
The Effects of Omega-3 Polyunsaturated Fatty Acids on Breast Cancer as a Preventive Measure or as an Adjunct to Conventional Treatments

 ,
,  , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
2.2. Inclusion Criteria
2.3. Exclusion Criteria
2.4. Data Compilation
2.5. Data Extraction
2.6. Risk of Bias Assessment
2.7. Data Analysis
3. Results
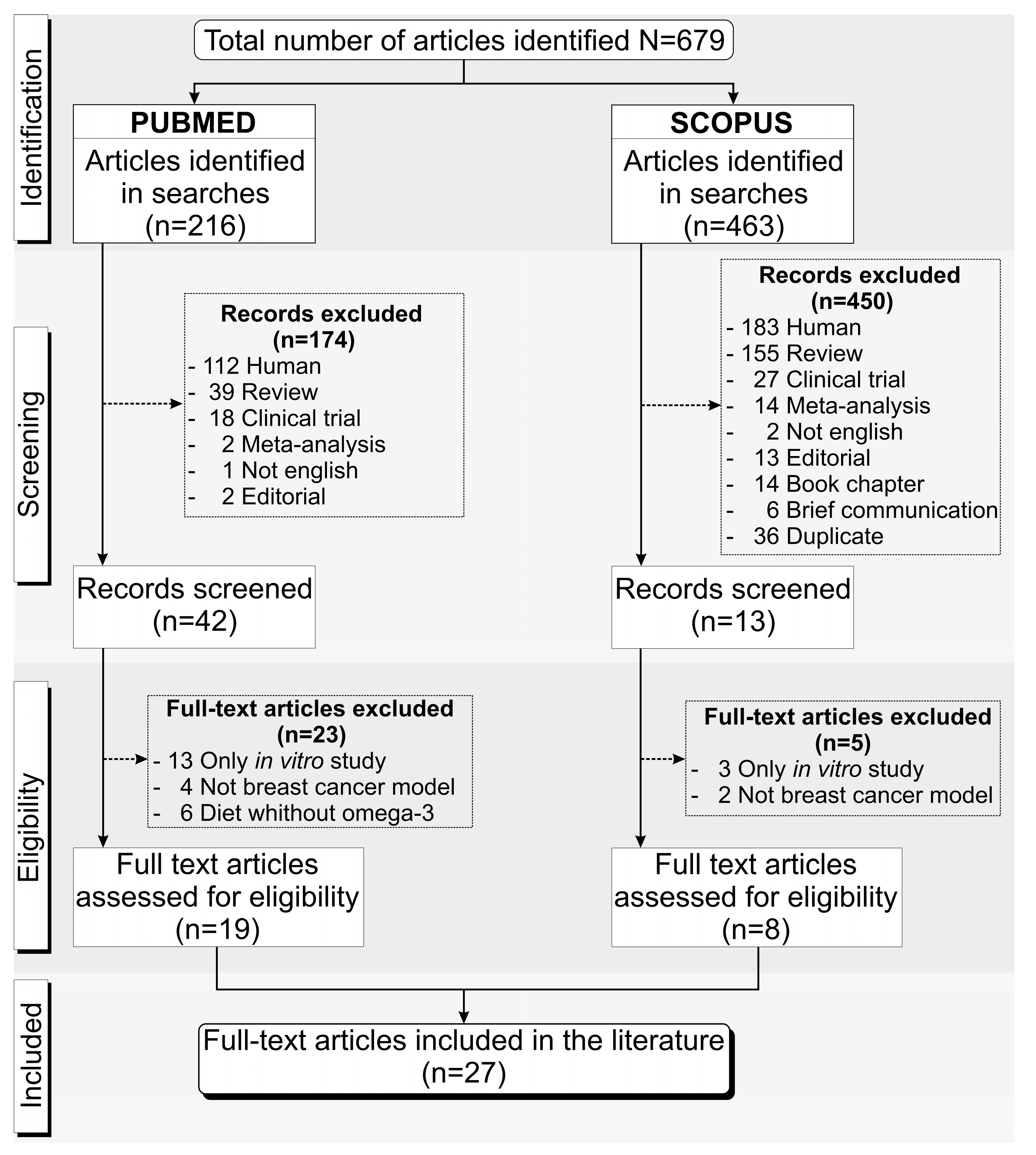
3.1. Overview of the Screening Process of the Included Studies
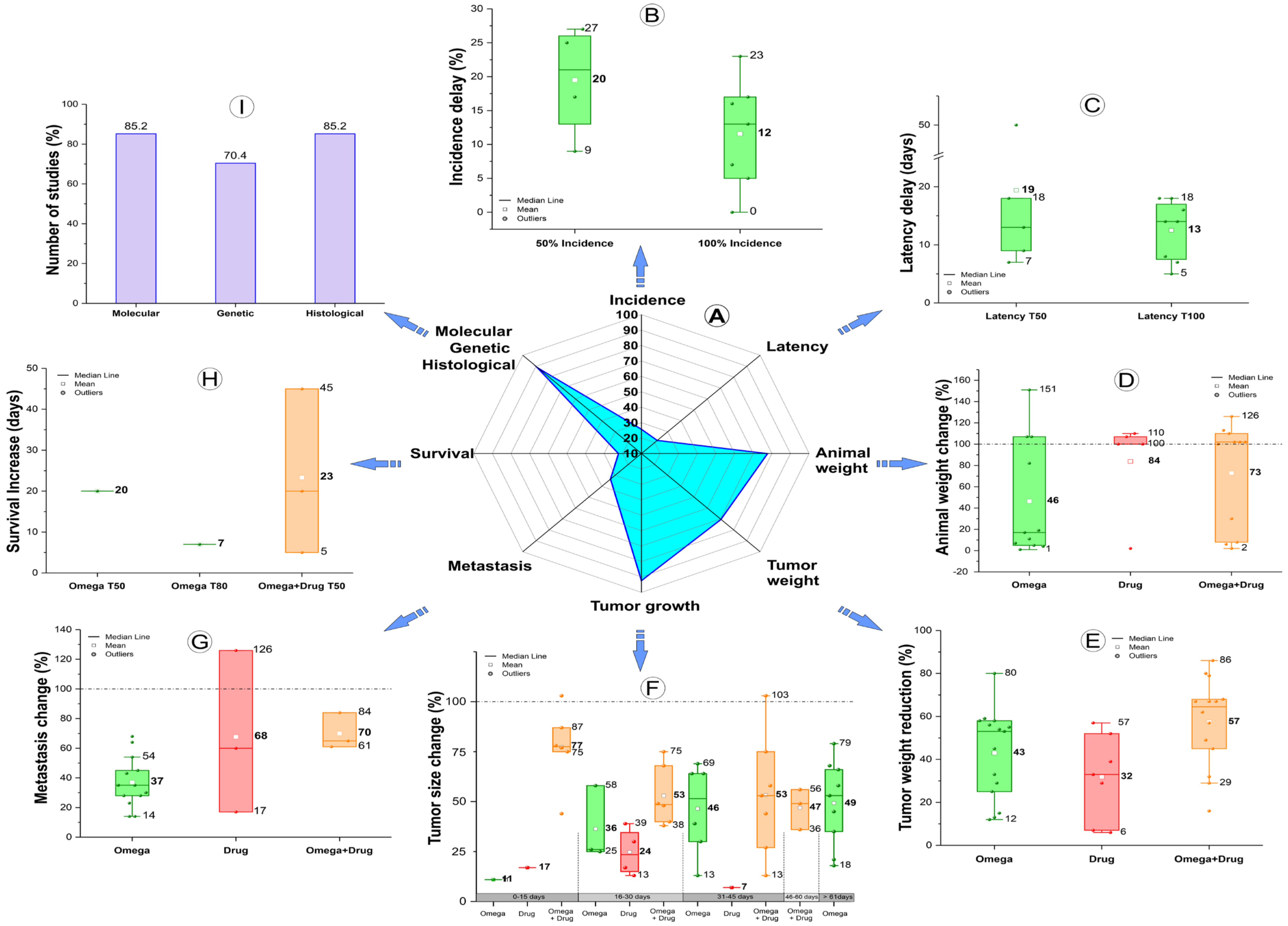
3.2. Characteristics of the Breast Cancer Experimental Animal Model
3.3. Breast Cancer Cell Transplant Models
3.4. Experimental Design and Characteristics of the Diet and Complementary Treatment
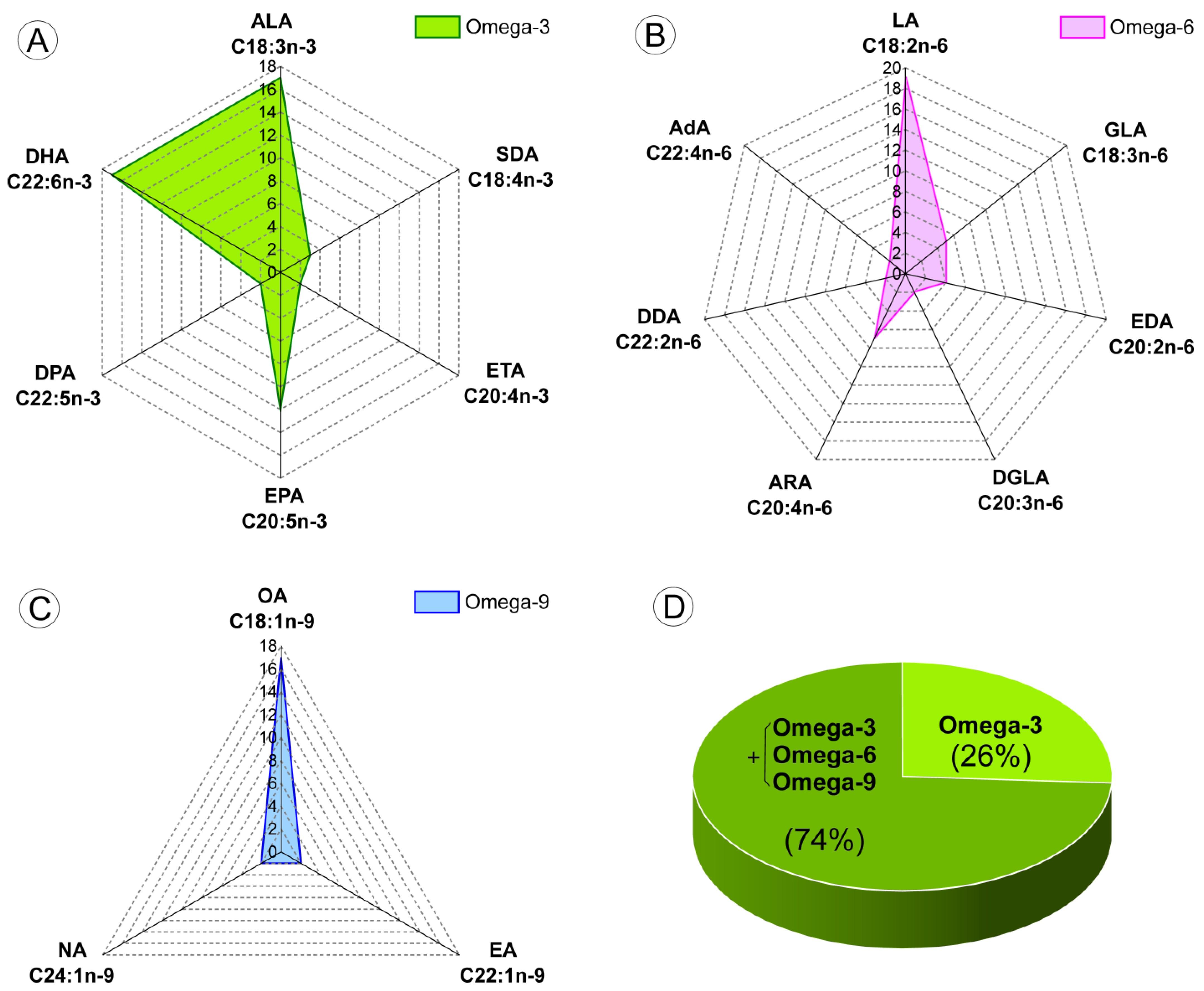
3.5. Dietary Compounds
4. Discussion
4.1. Outcomes of the Omega-3 PUFA Effect Combined with Antitumor Drugs or Not
4.2. Animal Models Used in the BC Induction
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Arnold, M.; Morgan, E.; Rumgay, H.; Mafra, A.; Singh, D.; Laversanne, M.; Vignat, J.; Gralow, J.R.; Cardoso, F.; Siesling, S.; et al. Current and future burden of breast cancer: Global statistics for 2020 and 2040. Breast 2022, 66, 15–23. [Google Scholar] [CrossRef]
- Peto, J. Cancer epidemiology in the last century and the next decade. Nature 2001, 411, 390–395. [Google Scholar] [CrossRef]
- De Cicco, P.; Catani, M.V.; Gasperi, V.; Sibilano, M.; Quaglietta, M.; Savini, I. Nutrition and Breast Cancer: A Literature Review on Prevention, Treatment and Recurrence. Nutrients 2019, 11, 1514. [Google Scholar] [CrossRef] [Green Version]
- Warburton, D.E.; Bredin, S.S. Reflections on Physical Activity and Health: What Should We Recommend? Can. J. Cardiol. 2016, 32, 495–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, C.; Milne, G.L.; Sandler, D.P.; Nichols, H.B. Oxidative stress in relation to diet and physical activity among premenopausal women. Br. J. Nutr. 2016, 116, 1416–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delort, L.; Rossary, A.; Farges, M.C.; Vasson, M.P.; Caldefie-Chézet, F. Leptin, adipocytes and breast cancer: Focus on inflammation and anti-tumor immunity. Life Sci. 2015, 140, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Nathanson, S.D.; Detmar, M.; Padera, T.P.; Yates, L.R.; Welch, D.R.; Beadnell, T.C.; Scheid, A.D.; Wrenn, E.D.; Cheung, K. Mechanisms of breast cancer metastasis. Clin. Exp. Metastasis 2022, 39, 117–137. [Google Scholar] [CrossRef]
- INCA, I.N.d.C. Conceito e Magnitude. Definição do Câncer de Mama e Dados de Incidência e Mortalidade no Brasil. Available online: https://www.gov.br/inca/pt-br/search?SearchableText=cancer%20de%20mama (accessed on 15 February 2023).
- Garrido-Castro, A.C.; Lin, N.U.; Polyak, K. Insights into Molecular Classifications of Triple-Negative Breast Cancer: Improving Patient Selection for Treatment. Cancer Discov 2019, 9, 176–198. [Google Scholar] [CrossRef] [Green Version]
- Katsura, C.; Ogunmwonyi, I.; Kankam, H.K.; Saha, S. Breast cancer: Presentation, investigation and management. Br. J. Hosp. Med. 2022, 83, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Doan, L.M.T.; Angione, C.; Occhipinti, A. Machine Learning Methods for Survival Analysis with Clinical and Transcriptomics Data of Breast Cancer. Methods Mol. Biol. 2023, 2553, 325–393. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, R.A.; Harvey, K.A.; Xu, Z.; Bammerlin, E.M.; Walker, C.; Altenburg, J.D. Docosahexaenoic acid: A natural powerful adjuvant that improves efficacy for anticancer treatment with no adverse effects. Biofactors 2011, 37, 399–412. [Google Scholar] [CrossRef]
- Pal, S.K.; Childs, B.H.; Pegram, M. Triple negative breast cancer: Unmet medical needs. Breast Cancer Res. Treat. 2011, 125, 627–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurvitz, S.; Mead, M. Triple-negative breast cancer: Advancements in characterization and treatment approach. Curr. Opin. Obstet. Gynecol. 2016, 28, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Haffty, B.G.; Yang, Q.; Reiss, M.; Kearney, T.; Higgins, S.A.; Weidhaas, J.; Harris, L.; Hait, W.; Toppmeyer, D. Locoregional relapse and distant metastasis in conservatively managed triple negative early-stage breast cancer. J. Clin. Oncol. 2006, 24, 5652–5657. [Google Scholar] [CrossRef] [PubMed]
- Newell, M.; Goruk, S.; Schueler, J.; Mazurak, V.; Postovit, L.M.; Field, C.J. Docosahexaenoic acid enrichment of tumor phospholipid membranes increases tumor necroptosis in mice bearing triple negative breast cancer patient-derived xenografts. J. Nutr. Biochem. 2022, 107, 109018. [Google Scholar] [CrossRef]
