
Vitamin K-Dependent Carboxylation of Osteocalcin in Bone—Ally or Adversary of Bone Mineral Status in Rats with Experimental Chronic Kidney Disease?


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Measurement of Vitamin K Concentrations in Rat Serum
2.3. Serum and Bone Levels of Vitamin K-Dependent Proteins
2.4. Preparation of Bone Tissue Homogenates
2.5. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR)
2.6. Genes of Osteoblastogenesis
2.7. The Mineral Status of Femurs
2.8. Statistical Analysis
3. Results
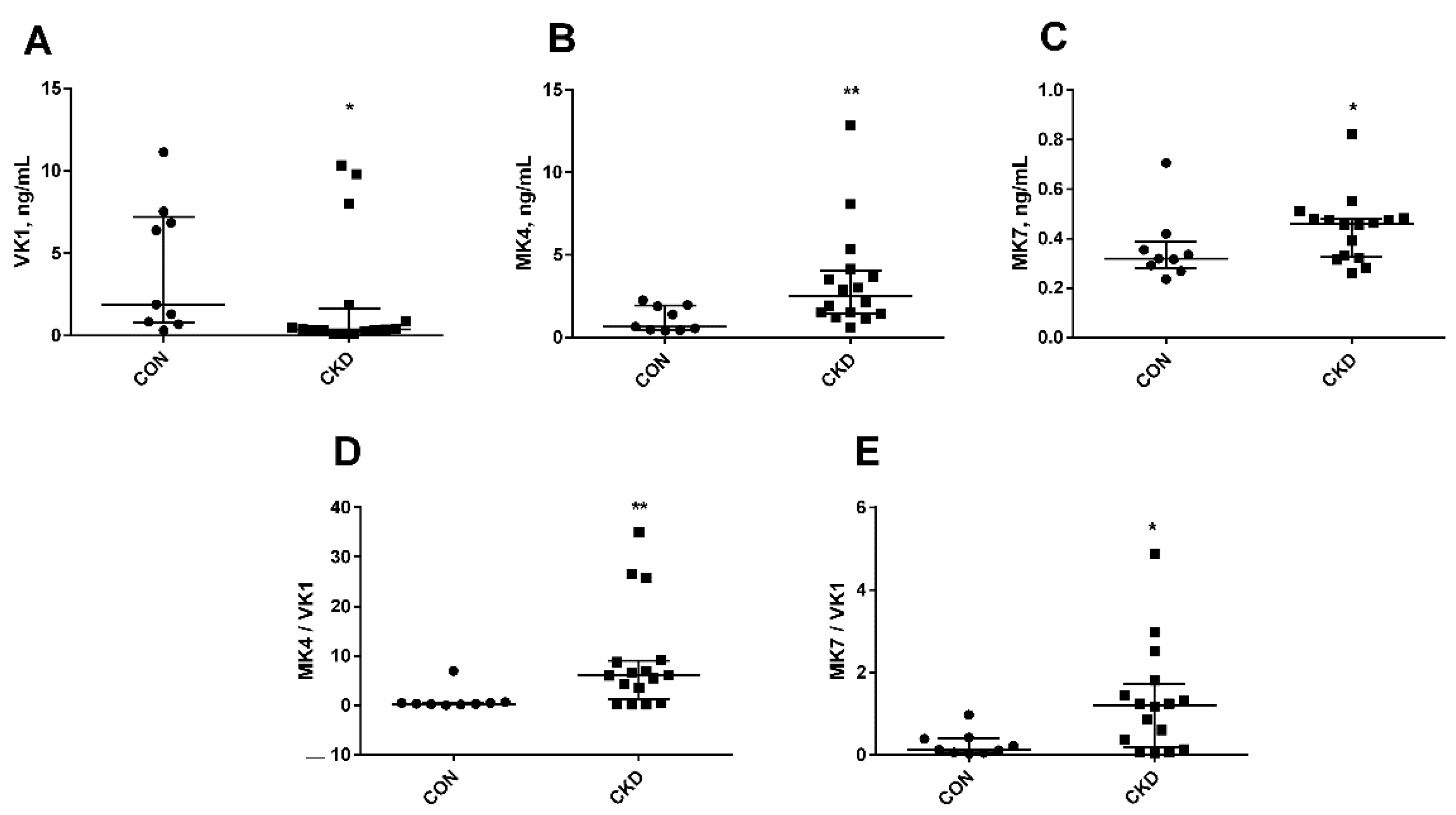
3.1. The Status of Endogenous Vitamin K in Rats with CKD
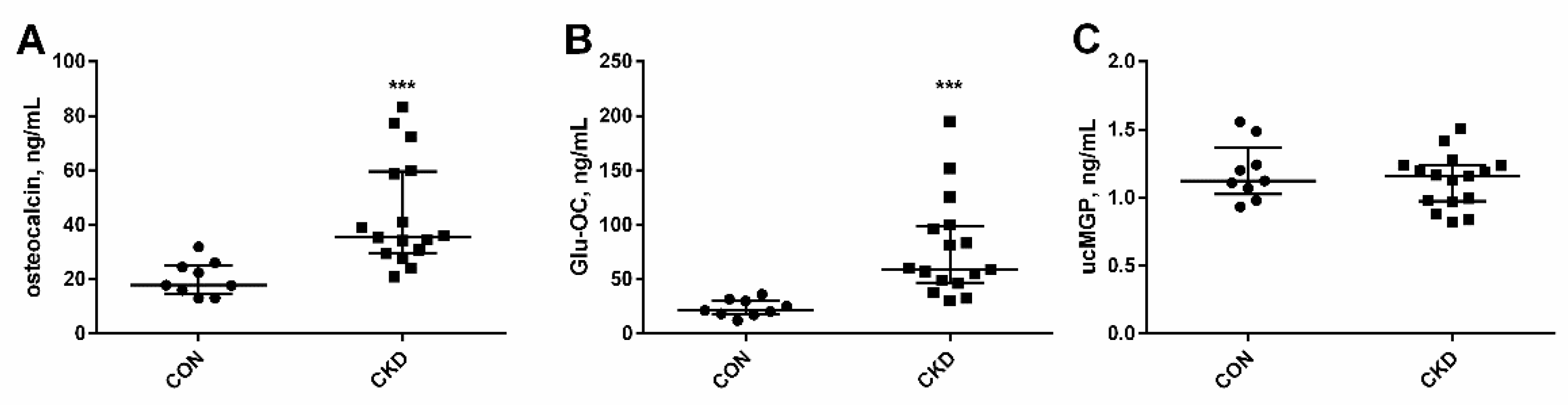
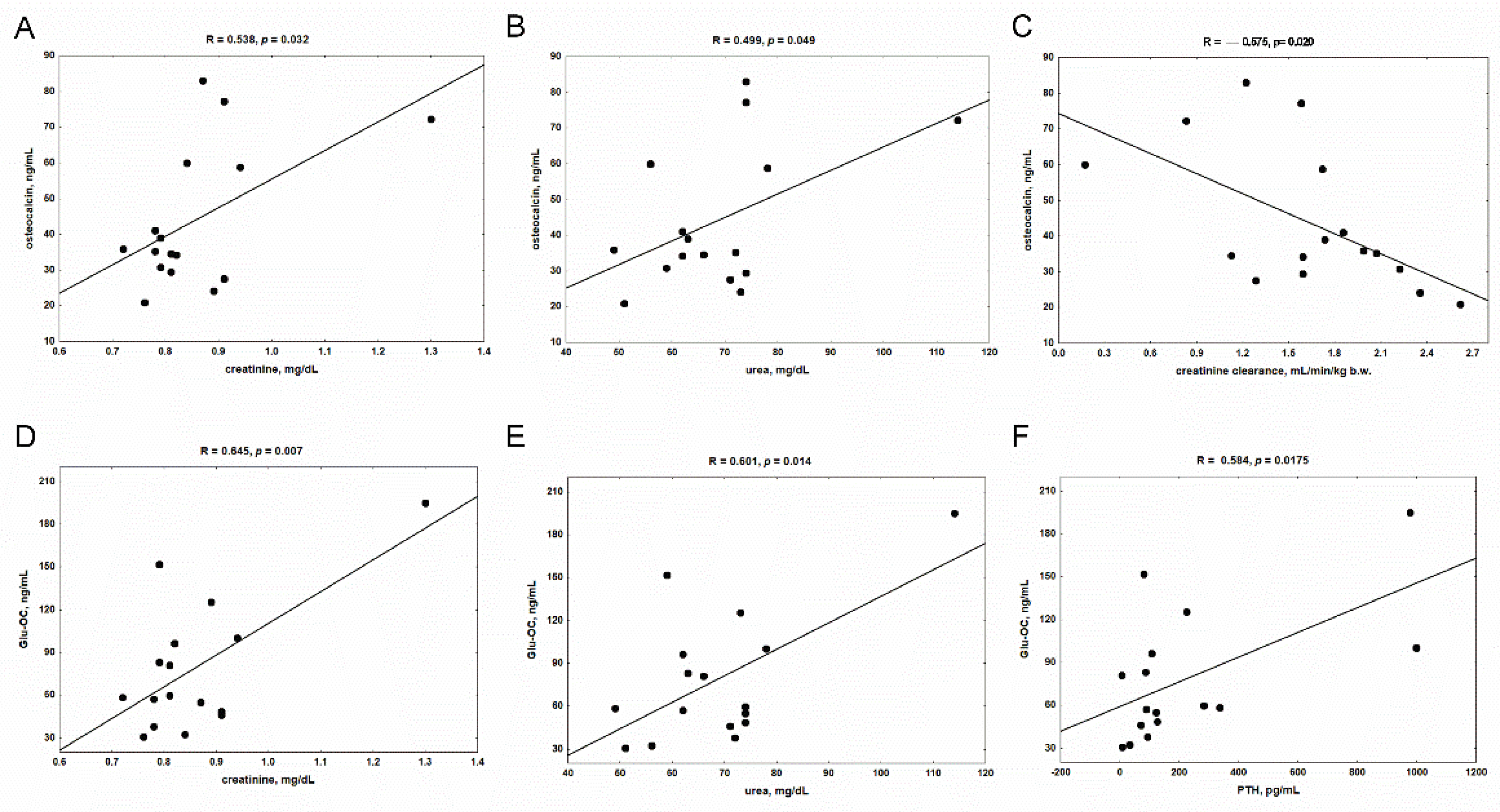
3.2. Serum Levels of Vitamin K-Dependent Proteins in Rats with CKD, and the Impact of Kidney Function and PTH on Their Concentrations
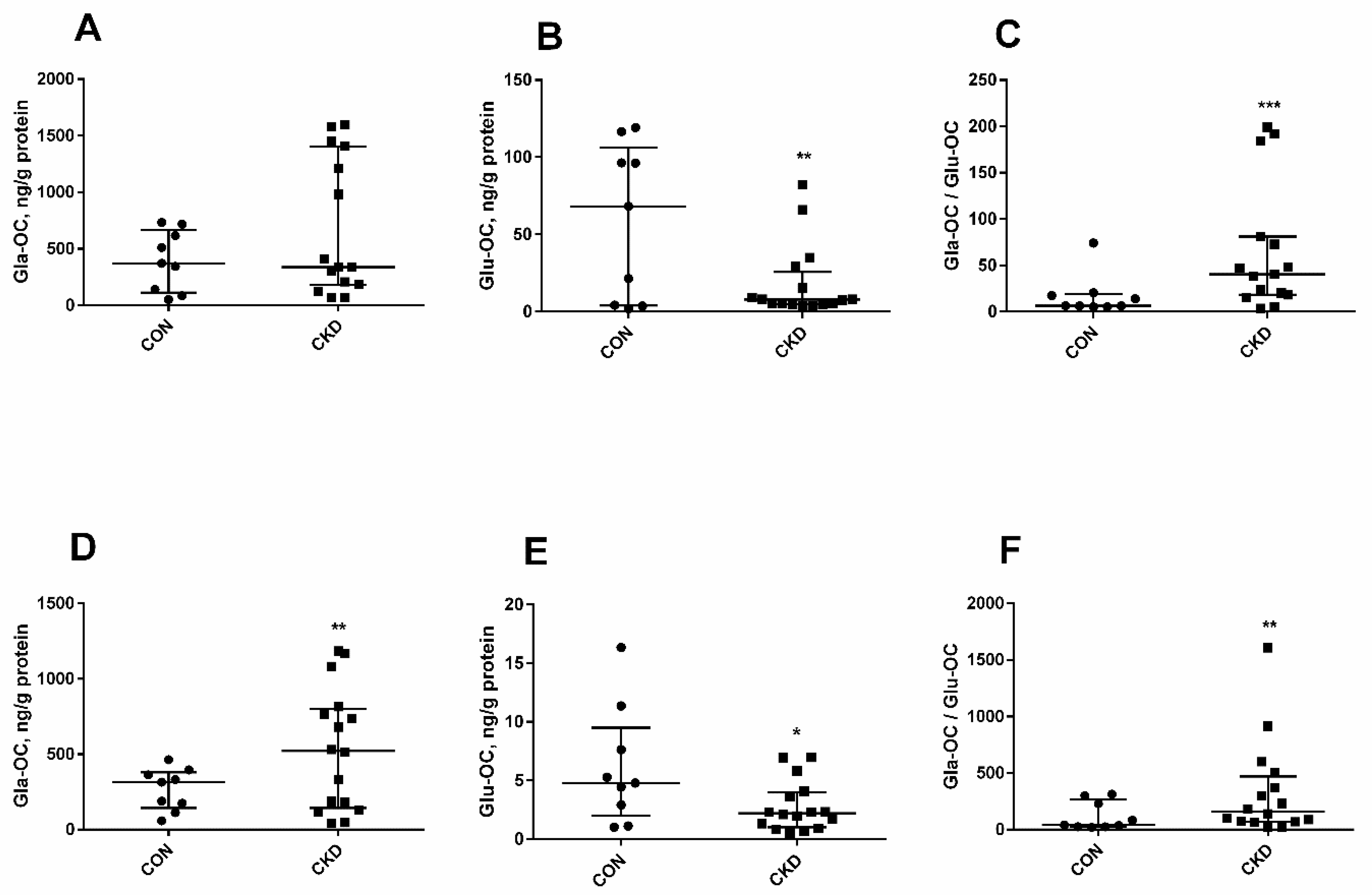
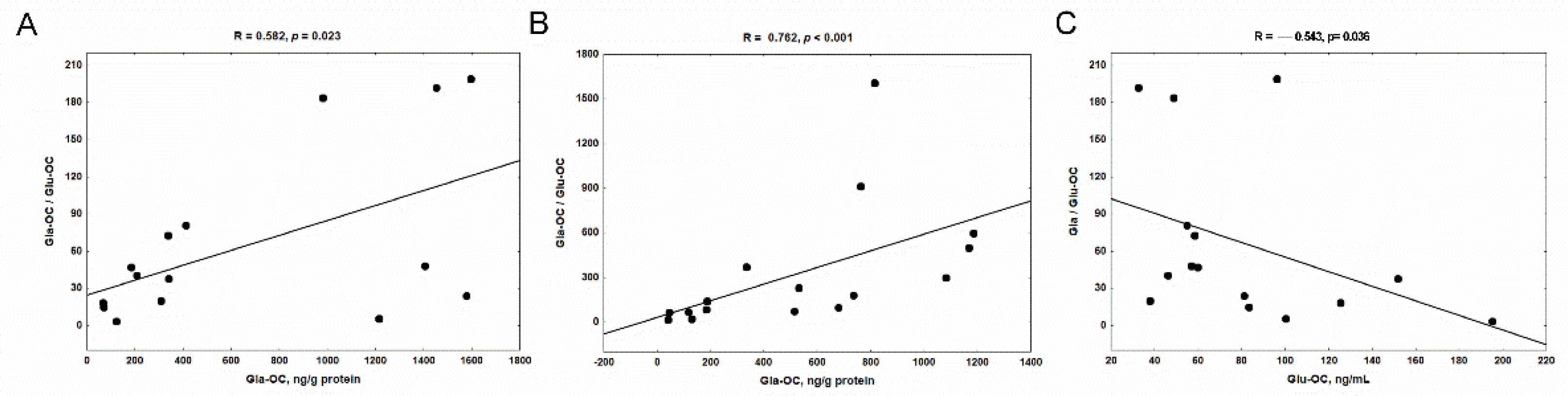
3.3. The Levels of Glu-OC, Gla-OC, and Gla-OC/Glu-OC Ratios in Femoral Bone Tissue of Rats with CKD, and Their Relations with Serum Glu-OC
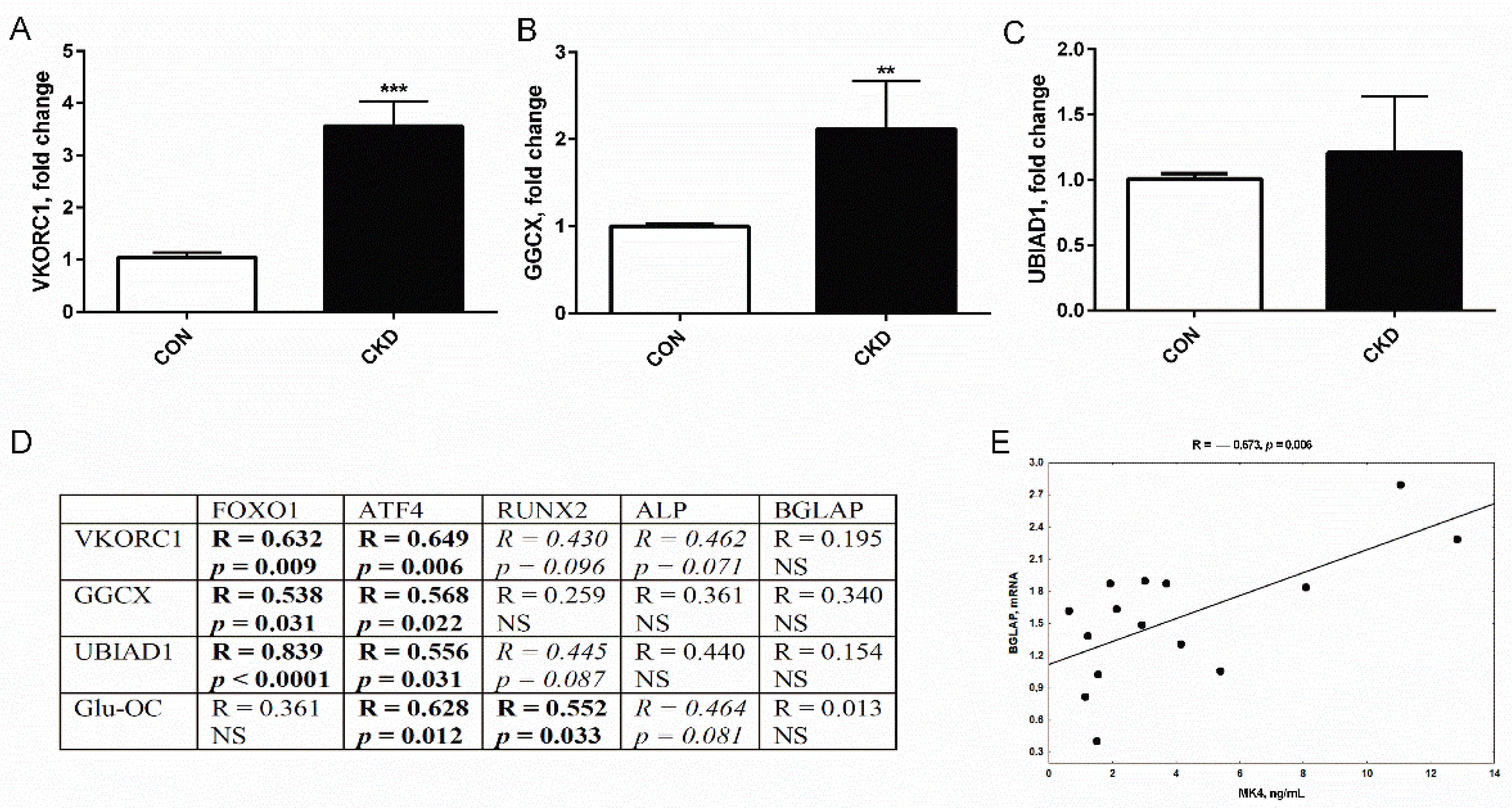
3.4. The Expression of Genes Coding Vitamin K Cycle Enzymes in Femurs of Rats with CKD, and Their Associations with Genes Participating in Osteoblastogenesis
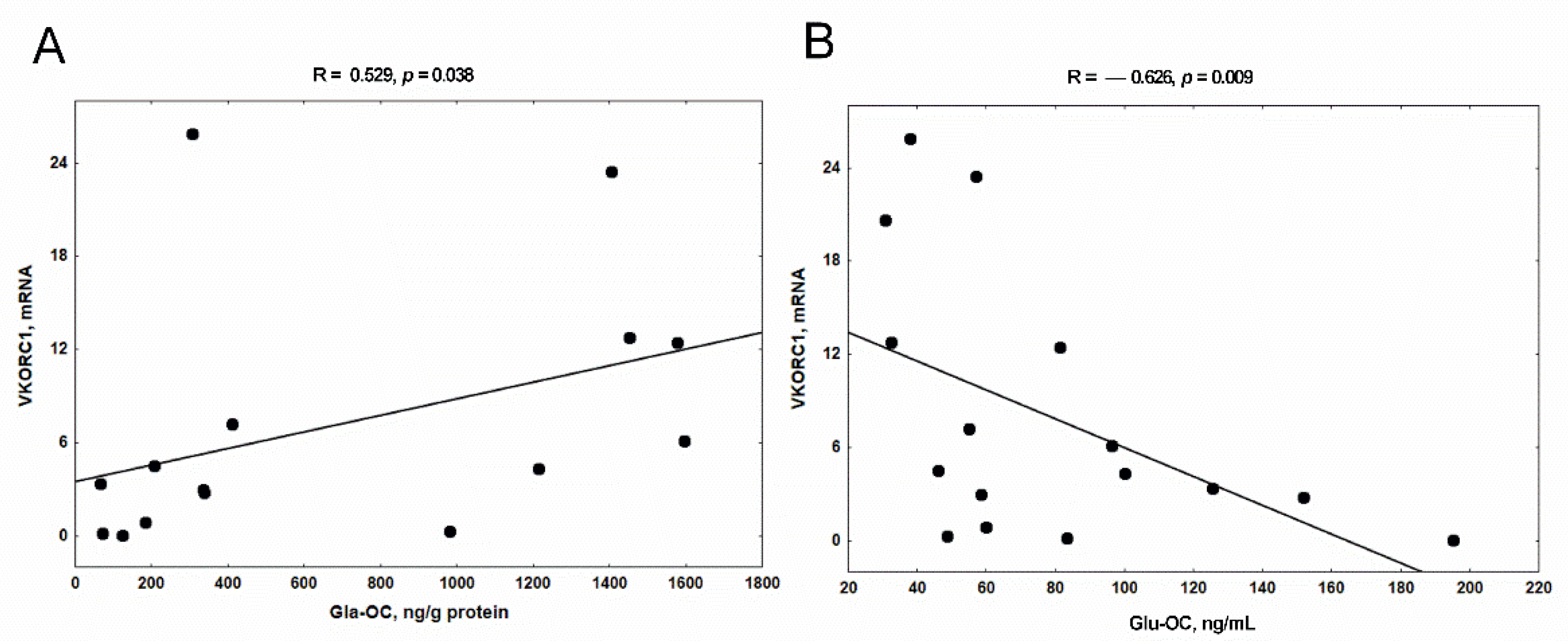
3.5. The Relationship between the Expression of VKORC1 and the Concentration of Gla-OC in Trabecular Bone Tissue and Serum Glu-OC
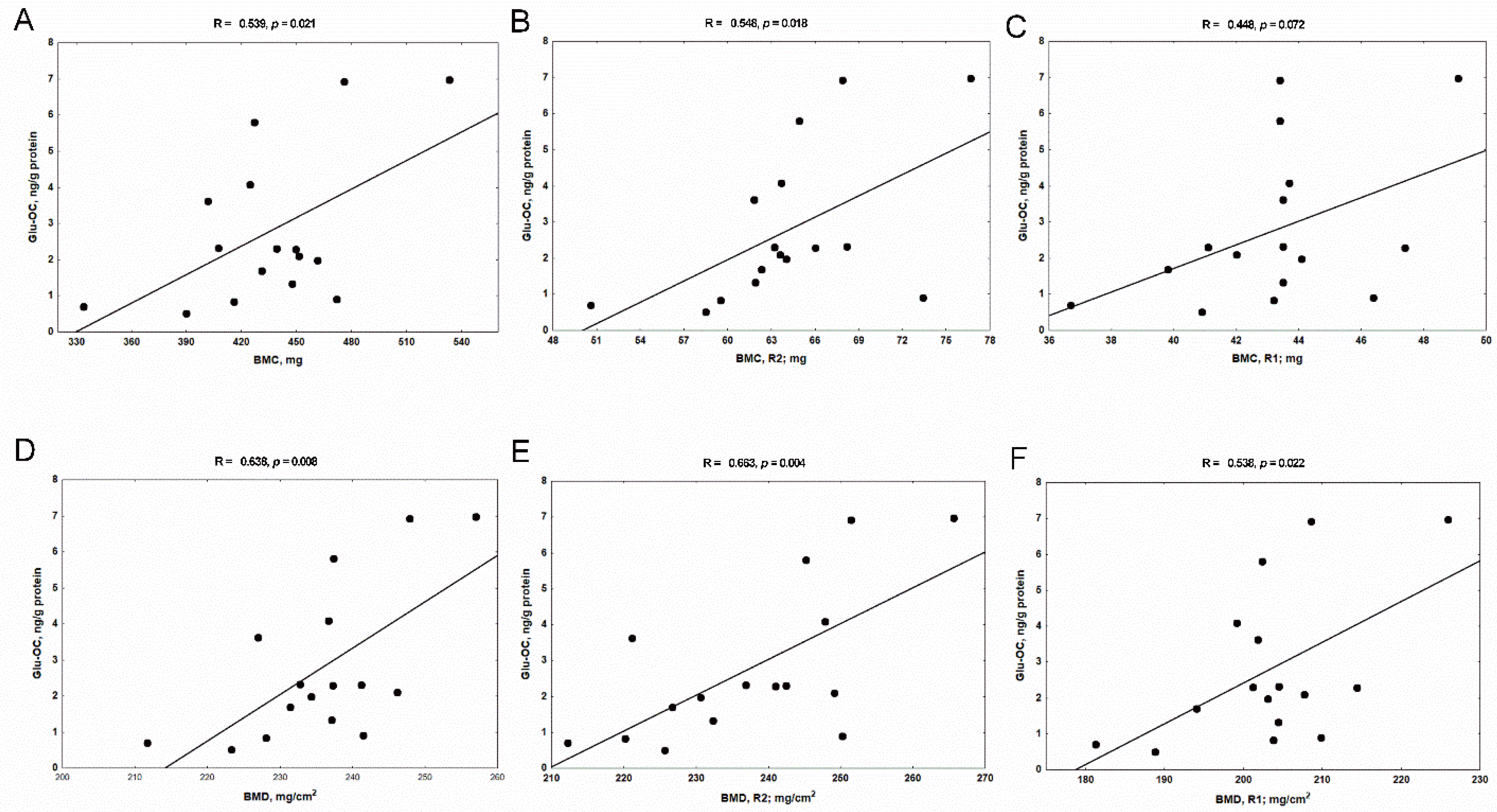
3.6. The Associations between Bone Glu-OC, Gla-OC, and Bone Mineral Status in Rats with CKD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1,25 (OH)2D | 1,2-dihydroxyvitamin D |
| ALP | alkaline phosphatase |
| ATF4 | activating transcription factor 4 |
| BGLAP | bone gamma carboxyglutamate protein; osteocalcin |
| BMC | bone mineral content |
| BMD | bone mineral density |
| CKD | chronic kidney disease |
| CKD-MBD | chronic kidney disease–mineral bone disorders |
| CON | control group |
| dp-ucMGP | desphospho-uncarboxylated matrix Gla protein |
| FOXO1 | forkhead box transcription factor 1 |
| GGCX | γ-glutamyl carboxylase |
| Gla | gamma carboxyglutamic acid |
| Gla-OC | carboxylated osteocalcin |
| Glu | glutamic acid |
| Glu-OC | undercarboxylated osteocalcin |
| HD | hemodialysis |
| MGP | matrix gla protein |
| MKs | menaquinones |
| MK4 | menaquinone 4 |
| MK7 | menaquinone 7 |
| OC | osteocalcin |
| PTH | parathyroid hormone |
| R1 | metaphyseal area of femur |
| R2 | diaphyseal area of femur |
| RUNX2 | Runt-related transcription factor 2 |
| UBIAD1 | UbiA prenyltransferase domain-containing 1 protein |
| ucMGP | uncarboxylated matrix gla protein |
| ucOC | uncarboxylated osteocalcin |
| VK | vitamin K |
| VK1 | vitamin K1; phylloquinone |
| VK2 | vitamin K2; menaquinones |
| VKDPs | vitamin K-dependent proteins |
| VKORC1 | vitamin K epoxide reductase complex subunit 1 |
References
