moccetti, T.; Verbruggen, M. Olive (Olea europaea) leaf extract effective in patients with stage-1 hypertension: Comparison with Captopril. Phytomedicine 2011, 18, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Violi, F.; Loffredo, L.; Pignatelli, P.; Angelico, F.; Bartimoccia, S.; Nocella, C.; Cangemi, R.; Petruccioli, A.; Monticolo, R.; Pastori, D.; et al. Extra virgin olive oil use is associated with improved post-prandial blood glucose and LDL cholesterol in healthy subjects. Nutr. Diabetes 2015, 5, e172. [Google Scholar] [CrossRef] [PubMed]
- Bonura, A.; Vlah, S.; Longo, A.; Bulati, M.; Melis, M.R.; Cibella, F.; Colombo, P. Hydroxytyrosol modulates Par j 1-induced IL-10 production by PBMCs in healthy subjects. Immunobiology 2016, 221, 1374–1377. [Google Scholar] [CrossRef]
- Del Río, L.F.; Gutiérrez-Casado, E.; Varela-López, A.; Villalba, J.M. Olive oil and the hallmarks of aging. Molecules 2016, 21, 163. [Google Scholar] [CrossRef] [Green Version]
- Lockyer, S.; Yaqoob, P.; Spencer, J.P.E.; Rowland, I. Olive leaf phenolics and cardiovascular risk reduction: Physiological effects and mechanisms of action. Nutr. Aging 2012, 1, 125–140. [Google Scholar] [CrossRef]
- Markellos, C.; Ourailidou, M.-E.; Gavriatopoulou, M.; Halvatsiotis, P.; Sergentanis, T.N.; Psaltopoulou, T. Olive oil intake and cancer risk: A systematic review and meta-analysis. PLoS ONE 2022, 17, e0261649. [Google Scholar] [CrossRef] [PubMed]
- Sirangelo, I.; Borriello, M.; Liccardo, M.; Scafuro, M.; Russo, P.; Iannuzzi, C. Hydroxytyrosol Selectively Affects Non-Enzymatic Glycation in Human Insulin and Protects by AGEs Cytotoxicity. Antioxidants 2021, 10, 1127. [Google Scholar] [CrossRef] [PubMed]
- de Bock, M.; Derraik, J.G.B.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) Leaf Polyphenols Improve Insulin Sensitivity in Middle-Aged Overweight Men: A Randomized, Placebo-Controlled, Crossover Trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef] [PubMed]
- Fytili, C.; Nikou, T.; Tentolouris, N.; Tseti, I.K.; Dimosthenopoulos, C.; Sfikakis, P.P.; Simos, D.; Kokkinos, A.; Skaltsounis, A.L.; Katsilambros, N.; et al. Effect of Long-Term Hydroxytyrosol Administration on Body Weight, Fat Mass and Urine Metabolomics: A Randomized Double-Blind Prospective Human Study. Nutrients 2022, 14, 1525. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Morze, J.; Hoffmann, G. Mediterranean diet and health status: Active ingredients and pharmacological mechanisms. Br. J. Pharmacol. 2020, 177, 1241–1257. [Google Scholar] [CrossRef]
- Cicerale, S.; Lucas, L.J.; Keast, R.S.J. Antimicrobial, antioxidant and anti-inflammatory phenolic activities in extra virgin olive oil. Curr. Opin. Biotechnol. 2012, 23, 129–135. [Google Scholar] [CrossRef]
- Serreli, G.; Deiana, M. Biological relevance of extra virgin olive oil polyphenols metabolites. Antioxidants 2018, 7, 170. [Google Scholar] [CrossRef]
- De La Torre, R. Bioavailability of olive oil phenolic compounds in humans. Inflammopharmacology 2008, 16, 245–247. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588. [Google Scholar] [CrossRef] [Green Version]
- Guan, L.; Yang, H.; Cai, Y.; Sun, L.; Di, P.; Li, W.; Liu, G.; Tang, Y. ADMET-score—A comprehensive scoring function for evaluation of chemical drug-likeness. Medchemcomm 2019, 10, 148–157. [Google Scholar] [CrossRef]
- Borah, P.; Hazarika, S.; Deka, S.; Venugopala, K.N.; Nair, A.B.; Attimarad, M.; Sreeharsha, N.; Mailavaram, R.P. Application of Advanced Technologies in Natural Product Research: A Review with Special Emphasis on ADMET Profiling. Curr. Drug Metab. 2020, 21, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Paine, M.F. Natural Products: Experimental Approaches to Elucidate Disposition Mechanisms and Predict Pharmacokinetic Drug Interactions. Drug Metab. Dispos. 2020, 48, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Galmés, S.; Reynés, B.; Palou, M.; Palou-March, A.; Palou, A. Absorption, Distribution, Metabolism, and Excretion of the Main Olive Tree Phenols and Polyphenols: A Literature Review. J. Agric. Food Chem. 2021, 69, 5281–5296. [Google Scholar] [CrossRef] [PubMed]
- Ooka, M.; Lynch, C.; Xia, M. Application of In Vitro Metabolism Activation in High-Throughput Screening. Int. J. Mol. Sci. 2020, 21, 8182. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Minihane, A.-M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef]
- Koppel, N.; Rekdal, V.M.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356, 1246–1257. [Google Scholar] [CrossRef]
- Cheng, F.; Li, W.; Liu, G.; Tang, Y. In Silico ADMET Prediction: Recent Advances, Current Challenges and Future Trends. Curr. Top. Med. Chem. 2013, 13, 1273–1289. [Google Scholar] [CrossRef]
- Reichel, A.; Lienau, P. Pharmacokinetics in Drug Discovery: An Exposure-Centred Approach to Optimising and Predicting Drug Efficacy and Safety. Handb. Exp. Pharmacol. 2016, 232, 235–260. [Google Scholar] [CrossRef]
- Derendorf, H. Pharmacokinetics of Natural Compounds. Planta Med. 2012, 78, IL41. [Google Scholar] [CrossRef]
- Hughes, J.P.; Rees, S.; Kalindjian, S.B.; Philpott, K.L. Principles of early drug discovery. Br. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef]
- González-Santiago, M.; Fonollá, J.; Lopez-Huertas, E. Human absorption of a supplement containing purified hydroxytyrosol, a natural antioxidant from olive oil, and evidence for its transient association with low-density lipoproteins. Pharmacol. Res. 2010, 61, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Piroddi, M.; Albini, A.; Fabiani, R.; Giovannelli, L.; Luceri, C.; Natella, F.; Rosignoli, P.; Rossi, T.; Taticchi, A.; Servili, M.; et al. Nutrigenomics of extra-virgin olive oil: A review. BioFactors 2017, 43, 17–41. [Google Scholar] [CrossRef] [PubMed]
