
Comparison of Quantification of Target-Specific Accumulation of [18F]F-siPSMA-14 in the HET-CAM Model and in Mice Using PET/MRI


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
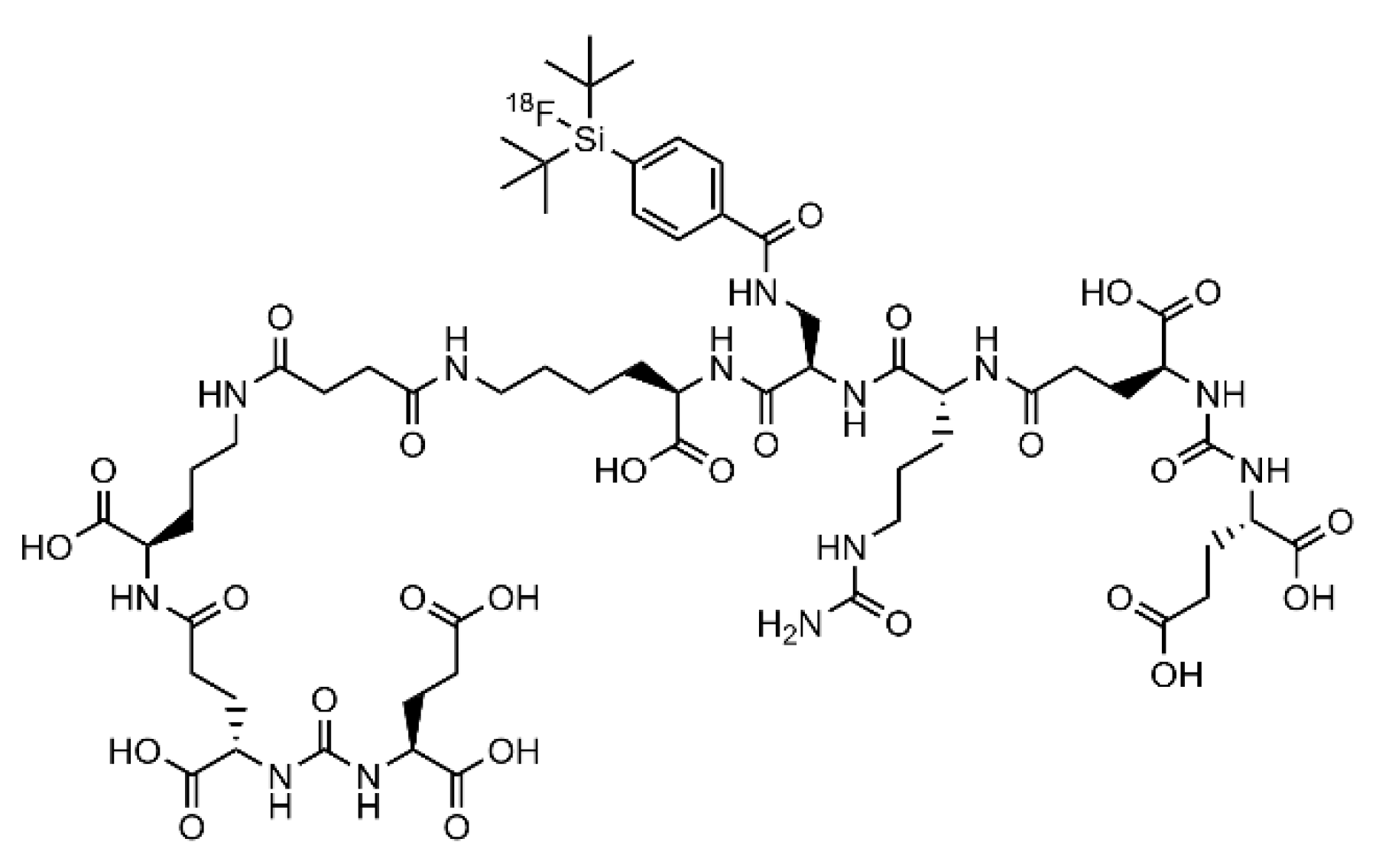
2.1. Synthesis and Radiolabeling
2.2. Cell Culture Preparation
2.3. HET-CAM Experiments
2.4. Animal Studies
2.5. MRI and PET Measurements
2.6. Ex Vivo Validation
2.7. Protein Expression and Histopathological Analyses
2.8. Statistical Evaluation
3. Results
3.1. Tumor Growth and Peptide Accumulation in the Tumors
3.2. Biodistribution
3.3. Evaluation of PSMA-Expression
4. Discussion
4.1. Evaluation of Tumor Accumulation of [18F]F-siPSMA-14 in the HET-CAM Model Compared to a Mouse Model
4.2. Analysis Regarding the Applicability of the HET-CAM Model for Biodistribution Studies in Comparison to the Mouse Model
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Koo, B.-K.; Knoblich, J.A. Human organoids: Model systems for human biology and medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef]
- Gunti, S.; Hoke, A.; Vu, K.; London, N. Organoid and Spheroid Tumor Models: Techniques and Applications. Cancers 2021, 13, 874. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.-Y.; Koh, A.P.-F.; Antony, J.; Huang, R.Y.-J. Applications of the Chick Chorioallantoic Membrane as an Alternative Model for Cancer Studies. Cells Tissues Organs 2021, 1–16. [Google Scholar] [CrossRef]
- Pawlikowska, P.; Tayoun, T.; Oulhen, M.; Faugeroux, V.; Rouffiac, V.; Aberlenc, A.; Pommier, A.L.; Honore, A.; Marty, V.; Bawa, O.; et al. Exploitation of the chick embryo chorioallantoic membrane (CAM) as a platform for anti-metastatic drug testing. Sci. Rep. 2020, 10, 16876. [Google Scholar] [CrossRef]
- Luepke, N.; Kemper, F. The HET-CAM test: An alternative to the draize eye test. Food Chem. Toxicol. 1986, 24, 495–496. [Google Scholar] [CrossRef]
- Herrmann, A.; Taylor, A.; Murray, P.; Poptani, H.; Sée, V. Magnetic Resonance Imaging for Characterization of a Chick Embryo Model of Cancer Cell Metastases. Mol. Imaging 2018, 17, 1536012118809585. [Google Scholar] [CrossRef] [Green Version]
- Fotinos, N.; Campo, M.A.; Popowycz, F.; Gurny, R.; Lange, N. 5-Aminolevulinic Acid Derivatives in Photomedicine: Characteristics, Application and Perspectives. Photochem. Photobiol. 2006, 82, 994–1015. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.; Zeisser-Labouèbe, M.; Lange, N.; Gurny, R.; Delie, F. The chick embryo and its chorioallantoic membrane (CAM) for the in vivo evaluation of drug delivery systems. Adv. Drug Deliv. Rev. 2007, 59, 1162–1176. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Syrovets, T.; Wu, Y.; Hafner, S.; Vernikouskaya, I.; Liu, W.; Ma, G.; Weil, T.; Simmet, T.; Rasche, V. The CAM cancer xenograft as a model for initial evaluation of MR labelled compounds. Sci. Rep. 2017, 7, srep46690. [Google Scholar] [CrossRef] [Green Version]
- Haller, S.; Ametamey, S.M.; Schibli, R.; Müller, C. Investigation of the chick embryo as a potential alternative to the mouse for evaluation of radiopharmaceuticals. Nucl. Med. Biol. 2015, 42, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.G.; Yalcin, M.; Cui, H.; Abdel-Nabi, H.; Sajjad, M.; Bernacki, R.; Veith, J.; Mousa, S.A. Increased tumor uptake of chemotherapeutics and improved chemoresponse by novel non-anticoagulant low molecular weight heparin. Anticancer Res. 2011, 31, 411–419. [Google Scholar]