- Waziri, B.; Duarte, R.; Naicker, S. Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD): Current perspectives. Int. J. Nephrol. Renovasc. Dis. 2019, 12, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R. Mineral and bone disorders in chronic kidney disease: New insights into mechanism and management. Ann. Clin. Biochem. 2012, 49, 432–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidney Disease: Improving Global Outcomes (KDIGO) CKD-MBD Update Work Group. KDIGO 2017 Clinical Practice Guideline Update for the Diagnosis, Evaluation, Prevention, and Treatment of Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD). Kidney Int. Suppl. 2017, 7, 1–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimentel, A.; Ureña-Torres, P.; Zillikens, M.C.; Bover, J.; Cohen-Solal, M. Fractures in patients with CKD-diagnosis, treatment, and prevention: A review by members of the European Calcified Tissue Society and the European Renal Association of Nephrology Dialysis and Transplantation. Kidney Int. 2017, 92, 1343–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, P.D. Chronic kidney disease and osteoporosis: Evaluation and management. Bonekey Rep. 2014, 3, 542. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, M. Skeletal remodeling in health and disease. Nat. Med. 2007, 13, 791–801. [Google Scholar] [CrossRef]
- Katsimbri, P. The biology of normal bone remodelling. Eur. J. Cancer Care (Engl.) 2017, 26. [Google Scholar] [CrossRef]
- Akbari, S.; Rasouli-Ghahroudi, A.A. Vitamin K and Bone Metabolism: A Review of the Latest Evidence in Preclinical Studies. Biomed Res. Int. 2018, 2018, 4629383. [Google Scholar] [CrossRef] [Green Version]
- Napoli, N.; Strollo, R.; Sprini, D.; Maddaloni, E.; Rini, G.B.; Carmina, E. Serum 25-OH vitamin D in relation to bone mineral density and bone turnover. Int. J. Endocrinol. 2014, 2014, 487463. [Google Scholar] [CrossRef]
- Booth, S.L. Vitamin K: Food composition and dietary intakes. Food Nutr. Res. 2012, 56, 5505. [Google Scholar] [CrossRef] [Green Version]
- Thijssen, H.H.; Drittij-Reijnders, M.J. Vitamin K distribution in rat tissues: Dietary phylloquinone is a source of tissue menaquinone-4. Br. J. Nutr. 1994, 72, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature. 2010, 468, 117–121. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Vermeer, C. Determination of phylloquinone and menaquinones in food. Effect of food matrix on circulating vitamin K concentrations. Haemostasis. 2000, 30, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Tie, J.K.; Stafford, D.W. Structural and functional insights into enzymes of the vitamin K cycle. J. Thromb. Haemost. 2016, 14, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silaghi, C.N.; Ilyés, T.; Filip, V.P.; Farcaș, M.; van Ballegooijen, A.J.; Crăciun, A.M. Vitamin K Dependent Proteins in Kidney Disease. Int. J. Mol. Sci. 2019, 20, 1571. [Google Scholar] [CrossRef] [Green Version]
- Fusaro, M.; Gallieni, M.; Rizzo, M.A.; Stucchi, A.; Delanaye, P.; Cavalier, E.; Moysés, R.; Jorgetti, V.; Iervasi, G.; Giannini, S.; et al. Vitamin K plasma levels determination in human health. Clin. Chem. Lab. Med. 2017, 55, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Caluwé, R.; Verbeke, F.; De Vriese, A.S. Evaluation of vitamin K status and rationale for vitamin K supplementation in dialysis patients. Nephrol. Dial. Transplant. 2020, 35, 23–33. [Google Scholar] [CrossRef]
- Cranenburg, E.C.; Schurgers, L.J.; Uiterwijk, H.H.; Beulens, J.W.; Dalmeijer, G.W.; Westerhuis, R.; Magdeleyns, E.J.; Herfs, M.; Vermeer, C.; Laverman, G.D. Vitamin K intake and status are low in hemodialysis patients. Kidney Int. 2012, 82, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Holden, R.M.; Morton, A.R.; Garland, J.S.; Pavlov, A.; Day, A.G.; Booth, S.L. Vitamins K and D status in stages 3-5 chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2010, 5, 590–597. [Google Scholar] [CrossRef] [Green Version]