- Landberg, R.; Manach, C.; Kerckhof, F.-M.; Minihane, A.-M.; Saleh, R.N.M.; De Roos, B.; Tomas-Barberan, F.; Morand, C.; Van de Wiele, T. Future prospects for dissecting inter-individual variability in the absorption, distribution and elimination of plant bioactives of relevance for cardiometabolic endpoints. Eur. J. Nutr. 2019, 58, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, E.; Sarria, B.; Espartero, J.L.; Correa, J.A.G.; Bravo-Clemente, L.; Mateos, R. Evaluation of the Bioavailability and Metabolism of Nitroderivatives of Hydroxytyrosol Using Caco-2 and HepG2 Human Cell Models. J. Agric. Food Chem. 2016, 64, 2289–2297. [Google Scholar] [CrossRef]
- Suárez, M.; Romero, M.P.; Macià, A.; Valls, R.M.; Fernández, S.; Solà, R.; Motilva, M.J. Improved method for identifying and quantifying olive oil phenolic compounds and their metabolites in human plasma by microelution solid-phase extraction plate and liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 4097–4106. [Google Scholar] [CrossRef]
- Corona, G.; Spencer, J.; Dessì, M. Extra virgin olive oil phenolics: Absorption, metabolism, and biological activities in the GI tract. Toxicol. Ind. Health 2009, 25, 285–293. [Google Scholar] [CrossRef]
- de la Torre-Carbot, K.; Chávez-Servín, J.L.; Jaúregui, O.; Castellote, A.I.; Lamuela-Raventós, R.M.; Fitó, M.; Covas, M.I.; Muñoz-Aguayo, D.; López-Sabater, M.C. Presence of virgin olive oil phenolic metabolites in human low density lipoprotein fraction: Determination by high-performance liquid chromatography-electrospray ionization tandem mass spectrometry. Anal. Chim. Acta 2007, 583, 402–410. [Google Scholar] [CrossRef]
- Tuck, K.L.; Hayball, P.J. Major phenolic compounds in olive oil: Metabolism and health effects. J. Nutr. Biochem. 2002, 13, 636–644. [Google Scholar] [CrossRef]
- Visioli, F.; Galli, C.; Galli, G.; Caruso, D. Biological activities and metabolic fate of olive oil phenols. Eur. J. Lipid Sci. Technol. 2002, 104, 677–684. [Google Scholar] [CrossRef]
- Vissers, M.N.; Zock, P.L.; Katan, M.B. Bioavailability and antioxidant effects of olive oil phenols in humans: A review. Eur. J. Clin. Nutr. 2004, 58, 955–965. [Google Scholar] [CrossRef]
- Mosele, J.I.; Martín-Peláez, S.; Macià, A.; Farràs, M.; Valls, R.M.; Catalán, Ú.; Motilva, M.J. Faecal microbial metabolism of olive oil phenolic compounds: In vitro and in vivo approaches. Mol. Nutr. Food Res. 2014, 58, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Marković, A.K.; Torić, J.; Barbarić, M.; Brala, C.J. Hydroxytyrosol, tyrosol and derivatives and their potential effects on human health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef] [PubMed]
- Martínez, L.; Ros, G.; Nieto, G. Hydroxytyrosol: Health Benefits and Use as Functional Ingredient in Meat. Medicines 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Panel, E.; Nda, A. Scientific Opinion on the substantiation of health claims related to polyphenols in olive and protection of LDL particles from oxidative damage (ID 1333, 1638, 1639, 1696, 2865), maintenance of normal blood HDL cholesterol concentrations (ID 1639), mainte. EFSA J. 2011, 9, 2033. [Google Scholar] [CrossRef]
- Manna, C.; Galletti, P.; Maisto, G.; Cucciolla, V.; D’Angelo, S.; Zappia, V. Transport mechanism and metabolism of olive oil hydroxytyrosol in Caco-2 cells. FEBS Lett. 2000, 470, 341–344. [Google Scholar] [CrossRef]
- Corona, G.; Tzounis, X.; Dessì, M.A.; Deiana, M.; Debnam, E.S.; Visioli, F.; Spencer, J.P.E. The fate of olive oil polyphenols in the gastrointestinal tract: Implications of gastric and colonic microflora-dependent biotransformation. Free Radic. Res. 2006, 40, 647–658. [Google Scholar] [CrossRef]
- Soler, A.; Romero, M.P.; Macià, A.; Saha, S.; Furniss, C.S.M.; Kroon, P.A.; Motilva, M.J. Digestion stability and evaluation of the metabolism and transport of olive oil phenols in the human small-intestinal epithelial Caco-2/TC7 cell line. Food Chem. 2010, 119, 703–714. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Mateos, R.; Saha, S.; Madrona, A.; Espartero, J.L.; Bravo, L.; Kroon, P.A. Transepithelial transport and metabolism of new lipophilic ether derivatives of hydroxytyrosol by enterocyte-like Caco-2/TC7 cells. J. Agric. Food Chem. 2010, 58, 11501–11509. [Google Scholar] [CrossRef]
- Mateos, R.; Pereira-Caro, G.; Saha, S.; Cert, R.; Redondo-Horcajo, M.; Bravo, L.; Kroon, P.A. Acetylation of hydroxytyrosol enhances its transport across differentiated Caco-2 cell monolayers. Food Chem. 2011, 125, 865–872. [Google Scholar] [CrossRef]
- Keservani, R.K.; Kesharwani, R.K.; Sharma, A.K.; Gautam, S.P.; Verma, S.K. Chapter 9—Nutraceutical Formulations and Challenges. In Developing New Functional Food and Nutraceutical Products; Bagchi, D., Nair, S., Eds.; Academic Press: San Diego, CA, USA, 2017; pp. 161–177. ISBN 978-0-12-802780-6. [Google Scholar]
- Rubió, L.; Macià, A.; Castell-Auví, A.; Pinent, M.; Blay, M.T.; Ardévol, A.; Romero, M.P.; Motilva, M.J. Effect of the co-occurring olive oil and thyme extracts on the phenolic bioaccesibility and bioavailability assessed by in vitro digestion and cell models. Food Chem. 2014, 149, 277–284. [Google Scholar] [CrossRef]
