Metabolites Identified during Varied Doses of Aspergillus Species in Zea mays Grains, and Their Correlation with Aflatoxin Levels

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Crop Development
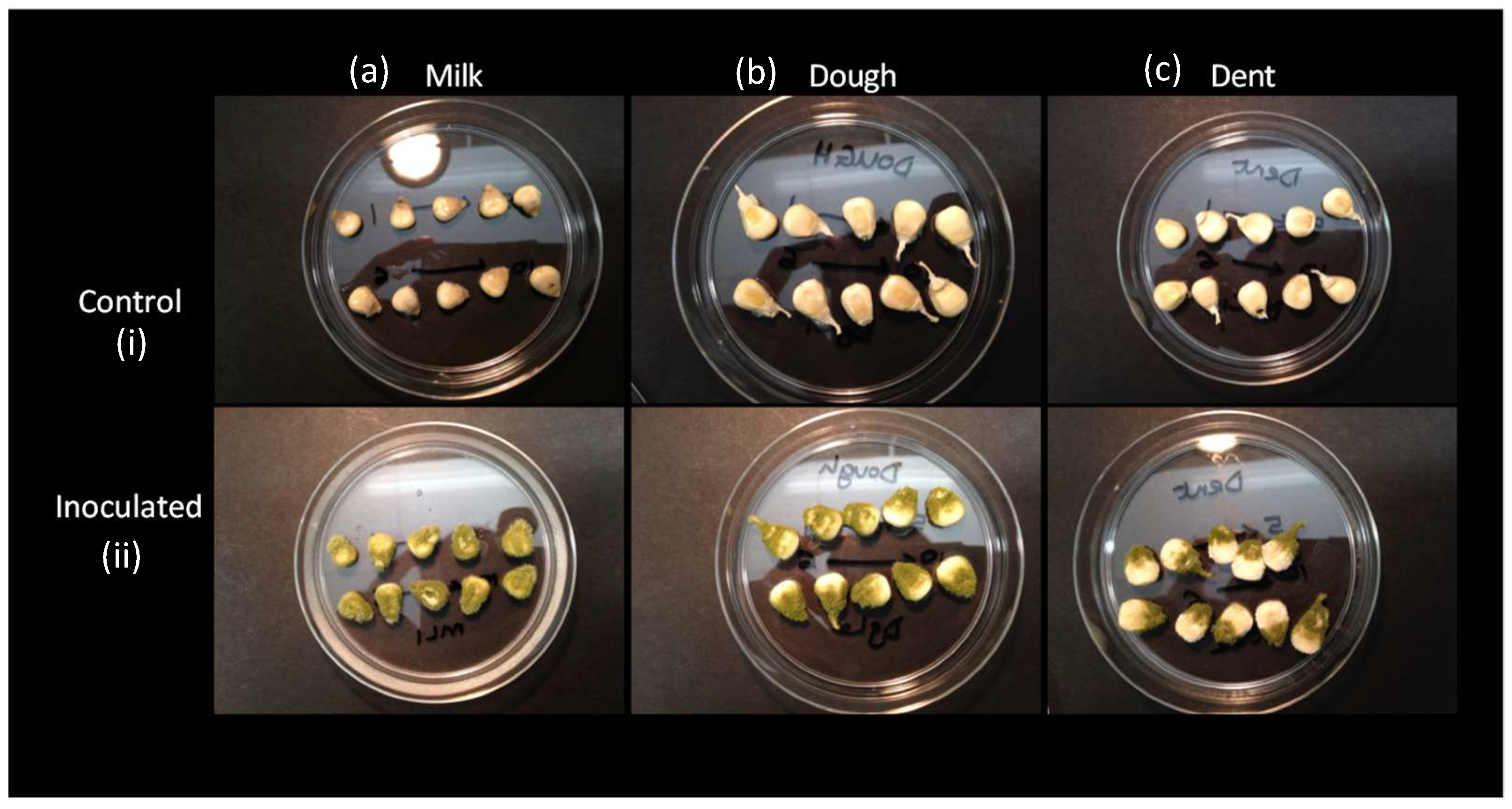
2.2. Grain Colonisation with Fungal Spores
2.3. Aflatoxin Levels
2.4. Polar Metabolites Identification
2.4.1. Stepwise Discriminant Analysis
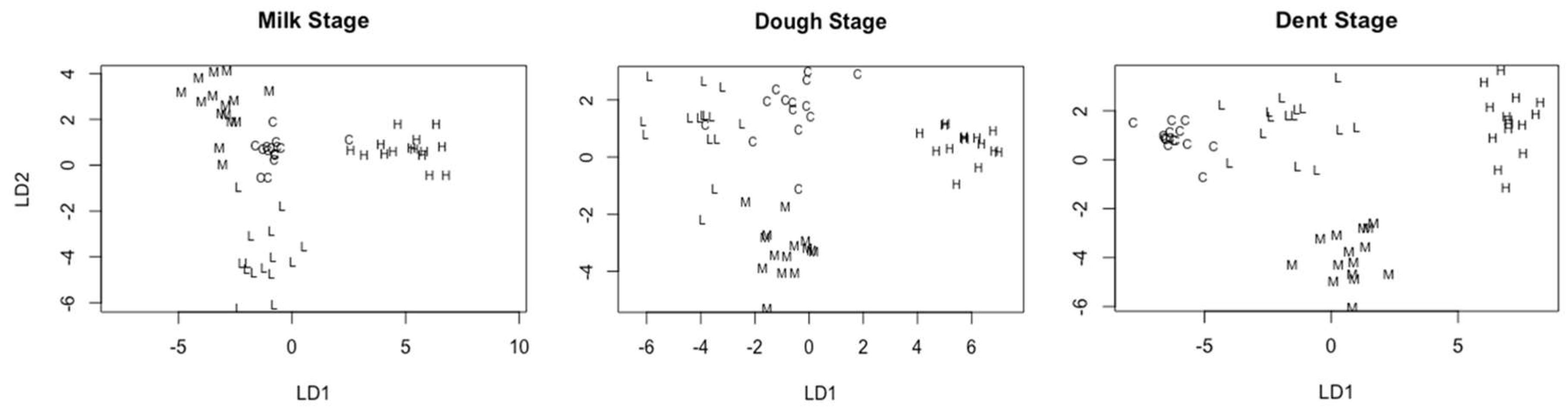
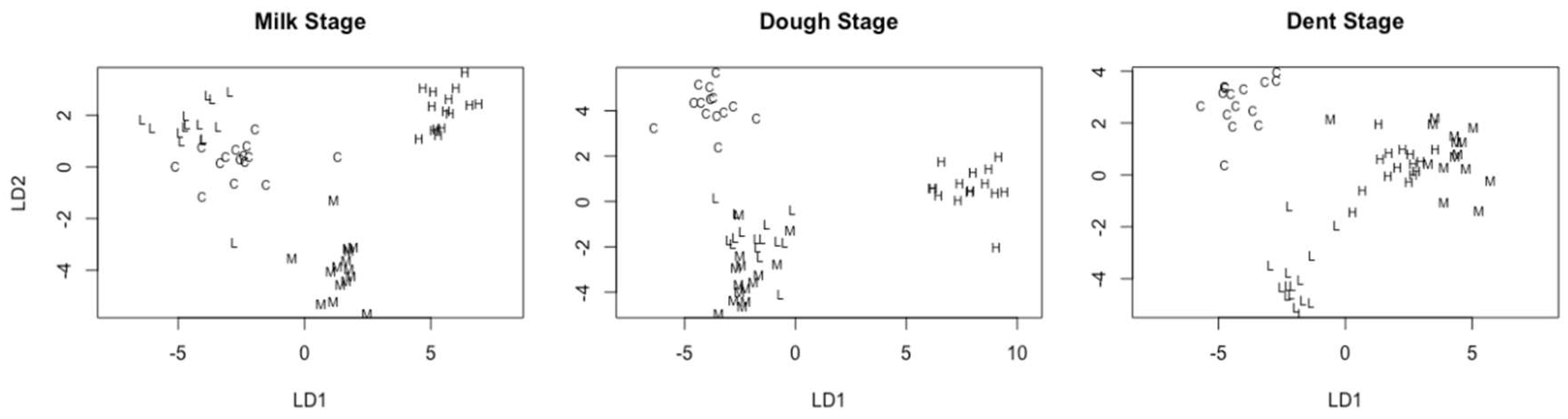
2.4.2. Linear Discriminant Analysis
3. Discussion
3.1. Sugars
3.2. Sugar Alcohols
3.3. Polar Acids
3.3.1. Amino Acids
3.3.2. Other Metabolites
3.3.3. Fatty Acid Methyl Esters (FAME) Identification
4. Conclusions
5. Materials and Methods
5.1. Crop Development
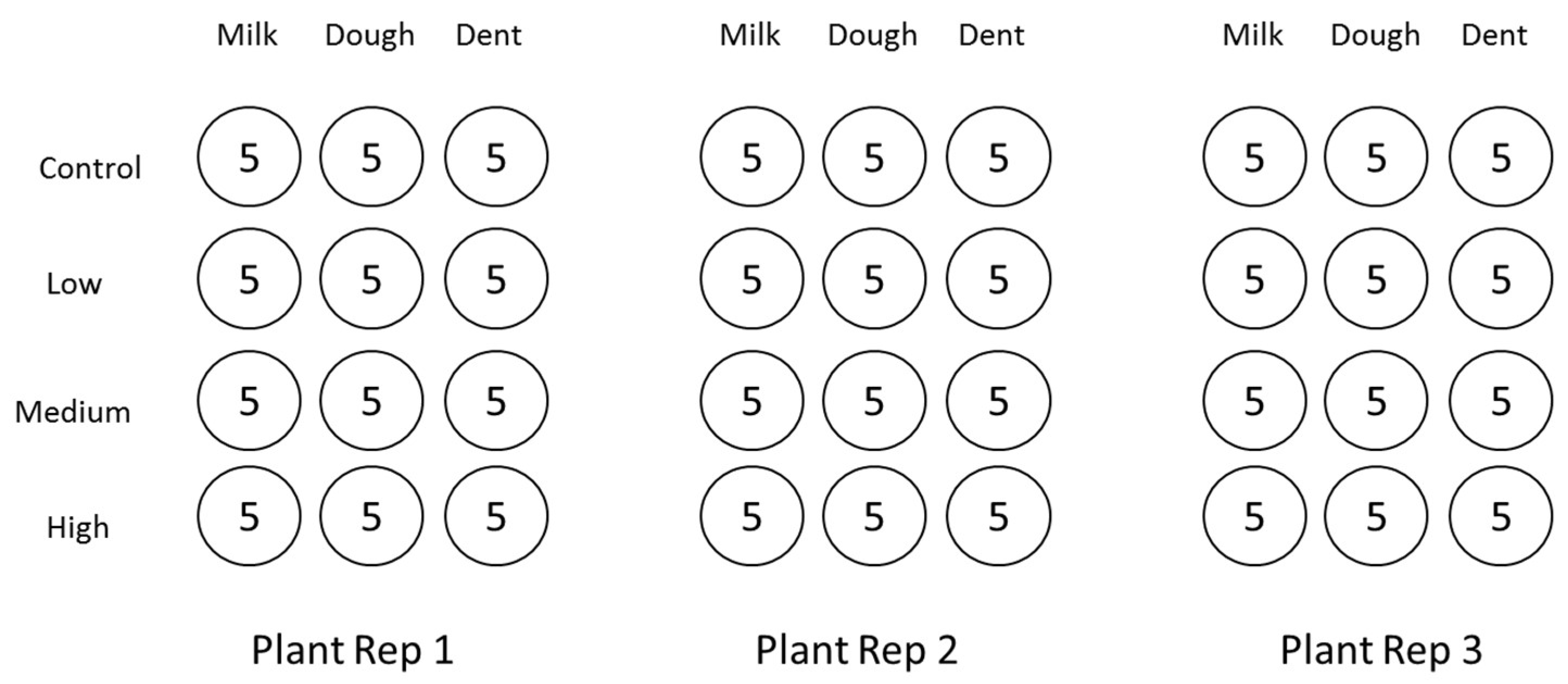
5.2. Grain Colonisation with Fungal Spores
5.3. Chemical Analyses
5.3.1. Sample Preparation and Extraction for Metabolomics Analysis
5.3.2. Polar Phase GC-MS Metabolomics Analysis
5.3.3. Non-Polar Phase GC-MS Fatty Acid Methyl Ester (FAME) Metabolomics Analysis
5.3.4. GC-MS Data Processing
5.3.5. Extraction and Analysis of Aflatoxins
5.4. Statistical Analyses of Aflatoxin and Metabolite Levels
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Asiki, G.; Seeley, J.; Srey, C.; Baisley, K.; Lightfoot, T.; Archileo, K.; Agol, D.; Abaasa, A.; Wakeham, K.; Routledge, M.N.; et al. A pilot study to evaluate aflatoxin exposure in a rural Ugandan population. Trop. Med. Int. Heal. 2014, 19, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Akande, A.; Mutegi, C.; Atehnkeng, J.; Kaptoge, L.; Senghor, A.L.; Adhikari, B.N.; Cotty, P.J. Biological control of aflatoxins in Africa: Current status and potential challenges in the face of climate change. World Mycotoxin J. 2016, 9, 771–789. [Google Scholar] [CrossRef]
- Mmongoyo, J.A.; Wu, F.; Linz, J.E.; Nair, M.G.; Mugula, J.K.; Tempelman, R.J.; Strasburg, G.M. Aflatoxin levels in sunflower seeds and cakes collected from micro- and small-scale sunflower oil processors in Tanzania. PLoS ONE 2017, 12, e0175801. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.E.; Groopman, J.D. Aflatoxin and liver cancer. Baillieres. Clin. Gastroenterol. 1999, 13, 545–555. [Google Scholar] [CrossRef]
- Kpodo, K.A.; Bankole, S.A. Mycotoxin contamination in foods in West and Central Africa. In Mycotoxins. Detection Methods, Management, Public Heallth and Agricultural Trade; Leslie, J.F., Bandyopadhyay, R., Visconti, A., Eds.; CABI International: Wallingford, UK, 2008; pp. 103–116. [Google Scholar]
- Egal, S.; Hounsa, A.; Gong, Y.Y.; Turner, P.C.; Wild, C.P.; Hall, A.J.; Hell, K.; Cardwell, K.F. Dietary exposure to aflatoxin from maize and groundnut in young children from Benin and Togo, West Africa. Int. J. Food Microbiol. 2005, 104, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Castelino, J.M.; Routledge, M.N.; Wilson, S.; Dunne, D.W.; Mwatha, J.K.; Gachuhi, K.; Wild, C.P.; Gong, Y.Y. Aflatoxin exposure is inversely associated with IGF1 and IGFBP3 levels in vitro and in Kenyan schoolchildren. Mol. Nutr. Food Res. 2015, 59, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.Y.; Cardwell, K.; Hounsa, A.; Egal, S.; Turner, P.C.; Hall, A.J.; Wild, C.P. Dietary aflatoxin exposure and impaired growth in young children from Benin and Togo: Cross sectional study. BMJ 2002, 325, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Price, M.S.; Conners, S.B.; Tachdjian, S.; Kelly, R.M.; Payne, G.A. Aflatoxin conducive and non-conducive growth conditions reveal new gene associations with aflatoxin production. Fungal Genet. Biol. 2005, 42, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.R.; Yu, J.; Abbas, H.K.; Scheffler, B.E.; Kim, H.S.