- McCabe, K.M.; Adams, M.A.; Holden, R.M. Vitamin K status in chronic kidney disease. Nutrients 2013, 5, 4390–4398. [Google Scholar] [CrossRef]
- Voong, K.; Harrington, D.; Goldsmith, D. Vitamin K status in chronic kidney disease: A report of a study and a mini-review. Int. Urol. Nephrol. 2013, 45, 1339–1344. [Google Scholar] [CrossRef] [PubMed]
- Stankowiak-Kulpa, H.; Krzyżanowska, P.; Kozioł, L.; Grzymisławski, M.; Wanic-Kossowska, M.; Moczko, J.; Walkowiak, J. Vitamin K status in peritoneally dialyzed patients with chronic kidney disease. Acta Biochim. Pol. 2011, 58, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Westenfeld, R.; Krueger, T.; Schlieper, G.; Cranenburg, E.C.; Magdeleyns, E.J.; Heidenreich, S.; Holzmann, S.; Vermeer, C.; Jahnen-Dechent, W.; Ketteler, M.; et al. Effect of vitamin K2 supplementation on functional vitamin K deficiency in hemodialysis patients: A randomized trial. Am. J. Kidney Dis. 2012, 59, 186–195. [Google Scholar] [CrossRef] [PubMed]
- McCabe, K.M.; Booth, S.L.; Fu, X.; Ward, E.; Adams, M.A.; Holden, R.M. Vitamin K Metabolism in a Rat Model of Chronic Kidney Disease. Am. J. Nephrol. 2017, 45, 4–13. [Google Scholar] [CrossRef]
- Pilkey, R.M.; Morton, A.R.; Boffa, M.B.; Noordhof, C.; Day, A.G.; Su, Y.; Miller, L.M.; Koschinsky, M.L.; Booth, S.L. Subclinical Vitamin K Deficiency in Hemodialysis Patients. Am. J. Kidney Dis. 2007, 49, 432–439. [Google Scholar] [CrossRef]
- Kohlmeier, M.; Saupe, J.; Shearer, M.J.; Schaefer, K.; Asmus, G. Bone health of adult hemodialysis patients is related to vitamin K status. Kidney Int. 1997, 51, 1218–1221. [Google Scholar] [CrossRef] [Green Version]
- Evenepoel, P.; Claes, K.; Meijers, B.; Laurent, M.; Bammens, B.; Naesens, M.; Sprangers, B.; Pottel, H.; Cavalier, E.; Kuypers, D. Poor Vitamin K Status Is Associated With Low Bone Mineral Density and Increased Fracture Risk in End-Stage Renal Disease. J. Bone Miner. Res. 2019, 34, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Kim, K.M.; Kim, B.T.; Joo, N.S.; Kim, K.N.; Lee, D.J. Correlation of undercarboxylated osteocalcin (ucOC) concentration and bone density with age in healthy Korean women. J. Korean Med. Sci. 2010, 25, 1171–1175. [Google Scholar] [CrossRef] [Green Version]
- Bügel, S. Vitamin K and bone health in adult humans. Vitam. Horm. 2008, 78, 393–416. [Google Scholar] [CrossRef]
- Rodríguez-Olleros Rodríguez, C.; Díaz Curiel, M. Vitamin K and Bone Health: A Review on the Effects of Vitamin K Deficiency and Supplementation and the Effect of Non-Vitamin K Antagonist Oral Anticoagulants on Different Bone Parameters. J. Osteoporos. 2019, 2019, 2069176. [Google Scholar] [CrossRef]