- Malapert, A.; Tomao, V.; Dangles, O.; Reboul, E. Effect of Foods and β-Cyclodextrin on the Bioaccessibility and the Uptake by Caco-2 Cells of Hydroxytyrosol from Either a Pure Standard or Alperujo. J. Agric. Food Chem. 2018, 66, 4614–4620. [Google Scholar] [CrossRef] [PubMed]
- Malapert, A.; Tomao, V.; Margier, M.; Nowicki, M.; Gleize, B.; Dangles, O.; Reboul, E. Β-Cyclodextrin Does Not Alter the Bioaccessibility and the Uptake By Caco-2 Cells of Olive By-Product Phenolic Compounds. Nutrients 2018, 10, 1653. [Google Scholar] [CrossRef] [PubMed]
- Mitsou, E.; Dupin, A.; Sassi, A.H.; Monteil, J.; Sotiroudis, G.T.; Leal-Calderon, F.; Xenakis, A. Hydroxytyrosol encapsulated in biocompatible water-in-oil microemulsions: How the structure affects in vitro absorption. Colloids Surf. B Biointerfaces 2019, 184, 110482. [Google Scholar] [CrossRef] [PubMed]
- Mateos, R.; Goya, L.; Bravo, L. Metabolism of the olive oil phenols hydroxytyrosol, tyrosol, and hydroxytyrosyl acetate by human hepatoma HepG2 cells. J. Agric. Food Chem. 2005, 53, 9897–9905. [Google Scholar] [CrossRef] [PubMed]
- Goya, L.; Mateos, R.; Martín, M.A.; Ramos, S.; Bravo, L. Uptake, Metabolism and Biological Effect of the Olive Oil Phenol Hydroxytyrosol in Human HepG2 Cells. In Olives and Olive Oil in Health and Disease Prevention; Elsevier Inc.: Amsterdam, The Netherlands, 2010; pp. 1157–1165. ISBN 9780123744203. [Google Scholar]
- Mosele, J.I.; Macià, A.; Motilva, M.J. Metabolic and microbial modulation of the large intestine ecosystem by non-absorbed diet phenolic compounds: A review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef]
- Deiana, M.; Serra, G.; Corona, G. Modulation of intestinal epithelium homeostasis by extra virgin olive oil phenolic compounds. Food Funct. 2018, 9, 4085–4099. [Google Scholar] [CrossRef]
- D’Angelo, S.; Manna, C.; Migliardi, V.; Mazzoni, O.; Morrica, P.; Capasso, G.; Pontoni, G.; Galletti, P.; Zappia, V. Pharmacokinetics and metabolism of hydroxytyrosol, a natural antioxidant from olive oil. Drug Metab. Dispos. 2001, 29, 1492–1498. [Google Scholar]
- Sakavitsi, M.E.; Breynaert, A.; Nikou, T.; Lauwers, S.; Pieters, L.; Hermans, N.; Halabalaki, M. Availability and Metabolic Fate of Olive Phenolic Alcohols Hydroxytyrosol and Tyrosol in the Human GI Tract Simulated by the In Vitro GIDM-Colon Model. Metabolites 2022, 12, 391. [Google Scholar] [CrossRef]
- Visioli, F.; Caruso, D.; Plasmati, E.; Patelli, R.; Mulinacci, N.; Romani, A.; Galli, G.; Galli, C. Hydroxytyrosol, as a component of olive mill waste water, is dose-dependently absorbed and increases the antioxidant capacity of rat plasma. Free Radic. Res. 2001, 34, 301–305. [Google Scholar] [CrossRef]
- Domínguez-Perles, R.; Auñón, D.; Ferreres, F.; Gil-Izquierdo, A. Gender differences in plasma and urine metabolites from Sprague–Dawley rats after oral administration of normal and high doses of hydroxytyrosol, hydroxytyrosol acetate, and DOPAC. Eur. J. Nutr. 2017, 56, 215–224. [Google Scholar] [CrossRef]
- Bai, C.; Yan, X.; Takenaka, M.; Sekiya, K.; Nagata, T. Determination of Synthetic Hydroxytyrosol in Rat Plasma by GC-MS. J. Agric. Food Chem. 1998, 46, 3998–4001. [Google Scholar] [CrossRef]
- de las Hazas, M.C.L.; Piñol, C.; Macià, A.; Romero, M.P.; Pedret, A.; Solà, R.; Rubió, L.; Motilva, M.J. Differential absorption and metabolism of hydroxytyrosol and its precursors oleuropein and secoiridoids. J. Funct. Foods 2016, 22, 52–63. [Google Scholar] [CrossRef]
- de las Hazas, M.C.L.; Godinho-Pereira, J.; Macià, A.; Almeida, A.F.; Ventura, M.R.; Motilva, M.J.; Santos, C.N. Brain uptake of hydroxytyrosol and its main circulating metabolites: Protective potential in neuronal cells. J. Funct. Foods 2018, 46, 110–117. [Google Scholar] [CrossRef]
- Kotronoulas, A.; Pizarro, N.; Serra, A.; Robledo, P.; Joglar, J.; Rubió, L.; Hernaéz, Á.; Tormos, C.; Motilva, M.J.; Fitó, M.; et al. Dose-dependent metabolic disposition of hydroxytyrosol and formation of mercapturates in rats. Pharmacol. Res. 2013, 77, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Egashira, Y.; Sanada, H. Ferulic acid is quickly absorbed from rat stomach as the free form and then conjugated mainly in liver. J. Nutr. 2004, 134, 3083–3088. [Google Scholar] [CrossRef]
- Ruiz-Gutiérrez, V.; Juan, M.E.; Cert, A.; Planas, J.M. Determination of hydroxytyrosol in plasma by HPLC. Anal. Chem. 2000, 72, 4458–4461. [Google Scholar] [CrossRef]
- Rodríguez-Gutiérrez, G.; Wood, S.; Guzmán, J.F.-B.; Duthie, G.G.; de Roos, B. Determination of 3,4-dihydroxyphenylglycol, hydroxytyrosol and tyrosol purified from olive oil by-products with HPLC in animal plasma and tissues. Food Chem. 2011, 126, 1948–1952. [Google Scholar] [CrossRef]
- Serra, A.; Rubió, L.; Borràs, X.; Macià, A.; Romero, M.P.; Motilva, M.J. Distribution of olive oil phenolic compounds in rat tissues after administration of a phenolic extract from olive cake. Mol. Nutr. Food Res. 2012, 56, 486–496. [Google Scholar] [CrossRef]
- de las Hazas, M.C.L.; Rubió, L.; Kotronoulas, A.; de la Torre, R.; Solà, R.; Motilva, M.J. Dose effect on the uptake and accumulation of hydroxytyrosol and its metabolites in target tissues in rats. Mol. Nutr. Food Res. 2015, 59, 1395–1399. [Google Scholar] [CrossRef]
- Goldstein, D.S.; Jinsmaa, Y.; Sullivan, P.; Holmes, C.; Kopin, I.J.; Sharabi, Y. 3,4-Dihydroxyphenylethanol (Hydroxytyrosol) Mitigates the Increase in Spontaneous Oxidation of Dopamine During Monoamine Oxidase Inhibition in PC12 Cells. Neurochem. Res. 2016, 41, 2173–2178. [Google Scholar] [CrossRef] [Green Version]