- Li, B.; Tan, T.; Chu, W.; Zhang, Y.; Ye, Y.; Wang, S.; Qin, Y.; Tang, J.; Cao, X. Co-delivery of paclitaxel (PTX) and docosahexaenoic acid (DHA) by targeting lipid nanoemulsions for cancer therapy. Drug Deliv. 2022, 29, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Chen, C.Y.; Li, X.; Zhang, X.; Su, C.W.; Liu, Y.; Cao, T.; Hao, L.; Wang, M.; Kang, J.X. Increased lipogenesis is critical for self-renewal and growth of breast cancer stem cells: Impact of omega-3 fatty acids. Stem Cells 2021, 39, 1660–1670. [Google Scholar] [CrossRef]
- Fabian, C.J.; Befort, C.A.; Phillips, T.A.; Nydegger, J.L.; Kreutzjans, A.L.; Powers, K.R.; Metheny, T.; Klemp, J.R.; Carlson, S.E.; Sullivan, D.K.; et al. Change in Blood and Benign Breast Biomarkers in Women Undergoing a Weight-Loss Intervention Randomized to High-Dose ω-3 Fatty Acids versus Placebo. Cancer Prev. Res. 2021, 14, 893–904. [Google Scholar] [CrossRef]
- Goupille, C.; Frank, P.G.; Arbion, F.; Jourdan, M.L.; Guimaraes, C.; Pinault, M.; Body, G.; Chevalier, S.; Bougnoux, P.; Ouldamer, L. Low Levels of Omega-3 Long-Chain Polyunsaturated Fatty Acids Are Associated with Bone Metastasis Formation in Premenopausal Women with Breast Cancer: A Retrospective Study. Nutrients 2020, 12, 3832. [Google Scholar] [CrossRef]
- Augimeri, G.; Montalto, F.I.; Giordano, C.; Barone, I.; Lanzino, M.; Catalano, S.; Andò, S.; De Amicis, F.; Bonofiglio, D. Nutraceuticals in the Mediterranean Diet: Potential Avenues for Breast Cancer Treatment. Nutrients 2021, 13, 2557. [Google Scholar] [CrossRef]
- Darwito, D.; Dharmana, E.; Riwanto, I.; Budijitno, S.; Suwardjo, S.; Purnomo, J.; Widodo, I.; Ghozali, A.; Aryandono, T.; Anwar, S.L. Effects of Omega-3 Supplementation on Ki-67 and VEGF Expression Levels and Clinical Outcomes of Locally Advanced Breast Cancer Patients Treated with Neoadjuvant CAF Chemotherapy: A Randomized Controlled Trial Report. Asian Pac. J. Cancer Prev. 2019, 20, 911–916. [Google Scholar] [CrossRef]
- Fabian, C.J.; Kimler, B.F.; Hursting, S.D. Omega-3 fatty acids for breast cancer prevention and survivorship. Breast Cancer Res. 2015, 17, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanckaert, V.; Ulmann, L.; Mimouni, V.; Antol, J.; Brancquart, L.; Chénais, B. Docosahexaenoic acid intake decreases proliferation, increases apoptosis and decreases the invasive potential of the human breast carcinoma cell line MDA-MB-231. Int. J. Oncol. 2010, 36, 737–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslan, C.; Maralbashi, S.; Kahroba, H.; Asadi, M.; Soltani-Zangbar, M.S.; Javadian, M.; Shanehbandi, D.; Baradaran, B.; Darabi, M.; Kazemi, T. Docosahexaenoic acid (DHA) inhibits pro-angiogenic effects of breast cancer cells via down-regulating cellular and exosomal expression of angiogenic genes and microRNAs. Life Sci. 2020, 258, 118094. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; Cicero, A.F.; Fogari, E.; D’Angelo, A.; Bonaventura, A.; Romano, D.; Maffioli, P. Effects of n-3 PUFAs on postprandial variation of metalloproteinases, and inflammatory and insulin resistance parameters in dyslipidemic patients: Evaluation with euglycemic clamp and oral fat load. J. Clin. Lipidol. 2012, 6, 553–564. [Google Scholar] [CrossRef]
- Spencer, L.; Mann, C.; Metcalfe, M.; Webb, M.; Pollard, C.; Spencer, D.; Berry, D.; Steward, W.; Dennison, A. The effect of omega-3 FAs on tumour angiogenesis and their therapeutic potential. Eur. J. Cancer 2009, 45, 2077–2086. [Google Scholar] [CrossRef]
- Chen, H.W.; Chao, C.Y.; Lin, L.L.; Lu, C.Y.; Liu, K.L.; Lii, C.K.; Li, C.C. Inhibition of matrix metalloproteinase-9 expression by docosahexaenoic acid mediated by heme oxygenase 1 in 12-O-tetradecanoylphorbol-13-acetate-induced MCF-7 human breast cancer cells. Arch. Toxicol. 2013, 87, 857–869. [Google Scholar] [CrossRef]
- Ravacci, G.R.; Brentani, M.M.; Tortelli, T.; Torrinhas, R.S.M.M.; Saldanha, T.; Torres, E.A.F.S.; Waitzberg, D.L. Lipid raft disruption by docosahexaenoic acid induces apoptosis in transformed human mammary luminal epithelial cells harboring HER-2 overexpression. J. Nutr. Biochem. 2013, 24, 505–515. [Google Scholar] [CrossRef]
- Ravacci, G.R.; Brentani, M.M.; Tortelli, T.C.; Torrinhas, R.S.; Santos, J.R.; Logullo, A.F.; Waitzberg, D.L. Docosahexaenoic Acid Modulates a HER2-Associated Lipogenic Phenotype, Induces Apoptosis, and Increases Trastuzumab Action in HER2-Overexpressing Breast Carcinoma Cells. Biomed. Res. Int. 2015, 2015, 838652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paixão, E.; Oliveira, A.C.M.; Pizato, N.; Muniz-Junqueira, M.I.; Magalhães, K.G.; Nakano, E.Y.; Ito, M.K. The effects of EPA and DHA enriched fish oil on nutritional and immunological markers of treatment naïve breast cancer patients: A randomized double-blind controlled trial. Nutr. J. 2017, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Unger, J.M.; Crew, K.D.; Till, C.; Greenlee, H.; Gralow, J.; Dakhil, S.R.; Minasian, L.M.; Wade, J.L., 3rd; Fisch, M.J.; et al. Omega-3 fatty acid use for obese breast cancer patients with aromatase inhibitor-related arthralgia (SWOG S0927). Breast Cancer Res. Treat. 2018, 172, 603–610. [Google Scholar] [CrossRef]