- Braam, L.A.; Knapen, M.H.; Geusens, P.; Brouns, F.; Hamulyák, K.; Gerichhausen, M.J.; Vermeer, C. Vitamin K1 supplementation retards bone loss in postmenopausal women between 50 and 60 years of age. Calcif. Tissue Int. 2003, 73, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Knapen, M.H.; Drummen, N.E.; Smit, E.; Vermeer, C.; Theuwissen, E. Three-year low-dose menaquinone-7 supplementation helps decrease bone loss in healthy postmenopausal women. Osteoporos. Int. 2013, 24, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- Shiraki, M.; Shiraki, Y.; Aoki, C.; Miura, M. Vitamin K2 (menatetrenone) effectively prevents fractures and sustains lumbar bone mineral density in osteoporosis. J. Bone Miner. Res. 2000, 15, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Cockayne, S.; Adamson, J.; Lanham-New, S.; Shearer, M.J.; Gilbody, S.; Torgerson, D.J. Vitamin K and the prevention of fractures: Systematic review and meta-analysis of randomized controlled trials. Arch. Intern. Med. 2006, 166, 1256–1261. [Google Scholar] [CrossRef]
- Iwamoto, J.; Takeda, T.; Ichimura, S. Effect of combined administration of vitamin D3 and vitamin K2 on bone mineral density of the lumbar spine in postmenopausal women with osteoporosis. J. Orthop. Sci. 2000, 5, 546–551. [Google Scholar] [CrossRef]
- Huang, Z.B.; Wan, S.L.; Lu, Y.J.; Ning, L.; Liu, C.; Fan, S.W. Does vitamin K2 play a role in the prevention and treatment of osteoporosis for postmenopausal women: A meta-analysis of randomized controlled trials. Osteoporos. Int. 2015, 26, 1175–1186. [Google Scholar] [CrossRef]
- Knapen, M.H.; Schurgers, L.J.; Vermeer, C. Vitamin K2 supplementation improves hip bone geometry and bone strength indices in postmenopausal women. Osteoporos. Int. 2007, 18, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Emaus, N.; Gjesdal, C.G.; Almås, B.; Christensen, M.; Grimsgaard, A.S.; Berntsen, G.K.; Salomonsen, L.; Fønnebø, V. Vitamin K2 supplementation does not influence bone loss in early menopausal women: A randomised double-blind placebo-controlled trial. Osteoporos. Int. 2010, 21, 1731–1740. [Google Scholar] [CrossRef]
- Binkley, N.; Harke, J.; Krueger, D.; Engelke, J.; Vallarta-Ast, N.; Gemar, D.; Checovich, M.; Chappell, R.; Suttie, J. Vitamin K treatment reduces undercarboxylated osteocalcin but does not alter bone turnover, density, or geometry in healthy postmenopausal North American women. J. Bone Miner. Res. 2009, 24, 983–991. [Google Scholar] [CrossRef]
- Booth, S.L.; Pennington, J.A.; Sadowski, J.A. Food sources and dietary intakes of vitamin K-1 (phylloquinone) in the American diet: Data from the FDA Total Diet Study. J. Am. Diet. Assoc. 1996, 96, 149–154. [Google Scholar] [CrossRef]
- Caluwé, R.; Vandecasteele, S.; Van Vlem, B.; Vermeer, C.; De Vriese, A.S. Vitamin K2 supplementation in haemodialysis patients: A randomized dose-finding study. Nephrol. Dial. Transplant. 2014, 29, 1385–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoun, M.; Makki, M.; Azar, H.; Matta, H.; Chelala, D.N. High Dephosphorylated-Uncarboxylated MGP in Hemodialysis patients: Risk factors and response to vitamin K2, A pre-post intervention clinical trial. BMC Nephrol. 2017, 18, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, N.; Kusano, E.; Takahashi, H.; Ando, Y.; Yano, K.; Tsuda, E.; Asano, Y. Vitamin K2 inhibits glucocorticoid-induced bone loss partly by preventing the reduction of osteoprotegerin (OPG). J. Bone Miner. Metab. 