- Rovithi, M.; Avan, A.; Funel, N.; Leon, L.G.; Gomez, V.E.; Wurdinger, T.; Griffioen, A.W.; Verheul, H.; Giovannetti, E. Development of bioluminescent chick chorioallantoic membrane (CAM) models for primary pancreatic cancer cells: A platform for drug testing. Sci. Rep. 2017, 7, srep44686. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.B.; Berry, R.G. Chorioallantoic membrane heterotransplantation of human brain tumors. Int. J. Cancer 1975, 15, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Strojnik, T.; Kavalar, R.; Barone, T.A.; Plunkett, R.J. Experimental model and immunohistochemical comparison of U87 human glioblastoma cell xenografts on the chicken chorioallantoic membrane and in rat brains. Anticancer Res. 2010, 30, 4851–4860. [Google Scholar]
- Romanoff, A.L. The Avian Embryo. Structural and Functional Development; Macmillan: New York, NY, USA, 1960; p. 1305. [Google Scholar]
- Ribatti, D.; Vacca, A.; De Falco, G.; Ria, R.; Roncali, L.; Dammacco, F. Role of hematopoietic growth factors in angiogenesis. Acta Haematol. 2001, 106, 157–161. [Google Scholar] [CrossRef]
- Ribatti, D. Chapter 5 Chick Embryo Chorioallantoic Membrane as a Useful Tool to Study Angiogenesis. Int. Rev. Cell Mol. Biol. 2008, 270, 181–224. [Google Scholar] [CrossRef]
- Fellah, J.; Jaffredo, T.; Nagy, N.; Dunon, D. Avian Immunology: Development of the Avian Immune System; Schat, K.A., Kaspers, B., Kaiser, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 45–63. [Google Scholar]
- Janse, E.M.; Jeurissen, S.H. Ontogeny and Function of Two Non-Lymphoid Cell Populations in the Chicken Embryo. Immunobiology 1991, 182, 472–481. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane as a model for tumor biology. Exp. Cell Res. 2014, 328, 314–324. [Google Scholar] [CrossRef]
- Komatsu, A.; Higashi, Y.; Matsumoto, K. Various CAM tumor models. Enzymes 2019, 46, 37–57. [Google Scholar] [CrossRef]
- Cimpean, A.M.; Ribatti, D.; Raica, M. The chick embryo chorioallantoic membrane as a model to study tumor metastasis. Angiogenesis 2008, 11, 311–319. [Google Scholar] [CrossRef]
- Deryugina, E.I.; Quigley, J.P. Chick embryo chorioallantoic membrane model systems to study and visualize human tumor cell metastasis. Histochem. Cell Biol. 2008, 130, 1119–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, G.; Koch, A.B.F.; Löffler, J.; Lindén, M.; Solbach, C.; Abaei, A.; Li, H.; Glatting, G.; Beer, A.J.; Rasche, V. Multi-Modal PET and MR Imaging in the Hen’s Egg Test-Chorioallantoic Membrane (HET-CAM) Model for Initial In Vivo Testing of Target-Specific Radioligands. Cancers 2020, 12, 1248. [Google Scholar] [CrossRef]
- Zlatopolskiy, B.D.; Zischler, J.; Schäfer, D.; Urusova, E.A.; Guliyev, M.; Bannykh, O.; Endepols, H.; Neumaier, B. Discovery of 7-[18F]Fluorotryptophan as a Novel Positron Emission Tomography (PET) Probe for the Visualization of Tryptophan Metabolism in Vivo. J. Med. Chem. 2017, 61, 189–206. [Google Scholar] [CrossRef]
- Warnock, G.; Turtoi, A.; Blomme, A.; Bretin, F.; Bahri, M.A.; Lemaire, C.; Libert, L.C.; Seret, A.E.; Luxen, A.; Castronovo, V.; et al. In Vivo PET/CT in a Human Glioblastoma Chicken Chorioallantoic Membrane Model: A New Tool for Oncology and Radiotracer Development. J. Nucl. Med. 2013, 54, 1782–1788. [Google Scholar] [CrossRef] [Green Version]