- Pauwels, E.K.; Kairemo, K. Fatty acid facts, part II: Role in the prevention of carcinogenesis, or, more fish on the dish? Drug News Perspect. 2008, 21, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Wendel, M.; Heller, A.R. Anticancer actions of omega-3 fatty acids--current state and future perspectives. Anticancer. Agents Med. Chem. 2009, 9, 457–470. [Google Scholar] [CrossRef]
- Martínez, N.; Herrera, M.; Frías, L.; Provencio, M.; Pérez-Carrión, R.; Díaz, V.; Morse, M.; Crespo, M.C. A combination of hydroxytyrosol, omega-3 fatty acids and curcumin improves pain and inflammation among early stage breast cancer patients receiving adjuvant hormonal therapy: Results of a pilot study. Clin. Transl. Oncol. 2019, 21, 489–498. [Google Scholar] [CrossRef]
- Bjorklund, G. The Adjuvant Nutritional Intervention in Cancer (ANICA) Trial. Nutr. Cancer 2015, 67, 1357–1360. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Yang, H.; Khan, A.R.; Yang, X.; Xu, J.; Ji, J.; Zhai, G. Redox-responsive hyaluronic acid-based nanoparticles for targeted photodynamic therapy/chemotherapy against breast cancer. J. Colloid Interface Sci. 2021, 598, 213–228. [Google Scholar] [CrossRef]
- Monk, J.M.; Liddle, D.M.; Hutchinson, A.L.; Burns, J.L.; Wellings, H.; Cartwright, N.M.; Muller, W.J.; Power, K.A.; Robinson, L.E.; Ma, D.W.L. Fish oil supplementation increases expression of mammary tumor apoptosis mediators and reduces inflammation in an obesity-associated HER-2 breast cancer model. J. Nutr. Biochem. 2021, 95, 108763. [Google Scholar] [CrossRef]
- Ion, G.; Akinsete, J.A.; Witte, T.R.; Bostan, M.; Hardman, W.E. Maternal fish oil consumption has a negative impact on mammary gland tumorigenesis in C3(1) Tag mice offspring. Eur. J. Nutr. 2021, 60, 3771–3781. [Google Scholar] [CrossRef]
- Guo, C.H.; Hsia, S.; Chung, C.H.; Lin, Y.C.; Shih, M.Y.; Chen, P.C.; Peng, C.L.; Henning, S.M.; Hsu, G.W.; Li, Z. Nutritional supplements in combination with chemotherapy or targeted therapy reduces tumor progression in mice bearing triple-negative breast cancer. J. Nutr. Biochem. 2021, 87, 108504. [Google Scholar] [CrossRef] [PubMed]
- Garay, M.I.; Comba, A.; Vara Messler, M.; Barotto, N.N.; Silva, R.A.; Repossi, G.; Quiroga, P.L.; Mazzudulli, G.M.; Brunotto, M.N.; Pasqualini, M.E. Tumorigenic effect mediated by ROS/eicosanoids and their regulation on TP53 expression in a murine mammary gland adenocarcinoma. Prostaglandins Other Lipid Mediat. 2021, 155, 106564. [Google Scholar] [CrossRef]
- Abbas, A.; Witte, T.; Patterson, W.L., 3rd; Fahrmann, J.F.; Guo, K.; Hur, J.; Hardman, W.E.; Georgel, P.T. Epigenetic Reprogramming Mediated by Maternal Diet Rich in Omega-3 Fatty Acids Protects From Breast Cancer Development in F1 Offspring. Front. Cell Dev. Biol. 2021, 9, 682593. [Google Scholar] [CrossRef] [PubMed]
- Newell, M.; Patel, D.; Goruk, S.; Field, C.J. Docosahexaenoic Acid Incorporation Is Not Affected by Doxorubicin Chemotherapy in either Whole Cell or Lipid Raft Phospholipids of Breast Cancer Cells in vitro and Tumor Phospholipids in vivo. Lipids 2020, 55, 549–565. [Google Scholar] [CrossRef]
- Liu, L.; Jin, R.; Hao, J.; Zeng, J.; Yin, D.; Yi, Y.; Zhu, M.; Mandal, A.; Hua, Y.; Ng, C.K.; et al. Consumption of the Fish Oil High-Fat Diet Uncouples Obesity and Mammary Tumor Growth through Induction of Reactive Oxygen Species in Protumor Macrophages. Cancer Res. 2020, 80, 2564–2574. [Google Scholar] [CrossRef] [Green Version]
- Hillyer, L.M.; Hucik, B.; Baracuhy, E.M.; Lin, Z.; Muller, W.J.; Robinson, L.E.; Ma, D.W.L. Her-2 Breast Cancer Outcomes Are Mitigated by Consuming n-3 Polyunsaturated, Saturated, and Monounsaturated Fatty Acids Compared to n-6 Polyunsaturated Fatty Acids. Nutrients 2020, 12, 3901. [Google Scholar] [CrossRef]
- Goupille, C.; Vibet, S.; Frank, P.G.; Maheo, K. EPA and DHA Fatty Acids Induce a Remodeling of Tumor Vasculature and Potentiate Docetaxel Activity. Int. J. Mol. Sci. 2020, 21, 4965. [Google Scholar] [CrossRef] [PubMed]
- Torres-Adorno, A.M.; Vitrac, H.; Qi, Y.; Tan, L.; Levental, K.R.; Fan, Y.Y.; Yang, P.; Chapkin, R.S.; Eckhardt, B.L.; Ueno, N.T. Eicosapentaenoic acid in combination with EPHA2 inhibition shows efficacy in preclinical models of triple-negative breast cancer by disrupting cellular cholesterol efflux. Oncogene 2019, 38, 2135–2150. [Google Scholar] [CrossRef]