- Auñon-Calles, D.; Canut, L.; Visioli, F. Toxicological evaluation of pure hydroxytyrosol. Food Chem. Toxicol. 2013, 55, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Christian, M.S.; Sharper, V.A.; Hoberman, A.M.; Seng, J.E.; Fu, L.; Covell, D.; Diener, R.M.; Bitler, C.M.; Crea, R. The toxicity profile of hydrolyzed aqueous olive pulp extract. Drug Chem. Toxicol. 2004, 27, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Soni, M.G.; Burdock, G.A.; Christian, M.S.; Bitler, C.M.; Crea, R. Safety assessment of aqueous olive pulp extract as an antioxidant or antimicrobial agent in foods. Food Chem. Toxicol. 2006, 44, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Heilman, J.; Anyangwe, N.; Tran, N.; Edwards, J.; Beilstein, P.; López, J. Toxicological evaluation of an olive extract, H35: Subchronic toxicity in the rat. Food Chem. Toxicol. 2015, 84, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Robles-Almazan, M.; Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Rodriguez-Garcia, C.; Quiles, J.L.; Ramirez-Tortosa, M. Hydroxytyrosol: Bioavailability, toxicity, and clinical applications. Food Res. Int. 2018, 105, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, Y.J.; Kim, M.J.; Ahn, J.; Ha, T.Y.; Lee, S.H.; Jang, Y.J.; Jung, C.H. Pharmacokinetics of tyrosol metabolites in rats. Molecules 2016, 21, 128. [Google Scholar] [CrossRef]
- Lozano-Castellón, J.; López-Yerena, A.; de Alvarenga, J.F.R.; del Castillo-Alba, J.R.; Vallverdú-Queralt, A.; Escribano-Ferrer, E.; Lamuela-Raventós, R.M. Health-promoting properties of oleocanthal and oleacein: Two secoiridoids from extra-virgin olive oil. Crit. Rev. Food Sci. Nutr. 2020, 60, 2532–2548. [Google Scholar] [CrossRef]
- Guo, N.; Zhu, M.; Han, X.; Sui, D.; Wang, Y.; Yang, Q. The metabolism of salidroside to its aglycone p-tyrosol in rats following the administration of salidroside. PLoS ONE 2014, 9, 103648. [Google Scholar] [CrossRef]
- Buckland, G.; Gonzalez, C.A. The role of olive oil in disease prevention: A focus on the recent epidemiological evidence from cohort studies and dietary intervention trials. Br. J. Nutr. 2015, 113, S94–S101. [Google Scholar] [CrossRef]
- Beauchamp, G.K.; Keast, R.S.J.; Morel, D.; Lin, J.; Pika, J.; Han, Q.; Lee, C.H.; Smith, A.B.; Breslin, P.A.S. Ibuprofen-like activity in extra-virgin olive oil. Nature 2005, 437, 45–46. [Google Scholar] [CrossRef]
- Celano, M.; Maggisano, V.; Lepore, S.M.; Russo, D.; Bulotta, S. Secoiridoids of olive and derivatives as potential coadjuvant drugs in cancer: A critical analysis of experimental studies. Pharmacol. Res. 2019, 142, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.L.; Chin, K.Y. The biological activities of oleocanthal from a molecular perspective. Nutrients 2018, 10, 570. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Garcia-Aloy, M.; Figueira, M.E.; Combet, E.; Mullen, W.; Bronze, M.R. High Resolution Mass Spectrometric Analysis of Secoiridoids and Metabolites as Biomarkers of Acute Olive Oil Intake—An Approach to Study Interindividual Variability in Humans. Mol. Nutr. Food Res. 2018, 62, 1700065. [Google Scholar] [CrossRef] [PubMed]
- Achat, S.; Tomao, V.; Madani, K.; Chibane, M.; Elmaataoui, M.; Dangles, O.; Chemat, F. Direct enrichment of olive oil in oleuropein by ultrasound-assisted maceration at laboratory and pilot plant scale. Ultrason. Sonochem. 2012, 19, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Hassen, I.; Casabianca, H.; Hosni, K. Biological activities of the natural antioxidant oleuropein: Exceeding the expectation—A mini-review. J. Funct. Foods 2015, 18, 926–940. [Google Scholar] [CrossRef]
- Romani, A.; Ieri, F.; Urciuoli, S.; Noce, A.; Marrone, G.; Nediani, C.; Bernini, R. Health effects of phenolic compounds found in extra-virgin olive oil, by-products, and leaf of Olea europaea L. Nutrients 2019, 11, 1776. [Google Scholar] [CrossRef]
- Ray, N.B.; Hilsabeck, K.D.; Karagiannis, T.C.; McCord, D.E. Bioactive olive oil polyphenols in the promotion of health. The Role of Functional Food Security in Global Health; Academic Press: Cambridge, MA, USA, 2018; pp. 623–637. [Google Scholar] [CrossRef]
- Özcan, M.M.; Matthäus, B. A review: Benefit and bioactive properties of olive (Olea europaea L.) leaves. Eur. Food Res. Technol. 2017, 243, 89–99. [Google Scholar] [CrossRef]
- Del Boccio, P.; Di Deo, A.; De Curtis, A.; Celli, N.; Iacoviello, L.; Rotilio, D. Liquid chromatography-tandem mass spectrometry analysis of oleuropein and its metabolite hydroxytyrosol in rat plasma and urine after oral administration. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2003, 785, 47–56. [Google Scholar] [CrossRef]
- Tan, H.W.; Tuck, K.L.; Stupans, I.; Hayball, P.J. Simultaneous determination of oleuropein and hydroxytyrosol in rat plasma using liquid chromatography with fluorescence detection. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2003, 785, 187–191. [Google Scholar] [CrossRef]
- Žugčić, T.; Abdelkebir, R.; Alcantara, C.; Collado, M.C.; García-Pérez, J.V.; Meléndez-Martínez, A.J.; Jambrak, A.R.; Lorenzo, J.M.; Barba, F.J. From extraction of valuable compounds to health promoting benefits of olive leaves through bioaccessibility, bioavailability and impact on gut microbiota. Trends Food Sci. Technol. 2019, 83, 63–77. [Google Scholar] [CrossRef]