2005, 23, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, D.; Znorko, B.; Kalaska, B.; Domaniewski, T.; Zawadzki, R.; Lipowicz, P.; Doroszko, M.; Łebkowska, U.; Grabowski, P.; Pawlak, K. LP533401 restores bone health in 5/6 nephrectomized rats by a decrease of gut-derived serotonin and regulation of serum phosphate through the inhibition of phosphate co-transporters expression in the kidneys. Bone 2018, 113, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Znorko, B.; Pawlak, D.; Oksztulska-Kolanek, E.; Domaniewski, T.; Pryczynicz, A.; Roszczenko, A.; Rogalska, J.; Lipowicz, P.; Doroszko, M.; Brzoska, M.M.; et al. RANKL/OPG system regulation by endogenous PTH and PTH1R/ATF4 axis in bone: Implications for bone accrual and strength in growing rats with mild uremia. Cytokine 2018, 106, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, D.; Domaniewski, T.; Sieklucka, B.; Jakuc, M.; Pawlak, K. Inhibition of peripheral serotonin synthesis by LP533401 and disturbances in calciotropic hormones attenuated excessive osteoblastogenesis with simultaneous improvement of bone mineral status in 5/6 nephrectomized rats. Biochim. Biophys. Acta Mol. Basis. Dis. 2019, 1865, 165528. [Google Scholar] [CrossRef]
- de Oliveira, R.B.; Stinghen, A.E.M.; Massy, Z.A. Vitamin K role in mineral and bone disorder of chronic kidney disease. Clin. Chim. Acta. 2020, 502, 66–72. [Google Scholar] [CrossRef]
- Bellone, F.; Cinquegrani, M.; Nicotera, R.; Carullo, N.; Casarella, A.; Presta, P.; Andreucci, M.; Squadrito, G.; Mandraffino, G.; Prunestì, M.; et al. Role of Vitamin K in Chronic Kidney Disease: A Focus on Bone and Cardiovascular Health. Int. J. Mol. Sci. 2022, 23, 5282. [Google Scholar] [CrossRef]
- Shearer, M.J.; Bach, A.; Kohlmeier, M. Chemistry, nutritional sources, tissue distribution and metabolism of vitamin K with special reference to bone health. J. Nutr. 1996, 126, 1181S–1186S. [Google Scholar] [CrossRef] [Green Version]
- Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of phylloquinone (Vitamin K1) into menaquinone-4 (Vitamin K2) in mice: Two possible routes for menaquinone-4 accumulation in cerebra of mice. J. Biol. Chem. 2008, 283, 11270–11279. [Google Scholar] [CrossRef] [Green Version]
- Thijssen, H.H.; Drittij-Reijnders, M.J.; Fischer, M.A. Phylloquinone and menaquinone-4 distribution in rats: Synthesis rather than uptake determines menaquinone-4 organ concentrations. J. Nutr. 1996, 126, 537–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, Y.; Tsugawa, N.; Nakagawa, K.; Suhara, Y.; Tanaka, K.; Uchino, Y.; Takeuchi, A.; Sawada, N.; Kamao, M.; Wada, A.; et al. Menadione (vitamin K3) is a catabolic product of oral phylloquinone (vitamin K1) in the intestine and a circulating precursor of tissue menaquinone-4 (vitamin K2) in rats. J. Biol. Chem. 2013, 288, 33071–33080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, S.; Hanzawa, F.; Takahashi, S.; Suzuki, N.; Sano, K.; Oda, H.; Uchida, T. Tissue Distribution of Menaquinone-7 and the Effect of α-Tocopherol Intake on Menaquinone-7 Concentration in Rats. J. Nutr. Sci. Vitaminol. (Tokyo) 2018, 64, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Booth, S.L.; Smith, D.E. Vitamin K contents of rodent diets: A review. J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 8–12. [Google Scholar]
- Fusaro, M.; Cozzolino, M.; Plebani, M.; Iervasi, G.; Ketteler, M.; Gallieni, M.; Aghi, A.; Locatelli, F.; Cunningham, J.; Salam, S.; et al. Sevelamer Use, Vitamin K Levels, Vascular Calcifications, and Vertebral Fractures in Hemodialysis Patients: Results from the VIKI Study. J. Bone Miner. Res. 2021, 36, 500–509. [Google Scholar] [CrossRef] [PubMed]