- Newell, M.; Goruk, S.; Mazurak, V.; Postovit, L.; Field, C.J. Role of docosahexaenoic acid in enhancement of docetaxel action in patient-derived breast cancer xenografts. Breast Cancer Res. Treat. 2019, 177, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Newell, M.; Brun, M.; Field, C.J. Treatment with DHA Modifies the Response of MDA-MB-231 Breast Cancer Cells and Tumors from nu/nu Mice to Doxorubicin through Apoptosis and Cell Cycle Arrest. J. Nutr. 2019, 149, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, K.; Gao, J.; Guo, X.; Lu, M.; Li, Z.; Li, D. Endogenously Synthesized n-3 Polyunsaturated Fatty Acids in Pregnant fat-1 Mice Decreases Mammary Cancer Risk of Female Offspring by Regulating Expression of Long Noncoding RNAs. Mol. Nutr. Food Res. 2019, 63, e1801150. [Google Scholar] [CrossRef]
- Fernando, W.; Coyle, K.; Marcato, P.; Vasantha Rupasinghe, H.P.; Hoskin, D.W. Phloridzin docosahexaenoate, a novel fatty acid ester of a plant polyphenol, inhibits mammary carcinoma cell metastasis. Cancer Lett. 2019, 465, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Bialek, M.; Bialek, A.; Czauderna, M. Conjugated Linoleic Acid Isomers Affect Profile of Lipid Compounds and Intensity of Their Oxidation in Heart of Rats with Chemically-Induced Mammary Tumors-Preliminary Study. Nutrients 2019, 11, 2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Feng, N.; Lin, G.; Tong, Y.; Jiang, X.; Yang, Q.; Wang, S.; Chen, W.; He, Z.; Chen, Y.Q. Metabolic Shift Induced by ω -3 PUFAs and Rapamycin Lead to Cancer Cell Death. Cell Physiol. Biochem. 2018, 48, 2318–2336. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Abdelmagid, S.A.; Pinelli, C.J.; Monk, J.M.; Liddle, D.M.; Hillyer, L.M.; Hucik, B.; Silva, A.; Subedi, S.; Wood, G.A.; et al. Marine fish oil is more potent than plant-based n-3 polyunsaturated fatty acids in the prevention of mammary tumors. J. Nutr. Biochem. 2018, 55, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Khadge, S.; Thiele, G.M.; Sharp, J.G.; McGuire, T.R.; Klassen, L.W.; Black, P.N.; DiRusso, C.C.; Cook, L.; Talmadge, J.E. Long-chain omega-3 polyunsaturated fatty acids decrease mammary tumor growth, multiorgan metastasis and enhance survival. Clin. Exp. Metastasis 2018, 35, 797–818. [Google Scholar] [CrossRef]
- Jiao, Y.; Watts, T.; Xue, J.; Hannafon, B.; Ding, W.Q. Sorafenib and docosahexaenoic acid act in synergy to suppress cancer cell viability: A role of heme oxygenase 1. BMC Cancer 2018, 18, 1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Lin, G.; Song, C.; Wu, Y.; Feng, N.; Chen, W.; He, Z.; Chen, Y.Q. RA and ω-3 PUFA co-treatment activates autophagy in cancer cells. Oncotarget 2017, 8, 109135–109150. [Google Scholar] [CrossRef] [Green Version]
- Vara-Messler, M.; Pasqualini, M.E.; Comba, A.; Silva, R.; Buccellati, C.; Trenti, A.; Trevisi, L.; Eynard, A.R.; Sala, A.; Bolego, C.; et al. Increased dietary levels of α-linoleic acid inhibit mammary tumor growth and metastasis. Eur. J. Nutr. 2017, 56, 509–519. [Google Scholar] [CrossRef]
- Jiao, Y.; Hannafon, B.N.; Zhang, R.R.; Fung, K.M.; Ding, W.Q. Docosahexaenoic acid and disulfiram act in concert to kill cancer cells: A mutual enhancement of their anticancer actions. Oncotarget 2017, 8, 17908–17920. [Google Scholar] [CrossRef] [Green Version]
- Dyari, H.R.E.; Rawling, T.; Chen, Y.; Sudarmana, W.; Bourget, K.; Dwyer, J.M.; Allison, S.E.; Murray, M. A novel synthetic analogue of ω-3 17,18-epoxyeicosatetraenoic acid activates TNF receptor-1/ASK1/JNK signaling to promote apoptosis in human breast cancer cells. Faseb J. 2017, 31, 5246–5257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, C.; Tewari, S.; Chakraborty, D.; Vaishnav, S. Omega-3 Fatty Acid and its Protective Effect against Cancer and Cancer-related Complication. J. Pharm. Res. Int. 2022, 34, 51–62. [Google Scholar] [CrossRef]
- Saadatian-Elahi, M.; Norat, T.; Goudable, J.; Riboli, E. Biomarkers of dietary fatty acid intake and the risk of breast cancer: A meta-analysis. Int. J. Cancer 2004, 111, 584–591. [Google Scholar] [CrossRef]
- Delpino, F.M.; Figueiredo, L.M. Effects of omega-3 supplementation on lean body mass in cancer patients: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2022, 76, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Global Recommendations for EPA and DHA Intake (Rev 19 November 2014). Available online: https://www.issfal.org/assets/globalrecommendationssummary19nov2014landscape_-3-.pdf (accessed on 12 August 2022).
- Intake Recommendations. Available online: https://goedomega3.com/intake-recommendations (accessed on 12 August 2022).