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E.; Yu, J.; Abbas, H.K.; et al. Aflatoxin formation and gene expression in response to carbon source media shift in Aspergillus parasiticus. Food Addit. Contam. 2007, 24, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.R.; Yu, J.; Bland, J.M.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E. Amino acid supplementation reveals differential regulation of aflatoxin biosynthesis in Aspergillus flavus NRRL 3357 and Aspergillus parasiticus SRRC 143. Appl. Microbiol. Biotechnol. 2007, 74, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Okoth, S.; Nyongesa, B.; Joutsjoki, V.; Korhonen, H.; Ayugi, V.; Kang’ethe, E. Sclerotia formation and toxin production in large sclerotial Aspergillus flavus isolates from Kenya. Adv. Microbiol. 2016, 6, 47–56. [Google Scholar] [CrossRef]
- Roze, L.V.; Chanda, A.; Laivenieks, M.; Beaudry, R.M.; Artymovich, K.A.; Koptina, A.V.; Awad, D.W.; Valeeva, D.; Jones, A.D.; Linz, J.E. Volatile profiling reveals intracellular metabolic changes in Aspergillus parasiticus: VeA regulates branched chain amino acid and ethanol metabolism. BMC Biochem. 2010, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Wahl, R.; Horst, R.J.; Voll, L.M.; Poree, F.; Stitt, M.; Sonnewald, U.; Kahmann, R. Reprogramming a maize plant: Transcriptional and metabolic changes induced by the fungal biotroph Ustilago maydis. Plant J. 2008, 56, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Marti, G.; Erb, M.; Boccard, J.; Glauser, G.; Doyen, G.R.; Villard, N.; Robert, C.A.M.; Turlings, T.C.; Rudaz, S.; Wolfender, J.L. Metabolomics reveals herbivore-induced metabolites of resistance and susceptibility in maize leaves and roots. Plant Cell Environ. 2013, 36, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Laparre, J.; Malbreil, M.; Letisse, F.; Portais, J.C.; Roux, C.; Becard, G.; Puech-Pages, V. Combining metabolomics and gene expression analysis reveals that propionyl- and butyryl-carnitines are involved in late stages of arbuscular mycorrhizal symbiosis. Mol. Plant 2014, 7, 554–566. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Polle, A. What the transcriptome does not tell—Proteomics and metabolomics are closer to the plants’ patho-phenotype. Curr. Opin. Plant Biol. 2015, 26, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, D.; Rajasekaran, K.; Payne, G.A.; Brown, R.L.; Yu, J.; Cleveland, T.E. The “omics” tools: Genomics, proteomics, metabolomics and their potential for solving the aflatoxin contamination problem. World Mycotoxin J. 2008, 1, 3–12. [Google Scholar] [CrossRef]
- Sun, C.; Gao, X.; Fu, J.; Zhou, J.; Wu, X. Metabolic response of maize (Zea mays L.) plants to combined drought and salt stress. Plant Soil 2015, 388, 99–117. [Google Scholar] [CrossRef]
- Gavaghan, C.L.; Li, J.V.; Hadfield, S.; Hole, S.; Nicholson, J.K.; Wilson, I.D.; Howe, P.W.; Stanley, P.D.; Holmes, E. Application of NMR-based metabolomics to the investigation of salt stress in maize (Zea mays). Phytochem. Anal. 2011, 22, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.; Cheng, F.; Hu, C.; Quan, S.; Lin, H.; Wang, J.; Chen, G.; Zhao, X.; Alexander, D.; Guo, L.; et al. Metabolic map of mature maize kernels. Metabolomics 2014, 10, 775–787. [Google Scholar] [CrossRef]
- Sun, C.X.; Li, M.Q.; Gao, X.; Liu, L.N.; Wu, X.F.; Zhou, J.H. Metabolic response of maize plants to multi-factorial abiotic stresses. Plant Biol. 2015, 18, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Malysheva, S.V.; Arroyo-Manzanares, N.; Cary, J.W.; Ehrlich, K.C.; Vanden Bussche, J.; Vanhaecke, L.; Bhatnagar, D.; Di Mavungu, J.D.; De Saeger, S. Identification of novel metabolites from Aspergillus flavus by high resolution and multiple stage mass spectrometry. Food Addit. Contam. 2014, 31, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Forseth, R.; Amaike, S.; Schwenk, D.; Affeldt, K.J.; Hoffmeister, D.; Schroeder, F.; Keller, N.P. Homologous NRPS-like gene clusters mediate redundant small molecule biosynthesis in Aspergillus flavus. Angew. Commun. 2013, 52, 1590–1594. [Google Scholar] [CrossRef] [PubMed]