- Dupertuis, Y.M.; Delie, F.; Cohen, M.; Pichard, C. In ovo method for evaluating the effect of nutritional therapies on tumor development, growth and vascularization. Clin. Nutr. Exp. 2015, 2, 9–17. [Google Scholar] [CrossRef]
- Steinemann, G.; Dittmer, A.; Schmidt, J.; Josuttis, D.; Fähling, M.; Biersack, B.; Beindorff, N.; Koziolek, E.J.; Schobert, R.; Brenner, W.; et al. Antitumor and antiangiogenic activity of the novel chimeric inhibitor animacroxam in testicular germ cell cancer. Mol. Oncol. 2019, 13, 2679–2696. [Google Scholar] [CrossRef]
- Uto, Y.; Abe, C.; Futawaka, M.; Yamada, H.; Tominaga, M.; Endo, Y. In vivo drug screening method of radiosensitizers using tumor-bearing chick embryo. Enzymes 2019, 46, 113–127. [Google Scholar] [CrossRef]
- Cho, C.-F.; Ablack, A.; Leong, H.-S.; Zijlstra, A.; Lewis, J. Evaluation of Nanoparticle Uptake in Tumors in Real Time Using Intravital Imaging. J. Vis. Exp. 2011, 52, 2808. [Google Scholar] [CrossRef] [Green Version]
- Würbach, L.; Heidrich, A.; Opfermann, T.; Gebhardt, P.; Saluz, H.P. Insights into Bone Metabolism of Avian Embryos in Ovo Via 3D and 4D 18F-fluoride Positron Emission Tomography. Mol. Imaging Biol. 2012, 14, 688–698. [Google Scholar] [CrossRef]
- Gebhardt, P.; Würbach, L.; Heidrich, A.; Heinrich, L.; Walther, M.; Opfermann, T.; Sørensen, B.; Saluz, H. Dynamic behaviour of selected PET tracers in embryonated chicken eggs. Rev. Española Med. Nucl. Imagen Mol. 2013, 32, 371–377. [Google Scholar] [CrossRef]
- Mamaeva, V.; Niemi, R.; Beck, M.; Özliseli, E.; Desai, D.; Landor, S.; Gronroos, T.; Kronqvist, P.; Pettersen, I.K.N.; Mc Cormack, E.; et al. Inhibiting Notch Activity in Breast Cancer Stem Cells by Glucose Functionalized Nanoparticles Carrying γ-secretase Inhibitors. Mol. Ther. 2016, 24, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Freesmeyer, M.; Kuehnel, C.; Opfermann, T.; Niksch, T.; Wiegand, S.; Stolz, R.; Huonker, R.; Witte, O.W.; Winkens, T. The Use of Ostrich Eggs for in Ovo Research: Making Preclinical Imaging Research Affordable and Available. J. Nucl. Med. 2018, 59, 1901–1906. [Google Scholar] [CrossRef]
- Winter, G.; Koch, A.B.F.; Löffler, J.; Jelezko, F.; Lindén, M.; Li, H.; Abaei, A.; Zuo, Z.; Beer, A.J.; Rasche, V. In vivo PET/MRI Imaging of the Chorioallantoic Membrane. Front. Phys. 2020, 8, 151. [Google Scholar] [CrossRef]
- Davis, M.I.; Bennett, M.J.; Thomas, L.M.; Bjorkman, P.J. Crystal structure of prostate-specific membrane antigen, a tumor marker and peptidase. Proc. Natl. Acad. Sci. USA 2005, 102, 5981–5986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miksch, J.; Prasad, V.; Di Carlo, D.; Strauss, A.-S.; Grunert, M.; Steinacker, J.; Thaiss, W.; Solbach, C.; Bolenz, C.; Beer, M.; et al. Novel [F-18]siPSMA-14 shows favourable kinetics and high interobserver agreement in staging of prostate cancer patients. J. Nucl. Med. 2021, 62, 1328. [Google Scholar]
- Wu, H.-C.; Hsieh, J.-T.; Gleave, M.E.; Brown, N.M.; Pathak, S.; Chung, L.W.K. Derivation of androgen-independent human LNCaP prostatic cancer cell sublines: Role of bone stromal cells. Int. J. Cancer 1994, 57, 406–412. [Google Scholar] [CrossRef]