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467s–1476s. [Google Scholar] [CrossRef] [Green Version]
- Bobin-Dubigeon, C.; Nazih, H.; Croyal, M.; Bard, J.M. Link between Omega 3 Fatty Acids Carried by Lipoproteins and Breast Cancer Severity. Nutrients 2022, 14, 2461. [Google Scholar] [CrossRef]
- Sonestedt, E.; Ericson, U.; Gullberg, B.; Skog, K.; Olsson, H.; Wirfält, E. Do both heterocyclic amines and omega-6 polyunsaturated fatty acids contribute to the incidence of breast cancer in postmenopausal women of the Malmö diet and cancer cohort? Int. J. Cancer 2008, 123, 1637–1643. [Google Scholar] [CrossRef]
- Signori, C.; El-Bayoumy, K.; Russo, J.; Thompson, H.J.; Richie, J.P.; Hartman, T.J.; Manni, A. Chemoprevention of breast cancer by fish oil in preclinical models: Trials and tribulations. Cancer Res. 2011, 71, 6091–6096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.; Kim, H. Anti-cancer Mechanism of Docosahexaenoic Acid in Pancreatic Carcinogenesis: A Mini-review. J. Cancer Prev. 2017, 22, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Arshad, A.; Chung, W.Y.; Steward, W.; Metcalfe, M.S.; Dennison, A.R. Reduction in circulating pro-angiogenic and pro-inflammatory factors is related to improved outcomes in patients with advanced pancreatic cancer treated with gemcitabine and intravenous omega-3 fish oil. HPB 2013, 15, 428–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Albert, C.M.; Gordon, D.; Copeland, T.; et al. Marine n-3 Fatty Acids and Prevention of Cardiovascular Disease and Cancer. N. Engl. J. Med. 2019, 380, 23–32. [Google Scholar] [CrossRef]
- Soni, S.; Torvund, M.; Mandal, C.C. Omega-3 Fatty Acid Treatment Combined with Chemotherapy to Prevent Toxicity, Drug Resistance, and Metastasis in Cancer. Curr. Drug Targets 2022, 23, 574–596. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Dong, Y.; Wang, Y.; Hu, P.; Wang, J.; Wang, R.Y. Pristimerin exerts antitumor activity against MDA-MB-231 triple-negative breast cancer cells by reversing of epithelial-mesenchymal transition via downregulation of integrin β3. Biomed. J. 2021, 44, S84–S92. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Li, W.; Chen, C.S. Breast cancer animal models and applications. Zool. Res. 2020, 41, 477–494. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ref | Year | Characteristics of Animal Model | Animal Description | |||||
|---|---|---|---|---|---|---|---|---|
| Type of Model | Source of Tumor | Specie | Strain | Genotype | Sex | Age (Week) | ||
| Newell et al. [17] | 2022 | Induction by cell | Human | Mice | NSG | NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ | F | 6 |
| Li et al. [18] | 2022 | Induction by cell | Human | Mice | BALB/c (Nude) | nu/nu | F | 3–4 |
| Wang et al. [39] | 2021 | Induction by cell | Mice | Mice | BALB/c | Wildtype | F | 6–7 |
| Monk et al. [40] | 2021 | Transgenic | Spontaneous | Mice | FVB/N × MMTV | MMTV-NeundlYD5 | F | 4 |
| Luo et al. [19] | 2021 | Induction by cell | Human | Mice | BALB/c (Nude) | J:nu | M | NR |
| Ion et al. [41] | 2021 | Transgenic | Spontaneous | Mice | SV129 × c(3)1-TAg | Hemizygous pups | F | 3 |
| Guo et al. [42] | 2021 | Induction by cell | Mice | Mice | BALB/c | Wildtype | F | 8 |
| Garay et al. [43] | 2021 | Induction by cell | Mice | Mice | BALB/c | Wildtype | F/M | NR |
| Abbas et al. [44] | 2021 | Induction by drug | DMBA drug | Mice | BALB/c | Wildtype | F | ~7.1 |
| Newell et al. [45] | 2020 | Induction by cell | Human | Mice | BALB/c (Nude) | nu/nu | F | 6 |
| Liu et al. [46] | 2020 | Induction by cell | Mice | Mice | C57BL/6 | A−FABP−/− | NR | NR |
| C57BL/6 | Wildtype | |||||||
| Hillyer et al. [47] | 2020 | Transgenic | Spontaneous | Mice | FVB/N × MMTV | MMTV-NeundlYD5 | F | NR |
| Goupille, et al. [48] | 2020 | Induction by drug | NMU drug | Rats | Sprague-Dawley | Wildtype | F | 6 |
| Torres-Adorno et al. [49] | 2019 | Induction by cell | Human | Mice | NSG | NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ | F | 4–6 |
| Newell et al. [50] | 2019 | Induction by cell | Human | Mice | NSG | NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ | F | 6 |
| Newell et al. [51] | 2019 | Induction by cell | Human | Mice | BALB/c (Nude) | nu/nu | F | 6 |
| Li et al. [52] | 2019 | Induction by drug | DMBA drug | Mice | Fat-1 × C57BL/6J | Wild-genotype offspring | F | 3 |
| Fernando et al. [53] | 2019 | Induction by cell | Human | Mice | NSG | Wildtype | F | 6–8 |
| Induction by cell | Mice | BALB/c | Wildtype | |||||
| Białek et al. [54] | 2019 | Induction by drug | DMBA drug | Rats | Sprague-Dawley | Wildtype | F | ~5.3 |
| Zhu et al. [55] | 2018 | Induction by cell | Human | Mice | BALB/c (Nude) | nu/nu | F | 4–5 |
| Transgenic | Spontaneous | FVB/N × MMTV | FVB/N-Tg(MMTV-PyVT)634Mul | 6 | ||||
| Liu et al. [56] | 2018 | Transgenic | Spontaneous | Mice | FVB/N × MMTV+/− | MMTV-NeundlYD6 | F | 3 |
| Khadge et al. [57] | 2018 | Induction by cell | Mice | Mice | BALB/c | Wildtype | F | 6 |
| Jiao et al. [58] | 2018 | Induction by cell | Human | Mice | BALB/c (Nude) | Foxn1nu | F | 5 |
| Zhu et al. [59] | 2017 | Induction by cell | Human | Mice | BALB/c (Nude) | nu/nu | F | 4–5 |
| Vara-Messler et al. [60] | 2017 | Induction by cell | Human | Mice | BALB/c | Wildtype | F/M | NR |
| Jiao et al. [61] | 2017 | Induction by cell | Human | Mice | BALB/c (Nude) | nu/nu | F | 5 |
| Dyari et al. [62] | 2017 | Induction by cell | Human | Mice | BALB/c (Nude) | nu/nu | F | 6 |
| Ref | Cell Characteristics | Breast Cancer Induction Tumor | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Tumor Tissue | Cell (Tumor Subtype) | Culture Medium % FBS | Strain | Cell Number | Administration Volume (µL) | Vehicle | Local Administration | Grafts | |
| Newell et al. [17] | Adenocarcinoma | MAXF401 (TNBC) | NR | NSG modified | NA | 30 mm3 | NA | Left flank | Xenograft |
| Invasive ductal carcinoma | MAXF574 (TNBC) | ||||||||
| Li et al. [18] | Mammary adenocarcinoma | MCF7 (Luminal A) | DMEM 10% FBS | BALB/c (Nude) | 5 × 105 | 80 | NR | Right flank | Xenograft |