- Brodhagen, M.; Keller, N.P. Signalling pathways connecting mycotoxin production and sporulation. Mol. Plant Pathol. 2006, 7, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Barnes, L.W. Aflatoxin Development in Preharvest Corn. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 1983. [Google Scholar]
- Dolezal, A.L.; Shu, X.; OBrian, G.R.; Nielsen, D.M.; Woloshuk, C.P.; Boston, R.S.; Payne, G.A. Aspergillus flavus infection induces transcriptional and physical changes in developing maize kernels. Front. Microbiol. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.X.; Allen, L.H.; Vaughan, M.M.; Hauser, B.A.; Boote, K.J. Solar ultraviolet radiation exclusion increases soybean internode lengths and plant height. Agric. For. Meteorol. 2014, 184, 170–178. [Google Scholar] [CrossRef]
- Temminghoff, E.J.; Van der Zee, S.E.; De Haan, F.A. Copper mobility in a copper-contaminated sandy soil as affected by pH and solid and dissolved organic matter. Environ. Sci. Technol. 1997, 31, 1109–1115. [Google Scholar] [CrossRef]
- Gupta, S.K.; Maggon, K.K.; Venkitasubramanian, T.A. Regulation of aflatoxin biosynthesis. 2 Comparative study of tricarboxylic acid cycle in aflatoxigenic and non-aflatoxigenic strains of Aspergillus flavus. Microbios 1977, 19, 7–15. [Google Scholar] [PubMed]
- Cleveland, T.E.; Lax, A.R.; Lee, L.S.; Bhatnagar, D. Appearance of enzyme activities catalyzing conversion of sterigmatocystin to aflatoxin B1 in late-growth-phase Aspergillus parasiticus cultures. Appl. Environ. Microbiol. 1987, 53, 1711–1713. [Google Scholar] [PubMed]
- Mateles, R.I.; Adye, J.C. Production of aflatoxins in submerged culture. Appl. Environ. Microbiol. 1965, 13, 208–211. [Google Scholar]
- Lasram, S.; Hamdi, Z.; Chenenaoui, S.; Mliki, A.; Ghorbel, A. Comparative study of toxigenic potential of Aspergillus flavus and Aspergillus niger isolated from barley as affected by temperature, water activity and carbon source. J. Stored Prod. Res. 2016, 69, 58–64. [Google Scholar] [CrossRef]
- Manda, A.; Naidu, B.P.; Nageswararao, C.R.; Graeme, W.; Shu, F. Aflatoxins and their relationship with sugars in peanut (Arachis hypogaea L.). In The 4th International Crop Science Congress; International Crop Science Congress: Brisbane, Australia, 2004. [Google Scholar]
- Woloshuk, C.P.; Cavaletto, J.R.; Cleveland, T.E. Inducers of aflatoxin biosynthesis from colonized maize kernels are generated by an amylase activity from Aspergillus flavus. Phytopathology 1997, 87, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Gentinetta, E.; Zambello, M.; Salamini, F. Free sugars in developing maize grain. Cereal Chem. 1979, 56, 81–83. [Google Scholar]
- Severns, D.E.; Clements, M.J.; Lambert, R.J.; White, D.G. Comparison of Aspergillus ear rot and aflatoxin contamination in grain of high-oil and normal-oil corn hybrids. J. Food Prot. 2003, 66, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Loesch, A.; Hutwimmer, S.; Strasser, H. Carbon utilization pattern as a potential quality control criterion for virulence of Beauveria brongniartii. J. Invertebr. Pathol. 2010, 104, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; Hoffman, M.G.; Romer, U.; Kockenberger, W.; Elling, L.; Roitsch, T. Metabolizable and non-metabolizable sugars activate different signal transduction pathways in tomato. Plant Physiol. 2002, 128, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Mellon, J.E.; Cotty, P.J.; Dowd, M.K. Influence of lipids with and without other cottonseed reserve materials on aflatoxin B1 production by Aspergillus flavus. J. Agric. Food Chem. 2000, 48, 3611–3615. [Google Scholar] [CrossRef] [PubMed]