- McCabe, K.M.; Booth, S.L.; Fu, X.; Shobeiri, N.; Pang, J.J.; Adams, M.A.; Holden, R.M. Dietary vitamin K and therapeutic warfarin alter the susceptibility to vascular calcification in experimental chronic kidney disease. Kidney Int. 2013, 83, 835–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferland, G.; Doucet, I.; Mainville, D. Phylloquinone and Menaquinone-4 Tissue Distribution at Different Life Stages in Male and Female Sprague-Dawley Rats Fed Different VK Levels Since Weaning or Subjected to a 40% Calorie Restriction since Adulthood. Nutrients 2016, 8, 141. [Google Scholar] [CrossRef]
- Price, P.A.; Williamson, M.K.; Lothringer, J.W. Origin of the vitamin K-dependent bone protein found in plasma and its clearance by kidney and bone. J. Biol. Chem. 1981, 256, 12760–12766. [Google Scholar] [CrossRef]
- Shimizu, T.; Takahata, M.; Kameda, Y.; Hamano, H.; Ito, T.; Kimura-Suda, H.; Todoh, M.; Tadano, S.; Iwasaki, N. Vitamin K-dependent carboxylation of osteocalcin affects the efficacy of teriparatide (PTH(1-34)) for skeletal repair. Bone 2014, 64, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Nagura, N.; Komatsu, J.; Iwase, H.; Hosoda, H.; Ohbayashi, O.; Nagaoka, I.; Kaneko, K. Effects of the combination of vitamin K and teriparatide on the bone metabolism in ovariectomized rats. Biomed. Rep. 2015, 3, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Harsløf, T.; Sikjær, T.; Sørensen, L.; Pedersen, S.B.; Mosekilde, L.; Langdahl, B.L.; Rejnmark, L. The Effect of Treatment With PTH on Undercarboxylated Osteocalcin and Energy Metabolism in Hypoparathyroidism. J. Clin. Endocrinol. Metab. 2015, 100, 2758–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maser, R.E.; Lenhard, M.J.; Pohlig, R.T.; Balagopal, P.B.; Abdel-Misih, R. Effect of parathyroidectomy on osteopontin and undercarboxylated osteocalcin in patients with primary hyperparathyroidism. Endocr. Res. 2018, 43, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Neradova, A.; Wasilewski, G.; Prisco, S.; Leenders, P.; Caron, M.; Welting, T.; van Rietbergen, B.; Kramann, R.; Floege, J.; Vervloet, M.G.; et al. Combining phosphate binder therapy with vitamin K2 inhibits vascular calcification in an experimental animal model of kidney failure. Nephrol. Dial. Transplant. 2022, 37, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, C.; Jie, K.S.; Knapen, M.H. Role of vitamin K in bone metabolism. Annu. Rev. Nutr. 1995, 15, 1–22. [Google Scholar] [CrossRef]
- Stafford, D.W. The vitamin K cycle. J. Thromb. Haemost. 2005, 3, 1873–1878. [Google Scholar] [CrossRef]
- Weng, S.J.; Xie, Z.J.; Wu, Z.Y.; Yan, D.Y.; Tang, J.H.; Shen, Z.J.; Li, H.; Bai, B.L.; Boodhun, V.; Dong, X.D.E.; et al. Effects of combined menaquinone-4 and PTH1-34 treatment on osetogenesis and angiogenesis in calvarial defect in osteopenic rats. Endocrine 2019, 63, 376–384. [Google Scholar] [CrossRef]
- Shearer, M.J.; Newman, P. Recent trends in the metabolism and cell biology of vitamin K with special reference to vitamin K cycling and MK-4 biosynthesis. J. Lipid Res. 2014, 55, 345–362. [Google Scholar] [CrossRef] [Green Version]