| Wang et al. [39] | Mammary adenocarcinoma | 4T1 (TNBC) | NR | BALB/c | 107 | NR | NR | Armpit | Allograft |
| Luo et al. [19] | Mammary adenocarcinoma | MCF7-CSC (Luminal A) | DMEM F12 FBS-free | BALB/c (Nude) modified | 1.5 × 107 | 150 | PBS | Lower right flank | Xenograft |
| Guo et al. [42] | Mammary adenocarcinoma | 4T1 (TNBC) | RPMI 1640 | BALB/c modified | 105 | 100 | RPM-1640 | Right hind thigh | Allograft |
| Garay et al. [43] | Mammary adenocarcinoma | LMM3 (TNBC) | MEM 10% FBS | BALB/c | 106 | NR | NR | Right flank | Allograft |
| Newell et al. [45] | Mammary adenocarcinoma | MDA-MB-231 (TNBC) | Iscove’s MD medium 5% FCS | BALB/c (Nude) | 2 × 106 | 100 | Iscove’s MD medium 5% FCS | Below the upper right scapula | Xenograft |
| Liu et al. [46] | Mammary adenocarcinoma | E0771 (Luminal B) | RPMI 1640 5% FBS | C57BL/6 modified | 5 × 105 | NR | NR | Fat pad of the 4th mammary gland | Allograft |
| MMT060562 (TNBC) | C57BL/6 | ||||||||
| Torres-Adorno et al. [49] | Mammary adenocarcinoma | SUM149PT (TNBC) | DMEM F12 5% FBS | NSG modified | 5 × 105 | 100 | NR | Fourth inguinal mammary fat pad | Xenograft |
| BCX010 (TNBC) | |||||||||
| Newell et al. [50] | Invasive ductal carcinoma | MAXF574 (TNBC) | NR | NSG modified | NR | NR | NR | Left flank | Xenograft |
| Adenocarcinoma | MAXF401 (TNBC) | ||||||||
| Newell et al. [51] | Mammary adenocarcinoma | MDA-MB-231 (TNBC) | Iscove’s MD medium 5% FCS | BALB/c (Nude) | 2 × 106 | 100 | Iscove’s MD medium 5% FCS | Below the upper right scapula | Xenograft |
| Fernando et al. [53] | Mammary adenocarcinoma | GFP-MDA-MB-231 (TNBC) | DMEM 10% FBS | NSG | 2 × 106 | 50 | PBS and Matrigel (1:1) | Left inguinal mammary fat pad | Xenograft |
| Mammary adenocarcinoma | 4T1 (TNBC) | BALB/c | 105 | PBS | Allograft | ||||
| Zhu et al. [55] | Mammary adenocarcinoma | MDA-MB-231 (TNBC) | DMEM 10% FBS | BALB/c (Nude) | 4 × 105 | NR | NR | NR | Xenograft |
| Khadge et al. [57] | Mammary adenocarcinoma | 4T1 (TNBC) | DMEM 10% FBS | BALB/c | 5 × 103 | 100 | CMF-HBSS | Left inguinal 5th mammary fat pad | Allograft |
| Jiao et al. [58] | Mammary adenocarcinoma | MDA-MB-231 (TNBC) | DMEM 10% FBS | BALB/c (Nude) modified | 3 × 106 | 100 | PBS with 20% Matrigel | Flanks | Xenograft |
| Zhu et al. [59] | Mammary adenocarcinoma | MDA-MB-231 (TNBC) | DMEM 10% FBS | BALB/c (Nude) | 4 × 105 | NR | NR | NR | Xenograft |
| Vara-Messler et al. [60] | Mammary adenocarcinoma | LM3 * | MEM 10% FBS | BALB/c | 2.5 × 105 | 200 | MEM | Left flank | Allograft |
| 5 × 105 | |||||||||
| 106 | |||||||||
| Jiao et al. [61] | Mammary adenocarcinoma | MDA-MB-231 (TNBC) | DMEM 10% FBS | BALB/c (Nude) | 106 | 100 | PBS with matrigel | Left flank | Xenograft |
| 5 × 106 | |||||||||
| Dyari et al. [62] | Mammary adenocarcinoma | MDA-MB-231 (TNBC) | DMEM 10% FBS | BALB/c (Nude) | 4 × 104 | 100 | Ice-cold PBS with Matrigel (1:1) | Left inguinal mammary gland | Xenograft |
| Ref | Groups | n/N | Diet | FAs supplement | Treatment | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FAs Proposal | ω-3 (g/kg) | ω-6 (g/kg) | ω-9 (g/kg) | Source | Via | Time (Week) | Drug/Technique | Time | ||||
| Newell et al. [17] | Control | 8/32 | Standard | Prevention and treatment | 4.8 | 42.84 | 72.8 | Oil of lard, vegetable, canola, olive, flax and Arasco | Oral | 7 | 0.9% saline or DTX–IP | 6 (2×/w) |
| Low and high DHA | 14.4 | 27/31.2 | 81.6/77.4 | Canola Oil/DHAsco + vegetable oil | DTX–IP | |||||||
| Li et al. [18] | Control | 6/42 | NR | Treatment | NA | NA | NA | NA | NA | Single dose | Saline or Taxol or PTX LN | Single dose |
| DHA/FA/LNs | NR or FA | NR | Soybean oil, cholesterol, egg phosphatidylcholine and Croda Inc. | IV | PTX by LN– IV | |||||||
| Wang et al. [39] | Control or Ce6 | 5/30 | NR + HA | Treatment | NA | NA | NA | Innochem® | IV | Single dose | Saline or DTX IV or NIR | Single dose (after 1 and 24 h) |
| CHD NP | ||||||||||||
| Monk et al. [40] | Low fat | ~10–16/30–48 | Experimental | Treatment | NR | NR | NR | Lard and corn oils | Oral | 16 | NA | NA |
| High fat | 18.24 | Lard, corn, menhaden, and fish oils | ||||||||||
| Luo et al. [19] | Control | 3–4/7 | Chow | Prevention and treatment | NR | NR | NR | Corn oil | Gavage | ~11 | NA | NA |
| ω3 PUFA + AA | 90 µl | NR | ||||||||||
| Ion, et al. [41] | Control/High ω3 (mothers) | 23/46 | AIN-76A | Prevention and treatment | 50 or 40 | 0 or 20 | NR | Corn and canola oils or fish oil | Oral | 2 | NA | NA |
| CC/FC | 4/16 | NR | 50 | Corn and canola oil | ~17; 18.5 and 20 | |||||||
| CF/FF | 40 | 20 | Fish oil | |||||||||
| Guo et al. [42] | Control Negative (CNB) | 8/64 | Standard Se free | Treatment | NA | NA | NA | NA | NA | NA | Saline | ~4 (1×/every 4 days) |
| Control Positive (TB) | ||||||||||||
| TB-NS | NA | |||||||||||
| TB-TAX or Adr or Ava | TAX–IP | |||||||||||
| Adr–IV | ||||||||||||
| Ava–IP | ||||||||||||
| TB-NS TAX or Adr or Ava | 3.52 µg | NA | NA | Nutrition supplementation (NS) with Selenium (Se) | Gavage | ~4 (2×/day) | TAX–IP | |||||
| Adr–IV | ||||||||||||
| Ava–IP | ||||||||||||
| Garay et al. [43] | CO-diet (ω6/ω3) | 8–10/24–30 | AIN-93 | Prevention | 0.396 | 32.31 | 19,14 | Corn oil | Oral | ~13 | NA | NA |
| SO-diet (ω6) | 0.014 | 45.48 | 8.56 | Safflower oil | ||||||||
| ChO-diet (ω3) | 37.8 | 12.80 | 4.03 | Chia oil | ||||||||
| Abbas et al. [44] | Ca (Phase I) | 15/30 | AIN-93 | Epigenetic modulation and prevention | NR | NR | NR | Canola oil or Corn oil (mother diet) | Oral | 14d | NA | NA |
| Co (Phase I) | ||||||||||||
| Ca-Co (Phase II) | 100/200 | 50d | ||||||||||
| Co-Co pups (Phase II) | ||||||||||||
| Ca-Co (Phase III) | 50/200 | Corn oil | 42d | |||||||||
| Co-Co (Phase III) | ||||||||||||
| Newell et al. [45] | OLA/LNA (Control) | 6/12 | AIN-76 | Treatment | 0 | 31.6 | 95.8 | Oleic and linoleic acid | Oral | 5 | Saline or DOX | 4 (2×/w) |
| DHA | 6.8 | 27.8 | 90.4 | DHAsco | ||||||||
| Liu et al. [46] | LFD (WT × A-FABP−/−) | 9/36 | Diet 10% fat | Treatment | NR | NR | NR | Soybean oil | Oral | 5 months | NA | NA |
| HFD-C | Diet 45% fat | 0 | Cocoa butter, soybean oils | |||||||||