- Fillinger, S.; Chaveroche, M.-K.; Van Dijck, P.; De Vries, R.; Ruijter, G.; Thevelein, J.; D’enfert, C. Trehalose is required for the acquisition of tolerance to a variety of stresses in the filamentous fungus Aspergillus nidulans. Microbiology 2001, 147, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Yu, J. A novel regulator couples sporogenesis and trehalose biogenesis in Aspergillus nidulans. PLoS ONE 2007, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered Pathway Genes in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Daryaei, A.; Jones, E.E.; Alizadeh, H.; Glare, T.R.; Falloon, R.E. Biochemical characteristics of Trichoderma atroviride associated with conidium fitness for biological control. Biocontrol. Sci. Technol. 2016, 26, 189–205. [Google Scholar] [CrossRef]
- Guo, B.; Chen, Z.-Y.; Lee, R.D.; Scully, B.T. Drought stress and preharvest aflatoxin contamination in agricultural commodity: Genetics, genomics and proteomics. J. Integr. Plant Biol. 2008, 50, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.-L.; Zhang, Q.; Sun, Z.-M.; Chen, L.-H.; Liu, B.-X.; Zhang, K.-X.; Zhu, X.-M.; Shao, J.-R.; Tang, Y.-X.; Wu, Y.-M. Trehalose metabolism-related genes in maize. J. Plant Growth Regul. 2013, 33, 256–271. [Google Scholar] [CrossRef]
- Yu, J.; Woloshuk, C.P.; Bhatnagar, D.; Cleveland, T.E. Cloning and characterization of avfA and omtB genes involved in aflatoxin biosynthesis in three Aspergillus species. Gene 2000, 248, 157–167. [Google Scholar] [CrossRef]
- Chang, P.K.; Horn, B.W.; Dorner, J.W. Sequence breakpoints in the aflatoxin biosynthesis gene cluster and flanking regions in nonaflatoxigenic Aspergillus flavus isolates. Fungal Genet. Biol. 2005, 42, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Donner, M.; Atehnkeng, J.; Sikora, R.A.; Bandyopadhyay, R.; Cotty, P.J. Molecular characterization of atoxigenic strains for biological control of aflatoxins in Nigeria. Food Addit. Contam. 2010, 27, 576–590. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Schmidt-Heydt, M.; Rodriguez, A.; Parra, R.; Geisen, R.; Magan, N. Impacts of environmental stress on growth, secondary metabolite biosynthetic gene clusters and metabolite production of xerotolerant/xerophilic fungi. Curr. Genet. 2015, 61, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Nesci, A.; Etcheverry, M.; Magan, N. Osmotic and matric potential effects on growth, sugar alcohol and sugar accumulation by Aspergillus section Flavi strains from Argentina. J. Appl. Microbiol. 2004, 96, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Mellon, J.E.; Dowd, M.K.; Cotty, P.J. Time course study of substrate utilization by Aspergillus flavus in medium simulating corn (Zea mays) kernels. J. Agric. Food Chem. 2002, 50, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Kebede, H.; Abbas, H.K.; Fisher, D.K.; Bellaloui, N. Relationship between aflatoxin contamination and physiological responses of corn plants under drought and heat stress. Toxins 2012, 4, 1385–1403. [Google Scholar] [CrossRef] [PubMed]
- Baidya, S.; Duran, R.M.; Lohmar, J.M.; Harris-Coward, P.Y.; Cary, J.W.; Hong, S.Y.; Roze, L.V.; Linz, J.E.; Calvo, A.M. VeA is associated with the response to oxidative stress in the aflatoxin producer Aspergillus flavus. Eukaryot. Cell 2014, 13, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Stahl, H.G. bility of various carbon sources to induce and support aflatoxin synthesis by Aspergillus parasiticus. J. Food Saf. 1984, 6, 271–279. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Ayres, J.C. Effect of various glycolytic and TCA intermediates on aflatoxin production. J. Food Saf. 1977, 1, 19–28. [Google Scholar] [CrossRef]