- Reboredo-Rodríguez, P.; Olmo-García, L.; Figueiredo-González, M.; González-Barreiro, C.; Carrasco-Pancorbo, A.; Cancho-Grande, B. Application of the INFOGEST Standardized Method to Assess the Digestive Stability and Bioaccessibility of Phenolic Compounds from Galician Extra-Virgin Olive Oil. J. Agric. Food Chem. 2021, 69, 11592–11605. [Google Scholar] [CrossRef] [PubMed]
- Edgecombe, S.C.; Stretch, G.L.; Hayball, P.J. Oleuropein, an antioxidant polyphenol from olive oil, is poorly absorbed from isolated perfused rat intestine. J. Nutr. 2000, 130, 2996–3002. [Google Scholar] [CrossRef]
- Kendall, M.; Batterham, M.; Prenzler, P.D.; Ryan, D.; Robards, K. Absorption, metabolism and excretion of phenolics derived from olive products. Funct. Plant Sci. Biotechnol. 2009, 3, 81–91. [Google Scholar]
- Franc, I.; Lipinski, A.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar]
- Paiva-Martins, F.; Gordon, M.H.; Gameiro, P. Activity and location of olive oil phenolic antioxidants in liposomes. Chem. Phys. Lipids 2003, 124, 23–36. [Google Scholar] [CrossRef]
- Pinto, J.; Paiva-Martins, F.; Corona, G.; Debnam, E.S.; Oruna-Concha, M.J.; Vauzour, D.; Gordon, M.H.; Spencer, J.P.E. Absorption and metabolism of olive oil secoiridoids in the small intestine. Br. J. Nutr. 2011, 105, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Galli, C.; Grande, S.; Colonnelli, K.; Patelli, C.; Galli, G. Hydroxytyrosol excretion differs between rats and humans and depends on the vehicle of administration. J. Nutr. 2003, 133, 2612–2615. [Google Scholar] [CrossRef]
- Kano, S.; Komada, H.; Yonekura, L.; Sato, A.; Nishiwaki, H.; Tamura, H. Absorption, Metabolism, and Excretion by Freely Moving Rats of 3,4-DHPEA-EDA and Related Polyphenols from Olive Fruits (Olea europaea). J. Nutr. Metab. 2016, 2016, 9104208. [Google Scholar] [CrossRef]
- Bazoti, F.N.; Gikas, E.; Tsarbopoulos, A. Simultaneous quantification of oleuropein and its metabolites in rat plasma by liquid chromatography electrospray ionization tandem mass spectrometry. Biomed. Chromatogr. 2009, 24, 506–515. [Google Scholar] [CrossRef]
- Tuck, K.L.; Freeman, M.P.; Hayball, P.J.; Stretch, G.L.; Stupans, I. The in vivo fate of hydroxytyrosol and tyrosol, antioxidant phenolic constituents of olive oil, after intravenous and oral dosing of labeled compounds to rats. J. Nutr. 2001, 131, 1993–1996. [Google Scholar] [CrossRef]
- Lin, P.; Qian, W.; Wang, X.; Cao, L.; Li, S.; Qian, T. The biotransformation of oleuropein in rats. Biomed. Chromatogr. 2013, 27, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Qian, T.; Wang, X.; Li, X. Application of LC-MS/MS method for the in vivo metabolite determination of oleuropein after intravenous administration to rat. Biomed. Chromatogr. 2011, 25, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, L.; Keast, R. Oleocanthal, a Phenolic Derived from Virgin Olive Oil: A Review of the Beneficial Effects on Inflammatory Disease. Int. J. Mol. Sci. 2014, 15, 12323–12334. [Google Scholar] [CrossRef] [PubMed]
- López-Yerena, A.; Vallverdú-Queralt, A.; Mols, R.; Augustijns, P.; Lamuela-Raventós, R.M.; Escribano-Ferrer, E. Absorption and intestinal metabolic profile of oleocanthal in rats. Pharmaceutics 2020, 12, 34. [Google Scholar] [CrossRef]
- López-Yerena, A.; Vallverdú-Queralt, A.; Jáuregui, O.; Garcia-Sala, X.; Lamuela-Raventós, R.M.; Escribano-Ferrer, E. Tissue Distribution of Oleocanthal and Its Metabolites after Oral Ingestion in Rats. Antioxidants 2021, 10, 688. [Google Scholar] [CrossRef]
- Dressman, J.B.; Thelen, K. Cytochrome P450-mediated metabolism in the human gut wall. J. Pharm. Pharmacol. 2009, 61, 541–558. [Google Scholar] [CrossRef]
- López-Yerena, A.; Pérez, M.; Vallverdú-Queralt, A.; Miliarakis, E.; Lamuela-Raventós, R.M.; Escribano-Ferrer, E. Oleacein Intestinal Permeation and Metabolism in Rats Using an In Situ Perfusion Technique. Pharmaceutics 2021, 13, 719. [Google Scholar] [CrossRef]
- Alemán-Jiménez, C.; Domínguez-Perles, R.; Medina, S.; Prgomet, I.; López-González, I.; Simonelli-Muñoz, A.; Campillo-Cano, M.; Auñón, D.; Ferreres, F.; Gil-Izquierdo, Á. Pharmacokinetics and bioavailability of hydroxytyrosol are dependent on the food matrix in humans. Eur. J. Nutr. 2021, 60, 905–915. [Google Scholar] [CrossRef]
- Visioli, F.; Galli, C.; Bornet, F.; Mattei, A.; Patelli, R.; Galli, G.; Caruso, D. Olive oil phenolics are dose-dependently absorbed in humans. FEBS Lett. 2000, 468, 159–160. [Google Scholar] [CrossRef]
- Suárez, M.; Valls, R.M.; Romero, M.P.; MacIà, A.; Fernández, S.; Giralt, M.; Solà, R.; Motilva, M.J. Bioavailability of phenols from a phenol-enriched olive oil. Br. J. Nutr. 2011, 106, 1691–1701. [Google Scholar] [CrossRef]
- Covas, M.I.; De La Torre, K.; Farré-Albaladejo, M.; Kaikkonen, J.; Fitó, M.; López-Sabater, C.; Pujadas-Bastardes, M.A.; Joglar, J.; Weinbrenner, T.; Lamuela-Raventós, R.M.; et al. Postprandial LDL phenolic content and LDL oxidation are modulated by olive oil phenolic compounds in humans. Free Radic. Biol. Med. 2006, 40, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Lamuela-Raventós, R.M.; Gimeno, E.; Fitó, M.; Castellote, A.I.; Covas, M.; De la Torre-Boronat, M.C.; López-Sabater, M.C. Interaction of olive oil phenol antioxidant components with low-density lipoprotein. Biol. Res. 2004, 37, 247–252. [Google Scholar] [CrossRef] [PubMed]