- Kaighn, M.E.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.F.; Jones, L.W. Establishment and characterization of a human prostatic carcinoma cell line (PC-3). Investig. Urol. 1979, 17, 16–23. [Google Scholar]
- Thalmann, G.N.; Anezinis, P.E.; Chang, S.M.; Zhau, H.E.; Kim, E.E.; Hopwood, V.L.; Pathak, S.; Von Eschenbach, A.C.; Chung, L.W. Androgen-independent cancer progression and bone metastasis in the LNCaP model of human prostate cancer. Cancer Res. 1994, 54, 2577–2581. [Google Scholar]
- Tai, S.; Sun, Y.; Squires, J.M.; Zhang, H.; Oh, W.; Liang, C.-Z.; Huang, J. PC3 is a cell line characteristic of prostatic small cell carcinoma. Prostate 2011, 71, 1668–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaszewski, M.; Quiros-Gonzalez, I.; O’Connor, J.P.; Abeyakoon, O.; Parker, G.; Williams, K.J.; Gilbert, F.; Bohndiek, S.E. Oxygen Enhanced Optoacoustic Tomography (OE-OT) Reveals Vascular Dynamics in Murine Models of Prostate Cancer. Theranostics 2017, 7, 2900–2913. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.; Winter, G.; Baur, B.; Kuntz, A.J.; Kull, T.; Solbach, C.; Beer, A.J.; Lindén, M. Quantitative and correlative biodistribution analysis of 89 Zr-labeled mesoporous silica nanoparticles intravenously injected into tumor-bearing mice. Nanoscale 2017, 9, 9743–9753. [Google Scholar] [CrossRef] [PubMed]
- Bain, M.M.; Fagan, A.J.; Mullin, J.M.; McNaught, I.; McLean, J.; Condon, B. Noninvasive monitoring of chick development in ovo using a 7T MRI system from day 12 of incubation through to hatching. J. Magn. Reson. Imaging 2007, 26, 198–201. [Google Scholar] [CrossRef]
- Zuo, Z.; Syrovets, T.; Genze, F.; Abaei, A.; Ma, G.; Simmet, T.; Rasche, V. High-resolution MRI analysis of breast cancer xenograft on the chick chorioallantoic membrane. NMR Biomed. 2015, 28, 440–447. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, J.S.; Im, K.C.; Kim, S.J.; Lee, D.S.; Moon, D.H. Performance Measurement of the microPET Focus 120 Scanner. J. Nucl. Med. 2007, 48, 1527–1535. [Google Scholar] [CrossRef] [Green Version]
- Fedorov, A.; Beichel, R.; Kalpathy-Cramer, J.; Finet, J.; Fillion-Robin, J.-C.; Pujol, S.; Bauer, C.; Jennings, D.; Fennessy, F.; Sonka, M.; et al. 3D Slicer as an image computing platform for the Quantitative Imaging Network. Magn. Reson. Imaging 2012, 30, 1323–1341. [Google Scholar] [CrossRef] [Green Version]
- Vollmar, S.; Hampl, J.A.; Kracht, L.; Herholz, K. Integration of Functional Data (PET) into Brain Surgery Planning and Neuronavigation. Adv. Med Eng. 2007, 114, 98–103. [Google Scholar] [CrossRef]
- Laidler, P.; Dulińska, J.; Lekka, M.; Lekki, J. Expression of prostate specific membrane antigen in androgen-independent prostate cancer cell line PC-Arch. Biochem. Biophys. 2005, 435, 1–14. [Google Scholar] [CrossRef]