- Downes, C.P.; Macphee, C.H. Myo-inositol metabolites as cellular signals. Eur. J. Biochem. 1990, 193, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, A.L. Interactions between Aspergillus flavus and the Developing Maize Kernel. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2010. [Google Scholar]
- Cherry, J.P.; Young, C.T.; Beuchat, L.R. Changes in proteins and free and total amino acids of peanuts (Arachis hypogaea) infected with Aspergillus parasiticus. Can. J. Bot. 1975, 53, 2639–2649. [Google Scholar] [CrossRef]
- Chang, P.K.; Hua, S.S.T.; Sarreal, S.B.L.; Li, R.W. Suppression of aflatoxin biosynthesis in Aspergillus flavus by 2-phenylethanol is associated with stimulated growth and decreased degradation of branched-chain amino acids. Toxins 2015, 7, 3887–3902. [Google Scholar] [CrossRef] [PubMed]
- Payne, G.A.; Hagler, W. Effect of specific amino acids on growth and aflatoxin production by Aspergillus parasiticus and Aspergillus flavus in defined media. Appl. Environ. Microbiol. 1983, 46, 805–812. [Google Scholar] [PubMed]
- Groppa, M.D.; Benavides, M.P. Polyamines and abiotic stress: Recent advances. Amino Acids 2008, 34, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Yoshiba, Y.; Kiyosue, T.; Katagiri, T.; Ueda, H.; Mizoguchi, T.; Yamaguchi-Shinozaki, K.; Wada, K.; Harada, Y.; SHINOZAKI, K. Correlation between the induction of a gene for delta 1-pyrroline-5-carboxylate synthetase and the accumulation of proline in Arabidopsis thaliana under osmotic stress. Plant J. 1995, 7, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Gao, X.; Chen, X.; Fu, J.; Zhang, Y. Metabolic and growth responses of maize to successive drought and re-watering cycles. Agric. Water Manag. 2016, 172, 62–73. [Google Scholar] [CrossRef]
- Yanagita, T. Biochemical aspects on the germination of conidiospores of Aspergillus niger. Arch. Mikrobiol. 1957, 26, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.D.; Diener, U.L.; Agnihotri, V.P. Production of aflatoxins B1 and G1 in chemically defined medium. Mycopathol. Mycol. Appl. 1967, 31, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Reddy, T.; Viswanathan, L.; Venkitasubramanian, T.A. Factors affecting aflatoxin production by Aspergillus parasiticus in a chemically defined medium. Microbiology 1979, 114, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R.P.; Solomon, P.S. Does the oxidative stress used by plants for defence provide a source of nutrients for pathogenic fungi? Trends Plant Sci. 2004, 9, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D. Primary metabolism and plant defense-fuel for the fire. Mol. Plant-Microbe Interact. 2009, 22, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.Y.; Ji, G.E. Characterization of soybean fermented by aflatoxin non-producing Aspergillus oryzae and γ-aminobutyric acid producing Lactobacillus brevis. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 703–708. [Google Scholar] [CrossRef]
- Van Buskirk, J.J.; Kirsch, W.M.; Kleyer, D.L.; Barkley, R.M.; Koch, T.H. Aminomalonic acid: Identification in Escherichia coli and atherosclerotic plaque. Proc. Natl. Acad. Sci. USA 1984, 81, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Copley, S.D.; Frank, E.; Kirsch, W.M.; Koch, T.H. Detection and possible origins of aminomalonic acid in protein hydrolysates. Anal. Biochem. 1992, 201, 152–157. [Google Scholar] [CrossRef]