- de Bock, M.; Thorstensen, E.B.; Derraik, J.G.B.; Henderson, H.V.; Hofman, P.L.; Cutfield, W.S. Human absorption and metabolism of oleuropein and hydroxytyrosol ingested as olive (Olea europaea L.) leaf extract. Mol. Nutr. Food Res. 2013, 57, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- De La Torre-Carbot, K.; Chávez-Servín, J.L.; Jaúregui, O.; Castellote, A.I.; Lamuela-Raventós, R.M.; Nurmi, T.; Poulsen, H.E.; Gaddi, A.V.; Kaikkonen, J.; Zunft, H.F.; et al. Elevated circulating LDL phenol levels in men who consumed virgin rather than refined olive oil are associated with less oxidation of plasma LDL. J. Nutr. 2010, 140, 501–508. [Google Scholar] [CrossRef]
- Kendall, M.; Batterham, M.; Callahan, D.L.; Jardine, D.; Prenzler, P.D.; Robards, K.; Ryan, D. Randomized controlled study of the urinary excretion of biophenols following acute and chronic intake of olive leaf supplements. Food Chem. 2012, 130, 651–659. [Google Scholar] [CrossRef]
- Miro-Casas, E.; Covas, M.I.; Farre, M.; Fito, M.; Ortuño, J.; Weinbrenner, T.; Roset, P.; De La Torre, R. Hydroxytyrosol disposition in humans. Clin. Chem. 2003, 49, 945–952. [Google Scholar] [CrossRef]
- García-Villalba, R.; Larrosa, M.; Possemiers, S.; Tomás-Barberán, F.A.; Espín, J.C. Bioavailability of phenolics from an oleuropein-rich olive (Olea europaea) leaf extract and its acute effect on plasma antioxidant status: Comparison between pre- and postmenopausal women. Eur. J. Nutr. 2014, 53, 1015–1027. [Google Scholar] [CrossRef]
- Khymenets, O.; Crespo, M.C.; Dangles, O.; Rakotomanomana, N.; Andres-Lacueva, C.; Visioli, F. Human hydroxytyrosol’s absorption and excretion from a nutraceutical. J. Funct. Foods 2016, 23, 278–282. [Google Scholar] [CrossRef]
- Pérez-Mañá, C.; Farré, M.; Pujadas, M.; Mustata, C.; Menoyo, E.; Pastor, A.; Langohr, K.; De La Torre, R. Ethanol induces hydroxytyrosol formation in humans. Pharmacol. Res. 2015, 95–96, 27–33. [Google Scholar] [CrossRef]
- Vissers, M.N.; Zock, P.L.; Roodenburg, A.J.C.; Leenen, R.; Katan, M.B. Olive oil phenols are absorbed in humans. J. Nutr. 2002, 132, 409–417. [Google Scholar] [CrossRef]
- de la Torre-Carbot, K.; Jauregui, O.; Castellote, A.I.; Lamuela-Raventós, R.M.; Covas, M.I.; Casals, I.; López-Sabater, M.C. Rapid high-performance liquid chromatography-electrospray ionization tandem mass spectrometry method for qualitative and quantitative analysis of virgin olive oil phenolic metabolites in human low-density lipoproteins. J. Chromatogr. A 2006, 1116, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Rubió, L.; Farràs, M.; de La Torre, R.; Macià, A.; Romero, M.P.; Valls, R.M.; Solà, R.; Farré, M.; Fitó, M.; Motilva, M.J. Metabolite profiling of olive oil and thyme phenols after a sustained intake of two phenol-enriched olive oils by humans: Identification of compliance markers. Food Res. Int. 2014, 65, 59–68. [Google Scholar] [CrossRef]
- Fernández-Ávila, C.; Montes, R.; Castellote, A.I.; Chisaguano, A.M.; Fitó, M.; Covas, M.I.; Muñoz-Aguallo, D.; Nyyssönen, K.; Zunft, H.J.; López-Sabater, M.C. Fast determination of virgin olive oil phenolic metabolites in human high-density lipoproteins. Biomed. Chromatogr. 2015, 29, 1035–1041. [Google Scholar] [CrossRef]
- García-Villalba, R.; Carrasco-Pancorbo, A.; Nevedomskaya, E.; Mayboroda, O.A.; Deelder, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Exploratory analysis of human urine by LC-ESI-TOF MS after high intake of olive oil: Understanding the metabolism of polyphenols. Anal. Bioanal. Chem. 2010, 398, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Granados-Principal, S.; Quiles, J.L.; Ramirez-Tortosa, C.L.; Sanchez-Rovira, P.; Ramirez-Tortosa, M.C. Hydroxytyrosol: From laboratory investigations to future clinical trials. Nutr. Rev. 2010, 68, 191–206. [Google Scholar] [CrossRef]
- Estruch, R.; Martínez-González, M.A.; Corella, D.; Salas-Salvadó, J.; Ruiz-Gutierez, V.; Covas, M.I.; Fiol, M.; Gomez-Gracia, E.; Lopez-Sabater, M.C.; Vinyoles, E.; et al. Annals of Internal Medicine Article Effects of a Mediterranean-Style Diet on Cardiovascular Risk Factors. Ann. Intern. Med. 2006, 145, 1–11. [Google Scholar] [CrossRef]
- Pérez-Mañá, C.; Farré, M.; Rodríguez-Morató, J.; Papaseit, E.; Pujadas, M.; Fitó, M.; Robledo, P.; Covas, M.I.; Cheynier, V.; Meudec, E.; et al. Moderate consumption of wine, through both its phenolic compounds and alcohol content, promotes hydroxytyrosol endogenous generation in humans. A randomized controlled trial. Mol. Nutr. Food Res. 2015, 59, 1213–1216. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | |
| Language | English and Spanish |
| Sample size | No limit |
| Date of publication |
|
| Type of publication: | Research articles, reviews, short communications |
| Exclusion criteria | |
| Language | Other than English and Spanish |
| Type of publication | Conference abstracts, review meta-analyses, case reports, ecological studies, letters to editors, comments referring to published papers |
| Source of Olive Metabolites (Concentration) [Reference] | Detected or Measured Metabolites in Plasma | Detected or Measured Metabolites in Urine | Detected or Measured Metabolites in Feces | Results and Comments |
|---|---|---|---|---|
| OO (3.2 HTyr mg) and yogurt (20 mg Htyr)/orally [111] | (Not analyzed) | Htyr, HVAlc, | (Not analyzed) | Htyr excretion was much higher after its administration as a natural component of OO than after its addition to refined OO or yogurt. |
| Extra virgin OO, refined OO, flax oil, grapeseed oil, margarine, pineapple juice (5 mg of Htyr) [122] | (Not analyzed) | Htyr, Htyr acetate, Tyr, DOPAC, HVAlc | (Not analyzed) | The intake of extra virgin OO, as well as fortified refined olive, flax, and grapeseed oils provided significantly higher urinary contents in Htyr compared with basal urine, whereas Htyr metabolites showed no significant changes. No differences were found between men and women. |