- Nguyen, D.P.; Xiong, P.L.; Liu, H.; Pan, S.; Leconet, W.; Navarro, V.; Guo, M.; Moy, J.; Kim, S.; Ramirez-Fort, M.K.; et al. Induction of PSMA and Internalization of an Anti-PSMA mAb in the Vascular Compartment. Mol. Cancer Res. 2016, 14, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Waschkies, C.; Nicholls, F.; Buschmann, J. Comparison of medetomidine, thiopental and ketamine/midazolam anesthesia in chick embryos for in Ovo Magnetic Resonance Imaging free of motion artifacts. Sci. Rep. 2015, 5, 15536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojtczak, J.A. The Hemodynamic Effects of Halothane and Isoflurane in Chick Embryo. Anesth. Analg. 2000, 90, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Heidrich, A.; Würbach, L.; Opfermann, T.; Saluz, H.P. Motion-Artifact-Free In Vivo Imaging Utilizing Narcotized Avian Embryos in Ovo. Mol. Imaging Biol. 2010, 13, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, W.; Gregoire, M.-C.; Reilhac, A.; Meikle, S.R. Characterisation of partial volume effect and region-based correction in small animal positron emission tomography (PET) of the rat brain. NeuroImage 2012, 60, 2144–2157. [Google Scholar] [CrossRef] [PubMed]
- Levin, C.S.; Hoffman, E.J. Calculation of positron range and its effect on the fundamental limit of positron emission tomography system spatial resolution. Phys. Med. Biol. 1999, 44, 781–799. [Google Scholar] [CrossRef]
- Palmer, M.; Zhu, X.; Parker, J. Modeling and simulation of positron range effects for high resolution PET imaging. IEEE Trans. Nucl. Sci. 2005, 52, 1391–1395. [Google Scholar] [CrossRef]
- Bailey, D.L.; Karp, J.S.; Surti, S. Physics and instrumentation in PET. In Positron Emission Tomography: Basic Sciences; Bailey, D.L., Townsend, D.W., Valk, P.E., Maisey, M.N., Eds.; Springer: London, UK, 2005; pp. 13–39. [Google Scholar]
- Conti, M.; Eriksson, L. Physics of pure and non-pure positron emitters for PET: A review and a discussion. EJNMMI Phys. 2016, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Attarwala, A.A.; Karanja, Y.W.; Hardiansyah, D.; Romanó, C.; Roscher, M.; Wängler, B.; Glatting, G. Investigation of the imaging characteristics of the ALBIRA II small animal PET system for 18F, 68Ga and 64Cu. Z. Med. Phys. 2017, 27, 132–144. [Google Scholar] [CrossRef]
- Kemerink, G.J.; Visser, M.G.W.; Franssen, R.; Beijer, E.; Zamburlini, M.; Halders, S.G.E.A.; Brans, B.; Mottaghy, F.M.; Teule, G.J.J. Effect of the positron range of 18F, 68Ga and 124I on PET/CT in lung-equivalent materials. Eur. J. Nucl. Med. Mol. Imaging 2011, 38, 940–948. [Google Scholar] [CrossRef]
- Bunka, M.; Müller, C.; Vermeulen, C.; Haller, S.; Türler, A.; Schibli, R.; van der Meulen, N.P. Imaging quality of 44Sc in comparison with five other PET radionuclides using Derenzo phantoms and preclinical PET. Appl. Radiat. Isot. 2016, 110, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Carter, L.M.; Kesner, A.L.; Pratt, E.; Sanders, V.A.; Massicano, A.V.F.; Cutler, C.S.; Lapi, S.E.; Lewis, J.S. The Impact of Positron Range on PET Resolution, Evaluated with Phantoms and PHITS Monte Carlo Simulations for Conventional and Non-conventional Radionuclides. Mol. Imaging Biol. 2019, 22, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Bettinardi, V.; Castiglioni, I.; De Bernardi, E.; Gilardi, M.C. PET quantification: Strategies for partial volume correction. Clin. Transl. Imaging 2014, 2, 199–218. [Google Scholar] [CrossRef]