| HFD-F (WT × A-FABP−/−) | 175.5 | Fish and soybean oils | ||||||||||
| Hillyer et al. [47] | ω6 PUFA, control | 11/46 | Standard | Prevention and treatment | 0.49 | 72.09 | 15.44 | Safflower oil | Oral | 20 | NA | NA |
| ω3 PUFA, control | 10/46 | 9.99 | 52.99 | 14.1 | Menhaden and safflower oils | |||||||
| ω3 PUFA | 9/46 | 17.44 | 53.90 | 16.76 | Flaxseed and safflower oils | |||||||
| MUFA | 6/46 | 0.66 | 11.15 | 64.72 | Olive oil | |||||||
| SFA | 10/46 | 2.1 | 22.46 | 35.78 | Lard | |||||||
| Goupille, et al. [48] | Control | 14/56 | Standard | Prevention and treatment | 0 | NR | NR | Peanut and rapeseed oils | Oral | 9 | DTX–IP | 6 (1×/w) |
| ω3 LCPUFA | Enriched diet | 35 | NR | NR | Peanut, rapeseed and fish oils | |||||||
| Torres-Adorno et al. [49] | Control (SUM149PT × BCX010) | 9–10/27–30 | AIN-76A | Treatment | NA | NA | NA | NA | Oral | ~6 | Dasatinib–IP | ~6 (6×/w) |
| EPA (SUM149PT × BCX010) | 0.4 or 0.8 | Fish oil | ||||||||||
| Newell et al. [50] | Control | 8/32 | NR | Prevention and treatment | NA | 52.46 | 69.8 | NR | Oral | 7 | Saline or DTX | 6 (2×/w) |
| DHA | 7.8 | 43.06 | 67.2 | DHAsco | ||||||||
| Newell et al. [51] | Control | 6/24 | AIN-76 | Prevention and treatment | NR | 30.98 | 95.8 | Sunflower, canola, olive, and ARAsco oils | Oral | 7 | Saline or DOX | 6 (2×/w) |
| DHA | 6.76 | 27.64 | 90.4 | DHAsco | ||||||||
| Li et al. [52] | Control | 24/43 | AIN-93G | Prevention | NR | NR | NR | Soybean oil | Oral | 3–7 | NA | NA |
| Fat-1 | 19/43 | ω3 PUFA endogenous from soybean oil | ||||||||||
| Fernando et al. [53] | Control (4T1 × MDA-MB-231) | NR | Standard | Treatment | NA | NA | NA | NA | IP | 5× in 9 days and 20× in 39 days | Saline | 5× in 9 days and 20× in 39 days |
| PZ-DHA (4T1 × MDA-MB-231) | NR | PZ | ||||||||||
| Białek et al. [54] | SAF (control) | 8/46 | Standard or Labofeed H | Prevention and treatment | 0.95 | 75.3 | 130.24 | Safflower oil | Gavage | 21 | NA | NA |
| SAF-plus | 14/46 | |||||||||||
| CLA (control) | 7/46 | NA | 42.7 | 37.187 | Bio conjugated linoleic acid | |||||||
| CLA-plus | 17/46 | |||||||||||
| Zhu et al. [55] | Cell induction model–Control, EPA, RP and EPA-RP | 10/40 | Standard | Treatment | 30 | NA | NA | EPA commercial product | Oral | 2 | Saline or Rapamycin | 2 |
| Transgenic model–Control, EPA, RP, EPA-RP and EPA-RP-NAC | 6/30 | EPA commercial product or NAC | 4 | 4 | ||||||||
| Liu et al. [56] | Control (ω6) | ~8/32 | AIN-93G | Prevention and treatment | 0.422 | 15.796 | 3.586 | Safflower oil | Oral | 6 and 20 | NA | NA |
| FS (ω3) | 1.14 | 3.66963 | 1.056 | Flaxseed oil | ||||||||
| FS-SF (ω3e6) | 12.694 | 3.63 | 3.3 | Flaxseed and safflower oil | ||||||||
| Menh-SF (ω3 > 6) | 0.43164 | 3.7686 | 0.9504 | Menhaden fish and safflower oil | ||||||||
| Khadge et al. [57] | ω6 (control) | 20/40 | Lieber-DeCarli (control diet) | Prevention | NR | NR | NR | Corn, olive and safflower oils | Oral | 10–16 | NA | NA |
| ω3 | Lieber-DeCarli modified | Corn, olive, fish and safflower oils | ||||||||||
| Jiao et al. [58] | ω3 | 5/20 | NR | Treatment | NR | NR | NR | Fish oil | Oral | Throughout | Vehicle or SFN–IP | Every 2 days/9 days |
| ω6 | Corn oil | |||||||||||
| Zhu et al. [59] | Control | 10/40 | NR | Treatment | 30 | NA | NA | EPA commercial product | NR | 2 | Vehicle or RA | 2 |
| RA (retinoic acid) | ||||||||||||
| EPA | ||||||||||||
| RA and EPA | ||||||||||||
| Vara-Messler et al. [60] | CO-diet (ω6) | 20/37 | Standard | Treatment | 0.8686 | 22.7943 | 5.7749 | Corn oil | Oral | ~6 | NA | NA |
| ChO-diet (ω3) | 17/37 | 27.1932 | 10.1695 | 0.9847 | Chia seed oil | |||||||
| Jiao et al. [61] | Less aggressive model | 5/40 | Standard | Prevention and treatment | NR | NR | NR | Corn oil | Oral | Throughout | Vehicle or DSF–IP | 5 days |
| Fish oil | ||||||||||||
| More aggressive model | Corn oil | |||||||||||
| Fish oil | ||||||||||||
| Dyari et al. [62] | Control | ~4–8/24 | NR | Treatment | NR | NR | NR | Corn oil | NA | 6 (1×/day) | Vehicle or AUDA–IP | 6 (1×/day) |
| AUDA (vehicle) | IP | |||||||||||
| C20E 0.05 | ω3 endogenous from corn oil | IP | ||||||||||
| C20E 0.5 | ||||||||||||
| Ref | Groups | ω-3 PUFA | ω-6 PUFA | ω-9 MUFA | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | ALA C18:3 | SDA C18:4 | ETA C20:3 | EPA C20:5 | DPA C22:5 | DHA C22:6 | Total | LA C18:2 | GLA C18:3 | EDA 20:2 | DGLA C20:3 | ARA C20:4 | DDA C22:2 | AdA C22:4 | Total | OA C18:1 | EA C22:1 | NerA C24:1 | ||
| Newell et al. [17] | Control | 4.8 g/kg | 4.8 g/kg | 42.8 g/kg | 42 g/kg | 0.84 g/kg | 72.8 g/kg | 72.8 g/kg | ||||||||||||
| High DHA | 14.4 g/kg | 6.8 g/kg | 7.6 g/kg | 31.2 g/kg | 30.4 g/kg | 0.8 g/kg | 77.4 g/kg | 77.4 g/kg | ||||||||||||
| Low DHA | 14.4 g/kg | 10.4 g/kg | 0.8 g/kg | 3.2 g/kg | 27.0 g/kg | 26.2 g/kg | 81.6 g/kg | 81.6 g/kg | ||||||||||||
| Li et al. [18] | DHA/FA/LN | 30 μL/mL | 30 μL/mL | NA | NA | |||||||||||||||
| Wang et al. [39] | CHD NP | NA | + | NA | NA | |||||||||||||||
| Monk et al. [40] | High fat | 18.24 g/kg | + | + | + | NR | + | NR | + | |||||||||||
| Luo et al. [19] | ω3 PUFA | 90 ul | + | + | + | NR | + | + | NR | |||||||||||
| Ion, et al. [41] | Control ω3 | 50 g/kg | + | + | NR | |||||||||||||||
| (mothers) | 40 g/kg | + | + | + | 20 g/kg | + | + | NR | ||||||||||||
| CC/FC | 50 g/kg | + | + | NR | ||||||||||||||||
| CF/FF | 40 g/kg | + | + | + | 20 g/kg | + | + | NR | ||||||||||||
| Guo et al. [42] | Control | NA | NA | NA | NA | NA | ||||||||||||||
| TB-NS | 3.52 ug | 2.04 ug | 1.48 ug | NA | NA | |||||||||||||||
| TB-NS drugs | ||||||||||||||||||||
| Garay et al. [43] | ChO-diet (ω3) | 37.8 g/kg | 37.8 g/kg | 12.8 g/kg | 12.8 g/kg | + | 4.03 g/kg | 4.03 g/kg | ||||||||||||
| CO-diet (ω6/ω3) | 0.4 g/kg | 0.4 g/kg | 32.3 g/kg | 32.3 g/kg | + | 19.1 g/kg | 19.1 g/kg | |||||||||||||
| SO-diet (ω6) | 0.014 g/kg | 0.014 g/kg | 45.5 g/kg | 45.5 g/kg | + | 8.56 g/kg | 8.56 g/kg | |||||||||||||
| Abbas et al. [44] | Ca(Phase I) | NR | + | NR | + | NR | + | |||||||||||||
| Co(Phase I) | NR | + | NR | + | NR | + | ||||||||||||||