| 30 mL virgin OO (Htyr, Tyr, HVA, p-coumaric, Olea, luteolin, pinoresinol, Oleo, acetoxypinoresinol, apigenin) [46] | Tyr-sulf, vanillin-sulf, Htyr-sulf, coumaric acid-sulf, vanillic acid-sulf, HVA-sulf, dihydroferulic acid-sulf, Htyr-glu, apigenin-glu | (Not analyzed) | (Not analyzed) | A μSPE-UPLC-ESI-MS/MS method was developed and applied to determine the phenolic compounds and their glucuronide, sulfate and methylated forms in human plasma after virgin OO ingestion. |
| OO (20 or 44 or 66 or 84 Htyr μg/mL and 36 or 72 or 110 or 140 Tyr μg/mL) [123] | (Not analyzed) | Htyr, Tyr | (Not analyzed) | Tyr and Htyr are dose-dependently absorbed in humans after ingestion and they are excreted in the urine as glucuronide conjugates. |
| Virgin OO or extra virgin OO (enriched in secoiridoids-89.4%, phenylalcohols-3.5% and flavonoids-6.0%) [124] | Phenyl alcohols and secoiridoids in sulfated and glucuronated forms | (Not analyzed) | (Not analyzed) | Compared with virgin OO, extra virgin OO increased plasma concentration of the phenol metabolites, particularly HTyr-sulf and vanillin-sulf. After the consumption of virgin OO, the maximum concentration of these peaks was reached at 60 min, while extra virgin OO shifted this maximum to 120 min. |
| HTyr (5 or 15 mg HTyr) as capsule [25] | (Not analyzed) | Hippuric acid, hydroxypippuric acid, epitestosterone-sulf, 5a-dihydrotestosterone-sulf, HVA, glutamine, HVA-sulf, p-cresol-sulf, 1,3-dimethyluric acid, homovanillic aldehyde-sulf. | (Not analyzed) | HTyr supplementation significantly affected the urine metabolome in a dose-dependent manner. |
| HTyr (2.5 mg/kg BW) as a supplement in an aqueous solution [42] | HTyr, HVAlc | HTyr, HVA, HVAlc, DOPAC in their sulfated and glucuronated form. | (Not analyzed) | The absorption of the HTyr from the supplement was very fast, as the plasma concentration showed maximum levels at 10 min after the administered dose. HTyr was excreted in urine mainly as sulfate and glucuronide conjugates. |
| 50 mL virigin OO (TPC 648 μg/mL, HTyr 70.6 μg/mL and Tyr 27.01 μg/mL) [48] | HTyr-glu, HTyr-sulf, Tyr-glu, Tyr-sulf, HVA-sulf | (Not analyzed) | (Not analyzed) | Quantitative methods were developed for the OO phenolic metabolites in human LDL |
| 40 mL OO (366 mg/kg or 164 mg/kg, or 2.7 mg/kg TPC) [125] | HTyr, Tyr and HVAlc, | (Not analyzed) | (Not analyzed) | Tyr and HTyr were dose-dependently absorbed. Total phenolic compounds in LDL increased in the postprandial state in a direct relationship with the phenolic compounds content of the OO ingested |
| 50 mL extra virgin OO [126] | Tyr and OO phenolics | (Not analyzed) | (Not analyzed) | This methodology allowed the demonstration of the in vitro binding capacity of Tyr |
| Olive leaf extract capsulated or liquid (Oleu-low dose 51.1 mg Oleu and 9.7 mg HTyr or Oleu-high 76.6 mg Oleu and 14.5 mg HTyr) [127] | HTyr-sulf, HTyr-glu, Oleu | HTyr-sulf, HTyr-glu, Oleu | (Not analyzed) | Gender’s effect on bioavailability, with males displaying greater plasma AUC for conjugated HTyr metabolites. All conjugated HTyr metabolites were recovered in the urine within 8 h. Oleu effectively delivers Oleu and HTyr metabolites to plasma in humans. |
| 25 mL of virgin OO (629 TPC mg/L), refined OO (0.0 TPC mg/L) [128] | HVA-sulf, HTyr-sulf, Tyr-sulf | (Not analyzed) | (Not analyzed) | The phenol concentration of OO modulates the phenolic metabolite content in LDL after sustained, daily consumption. The levels of LDL HTyr-sulf and HVA-sulf, but not of Tyr-sulf, were increased after virgin OO ingestion. |
| Leaf extract supplemented as capsule or liquid (20 mg Oleu/tablet and 22 mg Oleu/5 mL) [129] | (Not analyzed) | Oleu aglycone-glu, Oleu | (Not analyzed) | Following both chronic and acute ingestion, neither Oleu nor HTyr were detected in urine samples. However, Oleu aglycon-glu was detected in all urine samples up to 6 h following acute ingestion |
| 25 mL virgin OO (49.3 mg/L HTyr)/orally [130] | HTyr, HVAlc and their glucuronated conjugates | HTyr, HVAlc and their glucuronated conjugates | (Not analyzed) | HTyr and HVAlc were quantified in plasma after real-life doses of virgin OO. HTyr absorption was estimated at ∼98% in plasma and urine in conjugated forms, mainly glucuronated conjugates, suggesting extensive first-pass intestinal/hepatic metabolism of the ingested HTyr. |
| 25 mL virgin OO (HTyr 8.5 mg/kg, HTyr acetate 33.4 mg/kg, Olea 269.3 mg/kg, Oleu aglycone 28.5 mg/kg, Tyr 4.4 mg/kg, Oleo 11.4 mg/kg, Lig aglycone 9.9 mg/kg) [52] | (Not analyzed) | (Not analyzed) | HTyr, HTyr acetate, Tyr, phenylacetic acid, 2-(4′- hydroxyphenyl)acetic acid, 2-(3′-hydroxyphenyl)acetic acid, 3-(4′-hydroxyphenyl)-propionic acid | A moderate intake of a phenol-rich OO raised the concentration in human feces of free HTyr and phenylacetic and phenylpropionic acids. The products of colonic catabolism of OO phenolic compounds could be good candidates for novel preventive strategies and in colon and other bowel diseases. |
| Olive leaf extract (250 mg of olive leaf extract rich in oleuropein (40%) [131] | Luteolin, luteolin-glu, HTyr-sulf-glu, HTyr-glu, HTyr-sulf, HVAlc-glu, Oleu aglycon-glu, Oleu derivative, Oleu-sulf, Tyr-glu, HT acetate glu | HTyr-sulf-glu, HTyr-glu, HTyr-sulf, HVAlc-glu, HVAlc-sulf, Oleu aglycon-glu, Oleu derivative, Oleu-sulf, Tyr-glu, EA, EA-glu, HTyr acetate-glu | (Not analyzed) | Plasma levels of HTyr glu, HTyr-sulf, Oleu aglycon-glu and Oleu aglycon derivative were higher in post-menopausal women. Post-menopausal women excreted less sulfated metabolites in urine than pre-menopausal women. |