- Cal-Gonzalez, J.; Vaquero, J.J.; Herraiz, J.L.; Liva, M.P.; Soto-Montenegro, M.L.; Peña-Zalbidea, S.; Desco, M.; Udias, J. Improving PET Quantification of Small Animal [68Ga]DOTA-Labeled PET/CT Studies by Using a CT-Based Positron Range Correction. Mol. Imaging Biol. 2018, 20, 584–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jødal, L.; Le Loirec, C.; Champion, C. Positron range in PET imaging: An alternative approach for assessing and correcting the blurring. Phys. Med. Biol. 2012, 57, 3931–3943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller-Gärtner, H.W.; Links, J.M.; Prince, J.L.; Bryan, R.N.; McVeigh, E.; Leal, J.P.; Davatzikos, C.; Frost, J.J. Measurement of Radiotracer Concentration in Brain Gray Matter Using Positron Emission Tomography: MRI-Based Correction for Partial Volume Effects. Br. J. Pharmacol. 1992, 12, 571–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, H.; Ruest, T.; Schoenahl, F.; Montandon, M.-L. Comparative assessment of statistical brain MR image segmentation algorithms and their impact on partial volume correction in PET. NeuroImage 2006, 32, 1591–1607. [Google Scholar] [CrossRef]
- Gutierrez, D.; Montandon, M.-L.; Assal, F.; Allaoua, M.; Ratib, O.; Lovblad, K.-O.; Zaidi, H. Anatomically guided voxel-based partial volume effect correction in brain PET: Impact of MRI segmentation. Comput. Med Imaging Graph. 2012, 36, 610–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, D.A.; Pellicer, I.; Fair, W.R.; Heston, W.D.; Cordon-Cardo, C. Prostate-specific membrane antigen expression in normal and malignant human tissues. Clin. Cancer Res. 1997, 3, 81–85. [Google Scholar]
- Kinoshita, Y.; Kuratsukuri, K.; Landas, S.; Imaida, K.; Rovito, P.M.; Wang, C.Y.; Haas, G.P. Expression of Prostate-Specific Membrane Antigen in Normal and Malignant Human Tissues. World J. Surg. 2006, 30, 628–636. [Google Scholar] [CrossRef]
- Yang, D.; Holt, G.; Velders, M.P.; Kwon, E.D.; Kast, W.M. Murine six-transmembrane epithelial antigen of the prostate, prostate stem cell antigen, and prostate-specific membrane antigen: Prostate-specific cell-surface antigens highly expressed in prostate cancer of transgenic adenocarcinoma mouse prostate mice. Cancer Res. 2001, 61, 5857–5860. [Google Scholar]
- Schmittgen, T.D.; Zakrajsek, B.A.; Hill, R.E.; Liu, Q.; Reeves, J.J.; Axford, P.D.; Singer, M.J.; Reed, M.W. Expression pattern of mouse homolog of prostate-specific membrane antigen (FOLH1) in the transgenic adenocarcinoma of the mouse prostate model. Prostate 2003, 55, 308–316. [Google Scholar] [CrossRef]
- Kularatne, S.A.; Wang, K.; Santhapuram, H.-K.R.; Low, P. Prostate-Specific Membrane Antigen Targeted Imaging and Therapy of Prostate Cancer Using a PSMA Inhibitor as a Homing Ligand. Mol. Pharm. 2009, 6, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, C.; Xu, X.; Liu, F.; Guo, X.; Li, N.; Wang, X.; Yang, J.; Yang, X.; Zhu, H.; et al. Preclinical Evaluation and Pilot Clinical Study of Al18F-PSMA-BCH for Prostate Cancer PET Imaging. J. Nucl. Med. 2019, 60, 1284–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacich, D.J.; Pinto, J.T.; Tong, W.P.; Heston, W.D. Cloning, expression, genomic localization, and enzymatic activities of the mouse homolog of prostate-specific membrane antigen/NAALADase/folate hydrolase. Mamm. Genome 2001, 12, 117–123. [Google Scholar] [CrossRef]
- Di, L. Strategic Approaches to Optimizing Peptide ADME Properties. AAPS J. 2014, 17, 134–143. [Google Scholar] [CrossRef]