- Ahmed, M.U.; Thorpe, S.R.; Baynes, J.W. Identification of N epsilon-carboxymethyllysine as a degradation product of fructoselysine in glycated protein. J. Biol. Chem. 1986, 261, 4889–4894. [Google Scholar] [PubMed]
- Harding, J.J.; Hassett, P.; Rixon, K.C.; Bron, A.J.; Harvey, D.J. Sugars including erythronic and threonic acids in human aqueous humour. Curr. Eye Res. 1999, 19, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Campbell, B.C.; Molyneux, R.; Mahoney, N.; Chan, K.L.; Yu, J.; Wilkinson, J.; Cary, J.; Bhatnagar, D.; Cleveland, T.E. Gene targets for fungal and mycotoxin control. Mycotoxin Res. 2006, 22, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Maggon, K.; Gupta, S.; Venkitasubramanian, T. Biosynthesis of aflatoxins. Bacteriol. Rev. 1977, 41, 822–855. [Google Scholar] [PubMed]
- Challenger, F.; Subramaniam, V.; Walker, T.K. The mechanism of the formation of citric and oxalic acids from sugars by Aspergillus niger. Part I. J. Chem. Soc. 1927, 200–208. [Google Scholar] [CrossRef]
- Strijbis, K.; Distel, B. Intracellular acetyl unit transport in fungal carbon metabolism. Eukaryot Cell 2010, 9, 1809–1815. [Google Scholar] [CrossRef] [PubMed]
- Varsha, K.K.; Devendra, L.; Shilpa, G.; Priya, S.; Pandey, A.; Nampoothiri, K.M. 2,4-Di-tert-butyl phenol as the antifungal, antioxidant bioactive purified from a newly isolated Lactococcus sp. Int. J. Food Microbiol. 2015, 211, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.K.; Kim, J.D.; Kim, B.S.; Kim, K.D. Root treatment with rhizobacteria antagonistic to Phytophthora blight affects anthracnose occurrence, ripening, and yield of pepper fruit in the plastic house and field. Phytopathology 2011, 101, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Dharni, S.; Sanchita; Maurya, A.; Samad, A.; Srivastava, S.K.; Sharma, A.; Patra, D.D. Purification, characterization, and in vitro activity of 2,4-Di-tert-butylphenol from Pseudomonas monteilii PsF84: Conformational and molecular docking studies. J. Agric. Food Chem. 2014, 62, 6138–6146. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Wang, J. Effect of amendments with different phenolic acids on soil microbial biomass, activity, and community diversity. Appl. Soil Ecol. 2008, 39, 172–179. [Google Scholar] [CrossRef]
- Choi, S.J.; Kim, J.K.; Kim, H.K.; Harris, K.; Kim, C.-J.; Park, G.G.; Park, C.-S.; Shin, D.-H. 2,4-Di-tert-butylphenol from sweet potato protects against oxidative stress in PC12 cells and in mice. J. Med. Food 2013, 16, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, F.G.; White, P.J.; Pollak, L.M.; Brumm, T.J. Fatty acid composition of oil from adapted, elite corn breeding materials. J. Am. Oil Chem. Soc. 1995, 72, 981–987. [Google Scholar] [CrossRef]
- White, P.J.; Pollak, L.M.; Duvick, S. Improving the fatty acid composition of corn oil by using germplasm introgression. Lipid Technol. 2007, 19, 35–38. [Google Scholar] [CrossRef]
- Canavar, O.; Kaynak, M.A. Prevention of pre-harvest aflatoxin production and the effect of different harvest times on peanut (Arachis hypogaea L.) fatty acids. Food Addit. Contam. 2013, 30, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Zain, M. Effect of olive oil on secondary metabolite and fatty acid profiles of Penicillium expansum, Aspergillus flavus, A. parasiticus and A. ochraceus. Aust. J. Basic Appl. Sci. 2009, 3, 4274–4280. [Google Scholar]
- Farag, R.S.; Youssef, A.M.; Khalil, F.A.; Taha, R.A. The lipids of various fungi grown on an artificial medium. J. Am. Oil Chem. Soc. 1981, 58, 765–768. [Google Scholar] [CrossRef]