| HTyr 5 or 25 mg Htyr/day HT group supplemented as capsule [132] | (Not analyzed) | Htyr, Htyr-4-glucuronide, Htyr-3-glucuronide, Htyr-3-sulphate, Htyr-4-sulphate | (Not analyzed) | The results show that Htyr given as the foremost component of a nutraceutical preparation is bioavailable and is recovered in the urine chiefly as sulphate-3′, which can be adopted as biomarker of extra virgin OO consumption. |
| Ethanol (12, 18, 24, 30 and 42 g) or placebo [133] | (Not analyzed) | Htyr, Tyr, DOPAC and HVA | (Not analyzed) | Urinary excretion of Htyr and Tyr increased with ethanol administered dose. This study demonstrates an endogenous production of Htyr and Tyr in relation to ethanol administered dose in humans. |
| OO phenols (100 mg) as supplements [134] | (Not analyzed) | Htyr, Tyr | (Not analyzed) | Humans absorb a large part of ingested OO phenols and absorbed olive oil phenols are extensively modified in the body. |
| Olive leaf supplement as capsule or liquid (20 mg Oleu/tablet and 22 mg Oleu/5 mL) [129] | (Not analyzed) | Glucuronic acid conjugates, derived from oleuropein aglycone, uric acid, hippuric acid, 3- and 4-hydroxyhippuric acid | (Not analyzed) | The presence of the metabolites in urine indicates that Oleu reaches systemic circulation and is metabolized in the human system. |
| Spiked LDL samples with Htyr (0.15 μg/mL) and HVA (1 μg/mL) [135] | Htyr-glu, Htyr-sulf, Tyr-glu, Tyr-sulf, HVA-sulf | (Not analyzed) | (Not analyzed) | A rapid method for the detection and quantification of metabolites of OO phenolic compounds in LDL by SPE and HPLC/ESI-MS/was developed. |
| 50 mL of extra virgin OO (6 mg/20 g Htyr) [96] | EA + H2, Olea, Lig aglycone, Lig aglycone + H2 + glu, Oleu aglycon + OH, Oleo + H2 + glu, apigenin + CH3 + glu | Oleu aglycone, Oleu aglycone + H2 + glu, methyl-lig aglycone + H2 + gluc, lig aglycone +H2, Oleu aglycone + OH + sulf, luteolin + CH3+ glu, Oleo + glu, Oleu aglycone + OH + glu | (Not analyzed) | Maximum absorption levels of the plasma biomarkers were detected between 0.5 and 2 h, and maximum excretion of urinary biomarkers occurred in the first 4 h after extra virgin OO intake. Plasma secoiridoid compounds were selected as biomarkers to monitor extra virgin OO intake showing good predictive ability according to multivariate analysis. |
| Virgin OO (80 mg total phenols/kg oil), functional virgin OO enriched with its own phenolics (500 mg total phenols/kg oil), functional virgin OO enriched with its own phenolics plus complementary phenolics from Thyme (500 mg total phenols/kg oil, 50% from olive oil and 50% from thyme respectively) [136] | HTyr-sulf, thymol-sulf, HTyr acetate-sulf, HVAlc-sulf, HVA-sulf, hydroxyphenylpropionic acid-sulf, caffeic acid-sulf | HTyr-sulf, HVAlc-sulf, HTyr acetate-sulf, HVA-sulf, HTyr-glu, HVAlc-glu, hydroxyphenylpropionic acid-sulf, caffeic acid-sulf, thymol-sulf, thymol- glu, p-Cymene-diol glu | (Not analyzed) | HTyr-sulf and HTyr acetate-sulf appeared to be suitable biomarkers for monitoring compliance with OO intake as their values in plasma or/and 24 h urine were significantly higher after functional virgin OO compared to baseline pre-intervention concentrations. |
| 25 mL virgin OO (2.7 mg TPC/kg OO or 366 mg TPC/kg OO) [137] | HTyr-sulf, HVA-sulf, HVA-glu | (Not analyzed) | (Not analyzed) | The levels of the studied metabolites increased significantly after high TPC virgin OO ingestion compared to low TPC OO. Virgin OO consumption increases the levels of phenolic metabolites in HDL and thus provides human HDL with more efficient antioxidant protection. |
| 50 mL extra virgin OO (HTyr 8.31 mg/kg, Tyr 5.33 mg/kg, pinoresinol 3.25 m/kg, luteolin 2.65 mg/kg, apigenin 0.64 mg/kg, EA 34.91 mg/kg, lig aglycone 40.58 mg/kg) [138] | (Not analyzed) | 50 metabolites, obtained through phase I and phase II metabolic reactions were tentatively identified. | (Not analyzed) | Ten of these biomarkers and more than 50 metabolites obtained through phase I and phase II biotransformation reactions were tentatively identified. Additionally, kinetic studies were conducted on the metabolites identified as possible biomarkers; for most of the compounds, concentrations were maximum in the first two hours. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikou, T.; Sakavitsi, M.E.; Kalampokis, E.; Halabalaki, M. Metabolism and Bioavailability of Olive Bioactive Constituents Based on In Vitro, In Vivo and Human Studies. Nutrients 2022, 14, 3773. https://doi.org/10.3390/nu14183773
Nikou T, Sakavitsi ME, Kalampokis E, Halabalaki M. Metabolism and Bioavailability of Olive Bioactive Constituents Based on In Vitro, In Vivo and Human Studies. Nutrients. 2022; 14(18):3773. https://doi.org/10.3390/nu14183773
Chicago/Turabian StyleNikou, Theodora, Maria Eleni Sakavitsi, Evangelos Kalampokis, and Maria Halabalaki. 2022. "Metabolism and Bioavailability of Olive Bioactive Constituents Based on In Vitro, In Vivo and Human Studies" Nutrients 14, no. 18: 3773. https://doi.org/10.3390/nu14183773
APA StyleNikou, T., Sakavitsi, M. E., Kalampokis, E., & Halabalaki, M. (2022). Metabolism and Bioavailability of Olive Bioactive Constituents Based on In Vitro, In Vivo and Human Studies. Nutrients, 14(18), 3773. https://doi.org/10.3390/nu14183773