- Hirota, Y.; Nakagawa, K.; Sawada, N.; Okuda, N.; Suhara, Y.; Uchino, Y.; Kimoto, T.; Funahashi, N.; Kamao, M.; Tsugawa, N.; et al. Functional characterization of the vitamin K2 biosynthetic enzyme UBIAD1. PLoS ONE 2015, 10, e0125737. [Google Scholar] [CrossRef] [Green Version]
- Kaesler, N.; Magdeleyns, E.; Herfs, M.; Schettgen, T.; Brandenburg, V.; Fliser, D.; Vermeer, C.; Floege, J.; Schlieper, G.; Krüger, T. Impaired vitamin K recycling in uremia is rescued by vitamin K supplementation. Kidney Int. 2014, 86, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Kaesler, N.; Schreibing, F.; Speer, T.; Puente-Secades, S.; Rapp, N.; Drechsler, C.; Kabgani, N.; Kuppe, C.; Boor, P.; Jankowski, V.; et al. Altered vitamin K biodistribution and metabolism in experimental and human chronic kidney disease. Kidney Int. 2022, 101, 338–348. [Google Scholar] [CrossRef]
- Pereira, R.C.; Delany, A.M.; Khouzam, N.M.; Bowen, R.E.; Freymiller, E.G.; Salusky, I.B.; Wesseling-Perry, K. Primary osteoblast-like cells from patients with end-stage kidney disease reflect gene expression, proliferation, and mineralization characteristics ex vivo. Kidney Int. 2015, 87, 593–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balani, D.H.; Ono, N.; Kronenberg, H.M. Parathyroid hormone regulates fates of murine osteoblast precursors in vivo. J. Clin. Investig. 2017, 127, 3327–3338. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, K.; Sieklucka, B.; Pawlak, D. Paracrine Kynurenic Pathway Activation in the Bone of Young Uremic Rats Can Antagonize Anabolic Effects of PTH on Bone Turnover and Strength through the Disruption of PTH-Dependent Molecular Signaling. Int. J. Mol. Sci. 2021, 22, 6563. [Google Scholar] [CrossRef] [PubMed]
- Kalajzic, I.; Staal, A.; Yang, W.P.; Wu, Y.; Johnson, S.E.; Feyen, J.H.; Krueger, W.; Maye, P.; Yu, F.; Zhao, Y.; et al. Expression profile of osteoblast lineage at defined stages of differentiation. J. Biol. Chem. 2005, 280, 24618–24626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferron, M.; Lacombe, J.; Germain, A.; Oury, F.; Karsenty, G. GGCX and VKORC1 inhibit osteocalcin endocrine functions. J. Cell. Biol. 2019, 218, 2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkins, G.J.; Welldon, K.J.; Wijenayaka, A.R.; Bonewald, L.F.; Findlay, D.M. Vitamin K promotes mineralization, osteoblast-to-osteocyte transition, and an anticatabolic phenotype by {gamma}-carboxylation-dependent and -independent mechanisms. Am. J. Physiol. Cell Physiol. 2009, 297, C1358–C1367. [Google Scholar] [CrossRef] [Green Version]
- Boskey, A.L.; Gadaleta, S.; Gundberg, C.; Doty, S.B.; Ducy, P.; Karsenty, G. Fourier transform infrared microspectroscopic analysis of bones of osteocalcin-deficient mice provides insight into the function of osteocalcin. Bone 1998, 23, 187–196. [Google Scholar] [CrossRef]
- Roy, M.E.; Nishimoto, S.K.; Rho, J.Y.; Bhattacharya, S.K.; Lin, J.S.; Pharr, G.M. Correlations between osteocalcin content, degree of mineralization, and mechanical properties of C. carpio rib bone. J. Biomed. Mater. Res. 2001, 54, 547–553. [Google Scholar] [CrossRef]
- Lambert, L.J.; Challa, A.K.; Niu, A.; Zhou, L.; Tucholski, J.; Johnson, M.S.; Nagy, T.R.; Eberhardt, A.W.; Estep, P.N.; Kesterson, R.A.; et al. Increased trabecular bone and improved biomechanics in an osteocalcin-null rat model created by CRISPR/Cas9 technology. Dis. Model. Mech. 2016, 9, 1169–1179. [Google Scholar] [CrossRef] [Green Version]