- Kumar, A.K.; Vatsyayan, P.; Goswami, P. Production of lipid and fatty acids during growth of Aspergillus terreus on hydrocarbon substrates. Appl. Biochem. Biotechnol. 2010, 160, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Deepthi, B.V.; Poornachandra Rao, K.; Chennapa, G.; Naik, M.K.; Chandrashekara, K.T.; Sreenivasa, M.Y. Antifungal attributes of Lactobacillus plantarum MYS6 against fumonisin producing Fusarium proliferatum associated with poultry feeds. PLoS ONE 2016, 11, e0155122. [Google Scholar] [CrossRef] [PubMed]
- Vergopoulou, S.; Galanpoulou, D.; Markaki, P. Methyl jasmonate stimulates aflatoxin B1 biosynthesis by Aspergillus parasiticus. J. Agric. Food Chem. 2001, 49, 3494–3498. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, E.; Tulpule, P. Effect of free fatty acids on aflatoxin production in a synthetic medium. Food Cosmet. Toxicol. 1980, 18, 367–369. [Google Scholar] [CrossRef]
- Gao, X.; Kolomiets, M. V Host-derived lipids and oxylipins are crucial signals in modulating mycotoxin production by fungi. Toxin Rev. 2009, 28, 79–88. [Google Scholar] [CrossRef]
- Jellum, M.; Widstrom, N. Inheritance of stearic acid in germ oil of the maize kernel. J. Hered. 1983, 74, 383–384. [Google Scholar] [CrossRef]
- Alrefai, R.; Berke, T.G.; Rocheford, T.R. Quantitative trait locus analysis of fatty acid concentrations in maize. Genome 1995, 38, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.B.; Smith, J.E. Effect of exogenous lipids on growth and aflatoxin production by Aspergillus flavus. Trans. Br. Mycol. Soc. 1987, 89, 384–387. [Google Scholar] [CrossRef]
- Yan, S.; Liang, Y.; Zhang, J.; Chen, Z.; Liu, C.-M. Autoxidated linolenic acid inhibits aflatoxin biosynthesis in Aspergillus flavus via oxylipin species. Fungal Genet. Biol. 2015, 81, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Hibbard, B.E.; Bernklau, E.J.; Bjostad, L.B. Long-chain free fatty acids: Semiochemicals for host location by western corn rootworm larvae. J. Chem. Ecol. 1994, 20, 3335–3344. [Google Scholar] [CrossRef] [PubMed]
- Gornas, P.; Rudzinska, M.; Racyk, M.; Soliven, A. Lipophilic bioactive compounds in the oils recovered from cereal by-products. J. Sci. Food Agric. 2016, 96, 3256–3265. [Google Scholar] [CrossRef] [PubMed]
- Ekmay, R.D.; Papineni, S.; Herman, R.A. Crop composition, nutritional context, and suitability of animal studies. In Genetically Modified Organisms in Food: Production, Safety, Regulation and Public Health; Watson, R.R., Preedy, V.R., Eds.; Elsevier: San Diego, CA, USA, 2016. [Google Scholar]
- Monsanto Safety assessment of Roundup Ready® Corn event GA21. Safety Summaries. Available online: http://www.monsanto.com/products/documents/safety-summaries/corn_pss_ga21.pdf (accessed on 18 August 2016).
- Fanelli, C.; Fabbri, A.A. Relationship between lipids and aflatoxin biosynthesis. Mycopathologia 1989, 197, 115–120. [Google Scholar] [CrossRef]
- Lori, A.M.-H.; Richard, A.W.; Nancy, P.K. Fundamental contribution of [beta]-oxidation to polyketide mycotoxin production in planta. Mol. Plant.-Microbe Interact. 2005, 18, 783–793. [Google Scholar]
- Sheridan, K.J.; Dolan, S.K.; Doyle, S. Endogenous cross-talk of fungal metabolites. Front. Microbiol. 2015, 5, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Brodhagen, M.; Tsitsigiannis, D.I.; Hornung, E.; Goebel, C.; Feussner, I.; Keller, N.P. Reciprocal oxylipin-mediated cross-talk in the Aspergillus–seed pathosystem. Mol. Microbiol. 2008, 67, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Wee, J.; Day, D.M.; Linz, J.E. Effects of zinc chelators on aflatoxin production in Aspergillus parasiticus. Toxins 2016, 8, 171. [Google Scholar] [CrossRef] [PubMed]
- Hayes, K. Potting Media, Potting media used in UQ glasshouses. 2014.
- Nielsen, R.L. Grain Fill Stages in Corn. Corny News Network. Available online: https://www.agry.purdue.edu/ext/corn/news/timeless/GrainFill.html (accessed on 29 July 2016).