| Ca/Co(Phase II) | NR | + | NR | + | NR | + | ||||||||||||||
| Co/Co(Phase II) | NR | + | NR | + | NR | + | ||||||||||||||
| Ca/Co(Phase III) | NR | + | NR | + | NR | + | ||||||||||||||
| Co/Co(Phase III) | NR | + | NR | + | NR | + | ||||||||||||||
| Newell M et al. [45] | Control | 0 | 31.6 g/kg | 27.8 g/kg | 2.26 g/kg | 1.0 g/kg | 95.8 g/kg | 95.8 g/kg | ||||||||||||
| DHA | 6.8 g/kg | 1.2 g/kg | 5.6 g/kg | 27.8 g/kg | 26.2 g/kg | 0.6 g/kg | 1.0 g/kg | 90.4 g/kg | 90.4 g/kg | |||||||||||
| Liu et al. [46] | HFD-C | 0 g/kg | NR | NR | ||||||||||||||||
| HFD-F | 175.5 g/kg | + | + | NR | NR | |||||||||||||||
| Hillyer et al. [47] | ω6 PUFA, control | 0.49 g/kg | 0.31 g/kg | 0.13 g/kg | 0 g/kg | 0.05 g/kg | 72.1 g/kg | 71.93 g/kg | 0.11 g/kg | 0.05 g/kg | 0.11 g/kg | 15.4 g/kg | 15.25 g/kg | 0.02 g/kg | 0.17 g/kg | |||||
| ω3 PUFA, control | 9.99 g/kg | 0.86 g/kg | 0.9 g/kg | 0.2 g/kg | 4.20 g/kg | 0.75 g/kg | 3.08 g/kg | 53 g/kg | 51.7 g/kg | 0.16 g/kg | 0.12 g/kg | 0.10 g/kg | 0.49 g/kg | 0.27 g/kg | 0.14 g/kg | 14.1 g/kg | 13.78 g/kg | 0.09 g/kg | 0.2 g/kg | |
| ω3 PUFA | 17.44 g/kg | 17.3 g/kg | 0.1 g/kg | 53.9 g/kg | 53.81 g/kg | 0.09 g/kg | 16.8 g/kg | 16.59 g/kg | 0.02 g/kg | 0.2 g/kg | ||||||||||
| MUFA | 0.66 g/kg | 0.66 g/kg | 11.2 g/kg | 11.2 g/kg | 64.7 g/kg | 64.50 g/kg | 0.03 g/kg | 0.2 g/kg | ||||||||||||
| SFA | 2.1 g/kg | 1.56 g/kg | 0.2 g/kg | 0.3 g/kg | 0.12g/kg | 22.5 g/kg | 20.85 g/kg | 0.11 g/kg | 0.84 g/kg | 0.17 g/kg | 0.37 g/kg | 0.12 g/kg | 35.8 g/kg | 35.76 g/kg | 0.02 g/kg | |||||
| Goupille et al. [48] | Control | 0 g/kg | NR | + | NR | + | ||||||||||||||
| ω3 LCPUFA | 35 g/kg | 10 g/kg | 25 g/kg | NR | + | NR | + | |||||||||||||
| Torres-Adorno et al. [49] | EPA | 0.4 g/kg | 0.4 g/kg | NA | NA | |||||||||||||||
| EPA | 0.8 g/kg | 0.8 g/Kg | NA | NA | ||||||||||||||||
| Newell et al. [51] | Control | NA | 52.5 g/kg | 47.8 g/kg | 3.8 g/kg | 0.86 g/kg | 69.8 g/kg | 69.8 g/kg | ||||||||||||
| DHA | 7.8 g/kg | 7.8 g/kg | 43.1 g/kg | 37 g/kg | 5.2 g/kg | 0.86 g/kg | 67.2 g/kg | 67.2 g/kg | ||||||||||||
| Newell et al. [50] | Control | NR | 31 g/kg | 27.8 g/kg | 2.26 g/kg | 0.92 g/kg | 95.8 g/kg | 95.8 g/kg | ||||||||||||
| DHA | 6.76 g/kg | 1.12 g/kg | 5.64 g/kg | 27.6 g/kg | 26.2 g/kg | 0.52 g/kg | 0.92 g/kg | 90.4 g/kg | 90.4 g/kg | |||||||||||
| Li et al. [52] | Control | NR | + | NR | + | |||||||||||||||
| Fat-1 | NR | + | NR | + | NR | + | ||||||||||||||
| Fernando et al. [53] | Control | NR | NR | NR | ||||||||||||||||
| PZ-DHA | NA | + | NA | NA | ||||||||||||||||
| Białek et al. [54] | SAF (control) | 0.95 g/kg | 0.95 g/kg | 75.3 g/kg | 75.3 g/kg | 130.2 g/kg | 130 g/kg | 0.24 g/kg | ||||||||||||
| SAF-plus | 0.95 g/kg | 0.95 g/kg | 75.3 g/kg | 75.3 g/kg | 130.2 g/kg | 130 g/kg | 0.24 g/kg | |||||||||||||
| CLA (control) | 42.7 g/kg | 42.7 g/kg | 37.2 g/kg | 37 g/kg | 0.2 g/kg | |||||||||||||||
| CLA-plus | 42.7 g/kg | 42.7 g/kg | 37.2 g/kg | 37 g/kg | 0.2 g/kg | |||||||||||||||
| Zhu et al. [55] | Cell induction model- EPA | 30 g/kg | 30g/kg | NA | NA | |||||||||||||||
| Transgenic model- EPA | 30 g/kg | 30g/kg | NA | NA | ||||||||||||||||
| Liu et al. [56] | Control (ω6) | 0.422 g/kg | 0.04 g/kg | 0.02 g/kg | - | - | - | - | 15.8 g/kg | 15.73 g/kg | 0.07 g/kg | 3.59 g/kg | 3.59 g/kg | |||||||
| FS (ω3) | 1.14 g/kg | 1.14 g/kg | - | - | - | - | - | 3.67 g/kg | 3.66 g/kg | 0.01 g/kg | 1.06 g/kg | 1.06 g/kg | ||||||||
| FS-SF (ω3:w6) | 12.7 g/kg | 12.7 g/kg | - | 0.02 g/kg | - | - | - | 3.63 g/kg | 3.63 g/kg | - | 3.3 g/kg | 3.3 g/kg | ||||||||
| Menh-SF (ω3>w6) | 0.43 g/kg | 0.04 g/kg | 0.05 g/kg | 0.01 g/kg | 0.26 g/kg | 0.05 g/kg | 0.02 g/kg | 3.8 g/kg | 3.7 g/kg | 0.01 g/kg | 0.01 g/kg | 0.01 g/kg | 0.03 g/kg | 0.01 g/kg | 0.01 g/kg | 0.95 g/kg | 0.94 g/kg | 0.01 g/kg | ||
| Khadge et al. [57] | ω6 (control) | NR | + | NR | + | NR | + | |||||||||||||
| ω3 | NR | + | NR | + | NR | + | ||||||||||||||
| Jiao et al. [58] | ω-6 | NR | + | NR | + | NR | + | |||||||||||||
| ω-3 | NR | + | + | NR | NR | |||||||||||||||
| Zhu et al. [59] | EPA | 30 g/kg | + | 30 g/kg | + | NA | NA | |||||||||||||
| Vara-Messler et al. [60] | CO-diet (ω6) | 27.2 g/kg | 27.2 g/kg | 10.2 g/kg | 10.2 g/kg | 0.98 g/kg | 0.98 g/kg | |||||||||||||
| ChO-diet (ω3) | 0.87 g/kg | 0.87 g/kg | 22.8 g/kg | 22.8 g/kg | 5.8 g/kg | 5.8 g/kg | ||||||||||||||
| Jiao et al. [61] | Less aggressive model | NR | + | NR | + | NR | + | |||||||||||||
| More aggressive model | NR | + | NR | NR | ||||||||||||||||
| Dyari et al. [62] | C20E | NR | + | NR | + | NR | + | |||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Theinel, M.H.; Nucci, M.P.; Alves, A.H.; Dias, O.F.M.; Mamani, J.B.; Garrigós, M.M.; Oliveira, F.A.; Rego, G.N.A.; Valle, N.M.E.; Cianciarullo, G.; et al. The Effects of Omega-3 Polyunsaturated Fatty Acids on Breast Cancer as a Preventive Measure or as an Adjunct to Conventional Treatments. Nutrients 2023, 15, 1310. https://doi.org/10.3390/nu15061310
Theinel MH, Nucci MP, Alves AH, Dias OFM, Mamani JB, Garrigós MM, Oliveira FA, Rego GNA, Valle NME, Cianciarullo G, et al. The Effects of Omega-3 Polyunsaturated Fatty Acids on Breast Cancer as a Preventive Measure or as an Adjunct to Conventional Treatments. Nutrients. 2023; 15(6):1310. https://doi.org/10.3390/nu15061310
Chicago/Turabian StyleTheinel, Matheus H., Mariana P. Nucci, Arielly H. Alves, Olivia F. M. Dias, Javier B. Mamani, Murilo M. Garrigós, Fernando A. Oliveira, Gabriel N. A. Rego, Nicole M. E. Valle, Gabriela Cianciarullo, and et al. 2023. "The Effects of Omega-3 Polyunsaturated Fatty Acids on Breast Cancer as a Preventive Measure or as an Adjunct to Conventional Treatments" Nutrients 15, no. 6: 1310. https://doi.org/10.3390/nu15061310
APA StyleTheinel, M. H., Nucci, M. P., Alves, A. H., Dias, O. F. M., Mamani, J. B., Garrigós, M. M., Oliveira, F. A., Rego, G. N. A., Valle, N. M. E., Cianciarullo, G., & Gamarra, L. F. (2023). The Effects of Omega-3 Polyunsaturated Fatty Acids on Breast Cancer as a Preventive Measure or as an Adjunct to Conventional Treatments. Nutrients, 15(6), 1310. https://doi.org/10.3390/nu15061310