- Ducy, P.; Desbois, C.; Boyce, B.; Pinero, G.; Story, B.; Dunstan, C.; Smith, E.; Bonadio, J.; Goldstein, S.; Gundberg, C.; et al. Increased bone formation in osteocalcin-deficient mice. Nature 1996, 382, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Hara, K.; Kobayashi, M.; Akiyama, Y. Comparison of inhibitory effects of warfarin on gamma-carboxylation between bone and liver in rats. J. Bone Miner. Metab. 2005, 23, 366–372l. [Google Scholar] [CrossRef] [PubMed]
- Amizuka, N.; Li, M.; Hara, K.; Kobayashi, M.; de Freitas, P.H.; Ubaidus, S.; Oda, K.; Akiyama, Y. Warfarin administration disrupts the assembly of mineralized nodules in the osteoid. J. Electron. Microsc. (Tokyo) 2009, 58, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Simon, P.; Grüner, D.; Worch, H.; Pompe, W.; Lichte, H.; El Khassawna, T.; Heiss, C.; Wenisch, S.; Kniep, R. First evidence of octacalcium phosphate@osteocalcin nanocomplex as skeletal bone component directing collagen triple-helix nanofibril mineralization. Sci. Rep. 2018, 8, 13696. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; Irie, K.; Fukuhara, D.; Kataoka, K.; Hattori, T.; Ono, M.; Ekuni, D.; Kubota, S.; Morita, M. Commensal Microbiota Enhance Both Osteoclast and Osteoblast Activities. Molecules 2018, 23, 1517. [Google Scholar] [CrossRef] [Green Version]
- Mor, A.; Pawlak, K.; Kalaska, B.; Domaniewski, T.; Sieklucka, B.; Zieminska, M.; Cylwik, B.; Pawlak, D. Modulation of the Paracrine Kynurenic System in Bone as a New Regulator of Osteoblastogenesis and Bone Mineral Status in an Animal Model of Chronic Kidney Disease Treated with LP533401. Int. J. Mol. Sci. 2020, 21, 5979. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Precursor ion (m/z) | Production (m/z) | Collision Energy (V) | Retention Time (min) | |
|---|---|---|---|---|
| K1 | 451.4 | 187.2 | 26 | 6.02 |
| K1-d7 | 458.4 | 194.3 | 26 | 6 |
| MK4 | 445.3 | 187.3 | 18 | 5.42 |
| MK7 | 649.5 | 187.2 | 38 | 7.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziemińska, M.; Pawlak, D.; Sieklucka, B.; Chilkiewicz, K.; Pawlak, K. Vitamin K-Dependent Carboxylation of Osteocalcin in Bone—Ally or Adversary of Bone Mineral Status in Rats with Experimental Chronic Kidney Disease? Nutrients 2022, 14, 4082. https://doi.org/10.3390/nu14194082
Ziemińska M, Pawlak D, Sieklucka B, Chilkiewicz K, Pawlak K. Vitamin K-Dependent Carboxylation of Osteocalcin in Bone—Ally or Adversary of Bone Mineral Status in Rats with Experimental Chronic Kidney Disease? Nutrients. 2022; 14(19):4082. https://doi.org/10.3390/nu14194082
Chicago/Turabian StyleZiemińska, Marta, Dariusz Pawlak, Beata Sieklucka, Katarzyna Chilkiewicz, and Krystyna Pawlak. 2022. "Vitamin K-Dependent Carboxylation of Osteocalcin in Bone—Ally or Adversary of Bone Mineral Status in Rats with Experimental Chronic Kidney Disease?" Nutrients 14, no. 19: 4082. https://doi.org/10.3390/nu14194082
APA StyleZiemińska, M., Pawlak, D., Sieklucka, B., Chilkiewicz, K., & Pawlak, K. (2022). Vitamin K-Dependent Carboxylation of Osteocalcin in Bone—Ally or Adversary of Bone Mineral Status in Rats with Experimental Chronic Kidney Disease? Nutrients, 14(19), 4082. https://doi.org/10.3390/nu14194082