- Bumbaca, B.; Li, Z.; Shah, D.K. Pharmacokinetics of protein and peptide conjugates. Drug Metab. Pharmacokinet. 2019, 34, 42–54. [Google Scholar] [CrossRef]
- Merckx, M.G.; Tay, M.H.; Monaco, M.M.L.; van Zandvoort, M.A.; De Spiegelaere, W.; Lambrichts, I.; Bronckaers, A. Chorioallantoic Membrane Assay as Model for Angiogenesis in Tissue Engineering: Focus on Stem Cells. Tissue Eng. Part B Rev. 2020, 26, 519–539. [Google Scholar] [CrossRef] [PubMed]
- Bolin, G.; Burggren, W.W. Metanephric kidney development in the chicken embryo: Glomerular numbers, characteristics and perfusion. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 343–350. [Google Scholar] [CrossRef]
- Narbaitz, R.; Kacew, S. Ultrastructural and biochemical observations on the metanephros of normal and cultured chick embryos. Anat. Embryol. 1979, 155, 95–105. [Google Scholar] [CrossRef]
- Wideman, R.F. Maturation of glomerular size distribution profiles in domestic fowl (Gallus gallus). J. Morphol. 1989, 201, 205–213. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Wong, G.K.; Cavey, M.J. Development of the liver in the chicken embryo. I. Hepatic cords and sinusoids. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1992, 234, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.K.; Cavey, M.J. Development of the liver in the chicken embryo. II. Erythropoietic and granulopoietic cells. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1993, 235, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Israeli, R.S.; Powell, C.T.; Corr, J.G.; Fair, W.R.; Heston, W.D. Expression of the prostate-specific membrane antigen. Cancer Res. 1994, 54, 1807–1811. [Google Scholar] [PubMed]
- Schmittgen, T.D.; Teske, S.; Vessella, R.L.; True, L.D.; Zakrajsek, B.A. Expression of prostate specific membrane antigen and three alternatively spliced variants of PSMA in prostate cancer patients. Int. J. Cancer 2003, 107, 323–329. [Google Scholar] [CrossRef]
- Pomper, M.G.; Musachio, J.L.; Zhang, J.; Scheffel, U.; Zhou, Y.; Hilton, J.; Maini, A.; Dannals, R.F.; Wong, D.F.; Kozikowski, A.P. 11C-MCG: Synthesis, Uptake Selectivity, and Primate PET of a Probe for Glutamate Carboxypeptidase II (NAALADase). Mol. Imaging 2002, 1, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pullambhatla, M.; Foss, C.A.; Byun, Y.; Nimmagadda, S.; Senthamizhchelvan, S.; Sgouros, G.; Mease, R.C.; Pomper, M.G. 2-(3-{1-Carboxy-5-[(6-[18F]Fluoro-Pyridine-3-Carbonyl)-Amino]-Pentyl}-Ureido)-Pentanedioic Acid, [18F]DCFPyL, a PSMA-Based PET Imaging Agent for Prostate Cancer. Clin. Cancer Res. 2011, 17, 7645–7653. [Google Scholar] [CrossRef] [Green Version]
- Airan, R.D.; Foss, C.A.; Ellens, N.P.K.; Wang, Y.; Mease, R.C.; Farahani, K.; Pomper, M.G. MR-Guided Delivery of Hydrophilic Molecular Imaging Agents Across the Blood-Brain Barrier Through Focused Ultrasound. Mol. Imaging Biol. 2016, 19, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Nico, B.; Bertossi, M. The development of the blood-brain barrier in the chick. Studies with Evans blue and horseradish peroxidase. Ann. Anat. 1993, 175, 85–88. [Google Scholar] [CrossRef]
- Roncali, L.; Nico, B.; Ribatti, D.; Bertossi, M.; Mancini, L. Microscopical and ultrastructural investigations on the development of the blood-brain barrier in the chick embryo optic tectum. Acta Neuropathol. 1986, 70, 193–201. [Google Scholar] [CrossRef]