- Baker, J.M.; Hawkins, N.D.; Ward, J.L.; Lovegrove, A.; Napier, J.A.; Shewry, P.R.; Beale, M.H. A metabolomic study of substantial equivalence of field-grown genetically modified wheat. Plant Biotechnol. J. 2006, 4, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Sangster, T.; Major, H.; Plumb, R.; Wilson, A.J.; Wilson, I.D. A pragmatic and readily implemented quality control strategy for HPLC-MS and GC-MS-based metabonomic analysis. Analyst 2006, 131, 1075–1078. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.P.; Dear, G.J.; Griffin, J.L.; Haselden, J.N. An approach for the development and selection of chromatographic methods for high-throughput metabolomic screening of urine by ultra pressure LC-ESI-ToF-MS. Metabolomics 2009, 5, 166–182. [Google Scholar] [CrossRef]
- Falade, T.D.O.; Syed Mohdhamdan, S.H.; Sultanbawa, Y.; Fletcher, M.; Harvey, J.J.W.; Chaliha, M.; Fox, G.P. In vitro experimental environments lacking or containing soil disparately affect competition experiments of Aspergillus flavus and co-occurring fungi in maize grains. Food Addit. Contam. 2016, 33, 1241–1253. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Log AFT in Grains ± SD * | |||
|---|---|---|---|
| Dose | Milk | Dough | Dent |
| Control | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Low | 3.88 ± 0.34 b | 3.96 ± 0.44 b | 3.67 ± 0.45 b |
| Medium | 3.73 ± 0.59 b | 4.31 ± 0.34 c,∫ | 4.01 ± 0.33 c |
| High | 3.78 ± 0.21 b | 4.44 ± 0.38 c,∫ | 4.12 ± 0.27 c |
| Log Absol in Grains ± SD * | |||
|---|---|---|---|
| Dose | Milk | Dough | Dent |
| Control | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Low | 2.89 ± 0.34 b | 3.08 ± 0.41 b | 2.99 ± 0.45 b |
| Medium | 2.73 ± 0.59 b,∫ | 3.33 ± 0.35 c | 3.32 ± 0.33 c |
| High | 2.78 ± 0.21 b,∫ | 3.50 ± 0.40 c | 3.42 ± 0.27 c |
| Percentage Classified into Dose Group | |||||
|---|---|---|---|---|---|
| From Dose | Control | Low | Medium | High | Error Counts for Dose |
| Control | 73% | 16% | 2% | 9% | 0.27 |
| Low | 7% | 89% | 4% | 0% | 0.11 |
| Medium | 9% | 2% | 89% | 0% | 0.11 |
| High | 2% | 0% | 2% | 96% | 0.04 |
| Compound | R | p Value |
|---|---|---|
| 1,2-Propanediol | −0.46 | <0.0001 |
| Alanine | −0.41 | <0.0001 |
| Aminomalonic acid | −0.28 | 0.0010 |
| Arabitol or Xylitol | 0.48 | <0.0001 |
| Citric acid | 0.36 | <0.0001 |
| d-erythro-pentitol | 0.12 | 0.1617 |
| Erythronoic acid | −0.41 | <0.0001 |
| Fructose | −0.25 | <0.0030 |
| GABA | −0.39 | <0.0001 |
| Glyceric acid | −0.40 | <0.0001 |
| Glycerol | 0.41 | <0.0001 |
| Glycine | −0.31 | 0.0002 |
| Inositol | 0.45 | <0.0001 |
| Iso-leucine | −0.46 | <0.0001 |
| l-threitol | 0.35 | <0.0001 |
| Malate | 0.18 | 0.0310 |
| Malonic acid | 0.17 | 0.0450 |
| Mannitol | −0.16 | 0.0520 |
| Myo-inositol | −0.47 | <0.0001 |
| Phosphoric acid | 0.21 | 0.0140 |
| Proline | −0.48 | <0.0001 |
| Serine | −0.42 | <0.0001 |
| Sucrose | −0.30 | 0.0003 |
| Succinate | 0.04 | 0.6800 |
| Threonine | −0.44 | <0.0001 |
| Trehalose | −0.49 | <0.0001 |
| Turanose | −0.53 | <0.0001 |
| Uracil | −0.39 | <0.0001 |
| Valine | −0.45 | <0.0001 |
| 3,4-dimethylbenzoic acid (xylylic acid) | −0.08 | 0.3702 |
| Pentadecanoic acid (n-pentadecanoic acid) | 0.15 | 0.0800 |
| Hexadecanoic acid (palmitic acid) | −0.08 | 0.3491 |
| Octadecanoic acid (stearic acid) | 0.02 | 0.8282 |
| 11-icosenoic acid (gondoic acid) | −0.15 | 0.0705 |
| Icosanoic acid (arachidic acid). | −0.03 | 0.7195 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falade, T.D.O.; Chrysanthopoulos, P.K.; Hodson, M.P.; Sultanbawa, Y.; Fletcher, M.; Darnell, R.; Korie, S.; Fox, G. Metabolites Identified during Varied Doses of Aspergillus Species in Zea mays Grains, and Their Correlation with Aflatoxin Levels. Toxins 2018, 10, 187. https://doi.org/10.3390/toxins10050187
Falade TDO, Chrysanthopoulos PK, Hodson MP, Sultanbawa Y, Fletcher M, Darnell R, Korie S, Fox G. Metabolites Identified during Varied Doses of Aspergillus Species in Zea mays Grains, and Their Correlation with Aflatoxin Levels. Toxins. 2018; 10(5):187. https://doi.org/10.3390/toxins10050187
Chicago/Turabian StyleFalade, Titilayo D. O., Panagiotis K. Chrysanthopoulos, Mark P. Hodson, Yasmina Sultanbawa, Mary Fletcher, Ross Darnell, Sam Korie, and Glen Fox. 2018. "Metabolites Identified during Varied Doses of Aspergillus Species in Zea mays Grains, and Their Correlation with Aflatoxin Levels" Toxins 10, no. 5: 187. https://doi.org/10.3390/toxins10050187
APA StyleFalade, T. D. O., Chrysanthopoulos, P. K., Hodson, M. P., Sultanbawa, Y., Fletcher, M., Darnell, R., Korie, S., & Fox, G. (2018). Metabolites Identified during Varied Doses of Aspergillus Species in Zea mays Grains, and Their Correlation with Aflatoxin Levels. Toxins, 10(5), 187. https://doi.org/10.3390/toxins10050187