- Gregersen, M.I.; Rawson, R.A. The disappearance of T-1824 and structurally related dyes from the blood stream. Am. J. Physiol. 1943, 138, 698–707. [Google Scholar] [CrossRef] [Green Version]
- Wolman, M.; Klatzo, I.; Chui, E.; Wilmes, F.; Nishimoto, K.; Fujiwara, K.; Spatz, M. Evaluation of the dye-protein tracers in pathophysiology of the blood-brain barrier. Acta Neuropathol. 1981, 54, 55–61. [Google Scholar] [CrossRef]
- Hu, Q.; Agarwal, U.; Bequette, B.J. Gluconeogenesis, Non-essential amino acid synthesis and substrate partitioning in chicken embryos during later development. Poult. Sci. 2017, 96, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Eckrich, J.; Kugler, P.; Buhr, C.R.; Ernst, B.P.; Mendler, S.; Baumgart, J.; Brieger, J.; Wiesmann, N. Monitoring of tumor growth and vascularization with repetitive ultrasonography in the chicken chorioallantoic-membrane-assay. Sci. Rep. 2020, 10, 18585. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prostate Cancer Cell Line | HET-CAM | Mice | ||
|---|---|---|---|---|
| γ-Counter [%IA/mL] | PET [%IA/mL] | γ-Counter [%IA/g] | PET [%IA/mL] | |
| LNCaP C4-2 (PSMA+) | 12.3 ± 6.4 | 2.8 ± 1.3 | 17.0 ± 10.5 | 9.4 ± 4.4 |
| PC-3 (PSMA−) | 9.3 ± 4.1 | 2.1 ± 1.3 | 1.8 ± 0.4 | 1.6 ± 0.4 |
| Ratio [PSMA+/PSMA−] | 1.5 ± 0.8 | 2.0 ± 1.5 | 9.5 ± 5.1 | 6.4 ± 3.2 |
| Organ | HET-CAM | Mice | |
|---|---|---|---|
| PET [%IA/mL] | PET [%IA/mL] | γ-Counter [%IA/g] | |
| Brain | 1.8 ± 0.5 | 0.3 ± 0.1 | 0.1 ± 0.1 |
| Heart | 8.5 ± 2.0 | 2.6 ± 0.6 | 1.7 ± 0.4 |
| Liver | 7.4 ± 2.0 | 2.3 ± 0.5 | 1.7 ± 0.4 |
| Kidneys | 10.8 ± 4.2 | 47.0 ± 15.3 | 114.7 ± 34.2 |
| (Eye) | 1.8 ± 0.7 | - | - |
| Spleen | - | 12.1 ± 5.7 | 26.3 ± 15.3 |
| Lung | - | 2.1 ± 0.5 | 2.2 ± 0.6 |
| Muscle | - | 0.9 ± 0.2 | 0.8 ± 0.2 |
| Bone | - | 1.1 ± 0.3 | 1.5 ± 0.3 |
| Blood | - | - | 2.5 ± 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Löffler, J.; Hamp, C.; Scheidhauer, E.; Di Carlo, D.; Solbach, C.; Abaei, A.; Hao, L.; Glatting, G.; Beer, A.J.; Rasche, V.; et al. Comparison of Quantification of Target-Specific Accumulation of [18F]F-siPSMA-14 in the HET-CAM Model and in Mice Using PET/MRI. Cancers 2021, 13, 4007. https://doi.org/10.3390/cancers13164007
Löffler J, Hamp C, Scheidhauer E, Di Carlo D, Solbach C, Abaei A, Hao L, Glatting G, Beer AJ, Rasche V, et al. Comparison of Quantification of Target-Specific Accumulation of [18F]F-siPSMA-14 in the HET-CAM Model and in Mice Using PET/MRI. Cancers. 2021; 13(16):4007. https://doi.org/10.3390/cancers13164007
Chicago/Turabian StyleLöffler, Jessica, Carmen Hamp, Ellen Scheidhauer, Daniel Di Carlo, Christoph Solbach, Alireza Abaei, Li Hao, Gerhard Glatting, Ambros J. Beer, Volker Rasche, and et al. 2021. "Comparison of Quantification of Target-Specific Accumulation of [18F]F-siPSMA-14 in the HET-CAM Model and in Mice Using PET/MRI" Cancers 13, no. 16: 4007. https://doi.org/10.3390/cancers13164007
APA StyleLöffler, J., Hamp, C., Scheidhauer, E., Di Carlo, D., Solbach, C., Abaei, A., Hao, L., Glatting, G., Beer, A. J., Rasche, V., & Winter, G. (2021). Comparison of Quantification of Target-Specific Accumulation of [18F]F-siPSMA-14 in the HET-CAM Model and in Mice Using PET/MRI. Cancers, 13(16), 4007. https://doi.org/10.3390/cancers13164007