
MicroRNA Profiling of Red Blood Cells for Lung Cancer Diagnosis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Cohorts and Specimens
2.2. Isolation and Processing of Blood Cells
2.3. Assessment of Purity of Isolated RBCs, PBMCs, and Neutrophils
2.4. RNA Isolation
2.5. Real-Time PCR-Based Microarray Analysis of miRNAs
2.6. Droplet Digital PCR (ddPCR) Analysis of miRNAs
2.7. Statistical Analysis
3. Results
3.1. Isolation and Purification of Blood Cells and RNA Samples
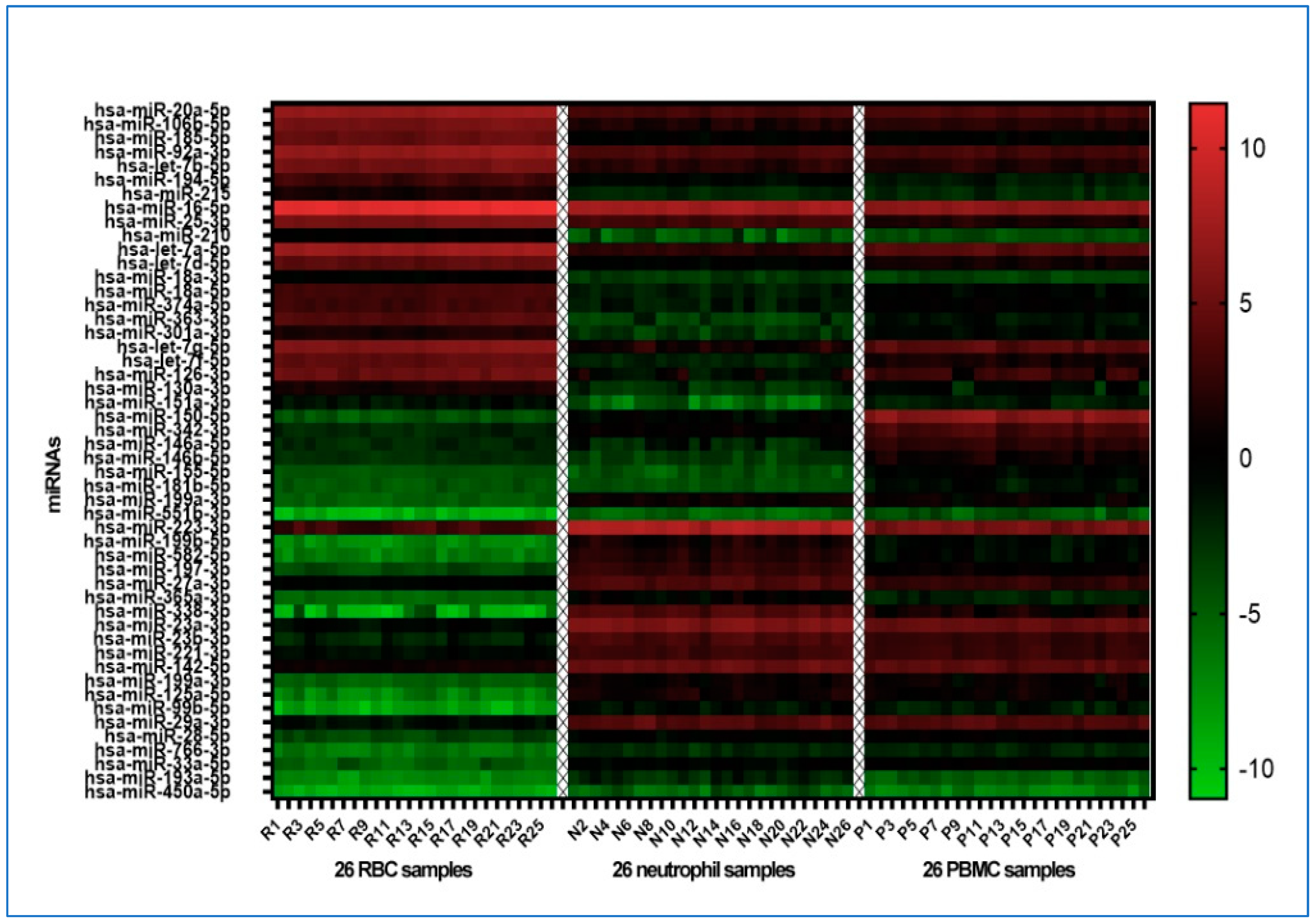
3.2. Distinct miRNA Patterns Can Differentiate among Various Blood Cell Types
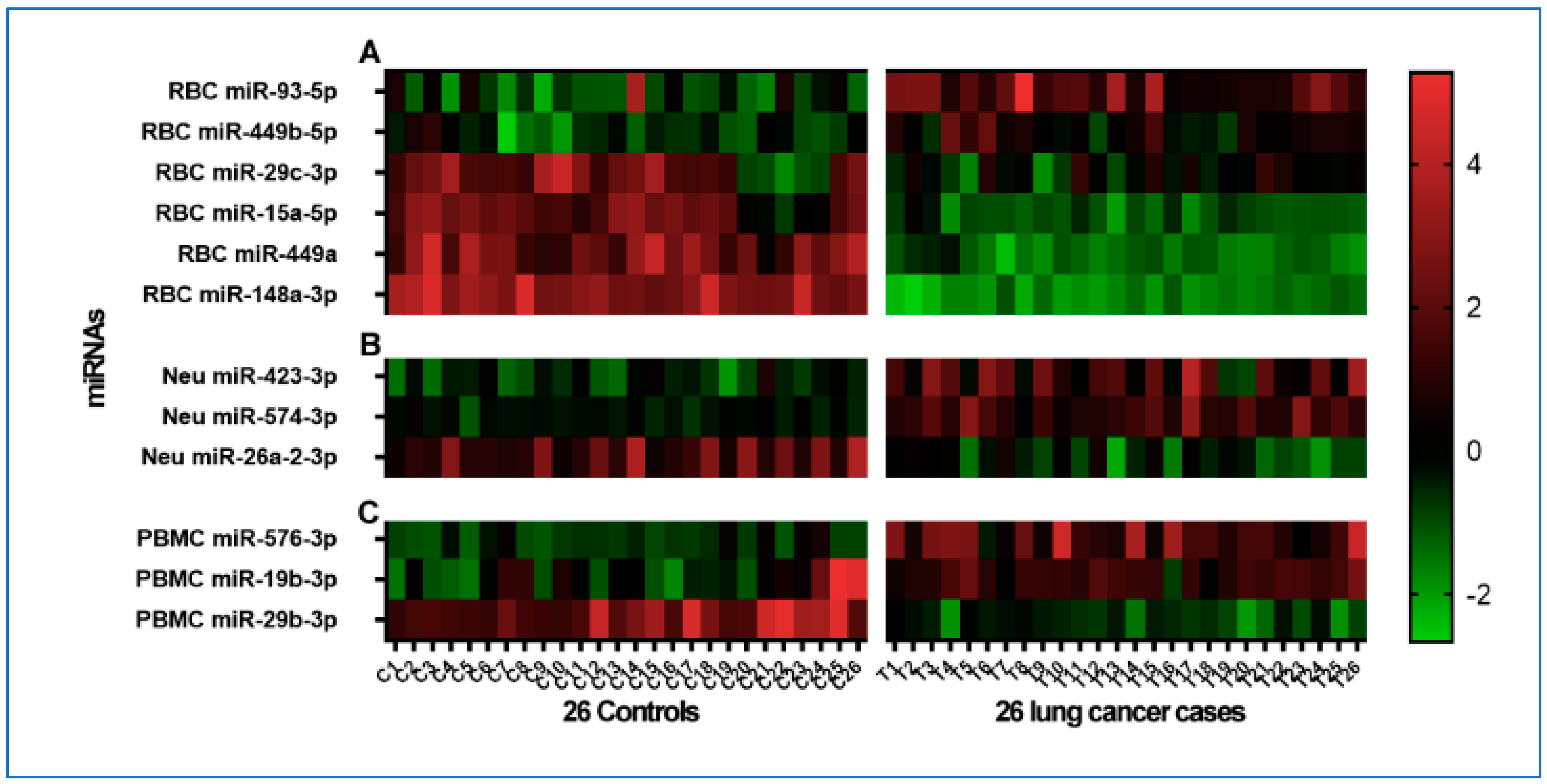
3.3. Distinct Cell-Specific miRNA Profiles Associated with Lung Cancer Patients
3.4. Distinct miRNA Profiles in Cell Types for Differentiating Lung Cancer from Controls
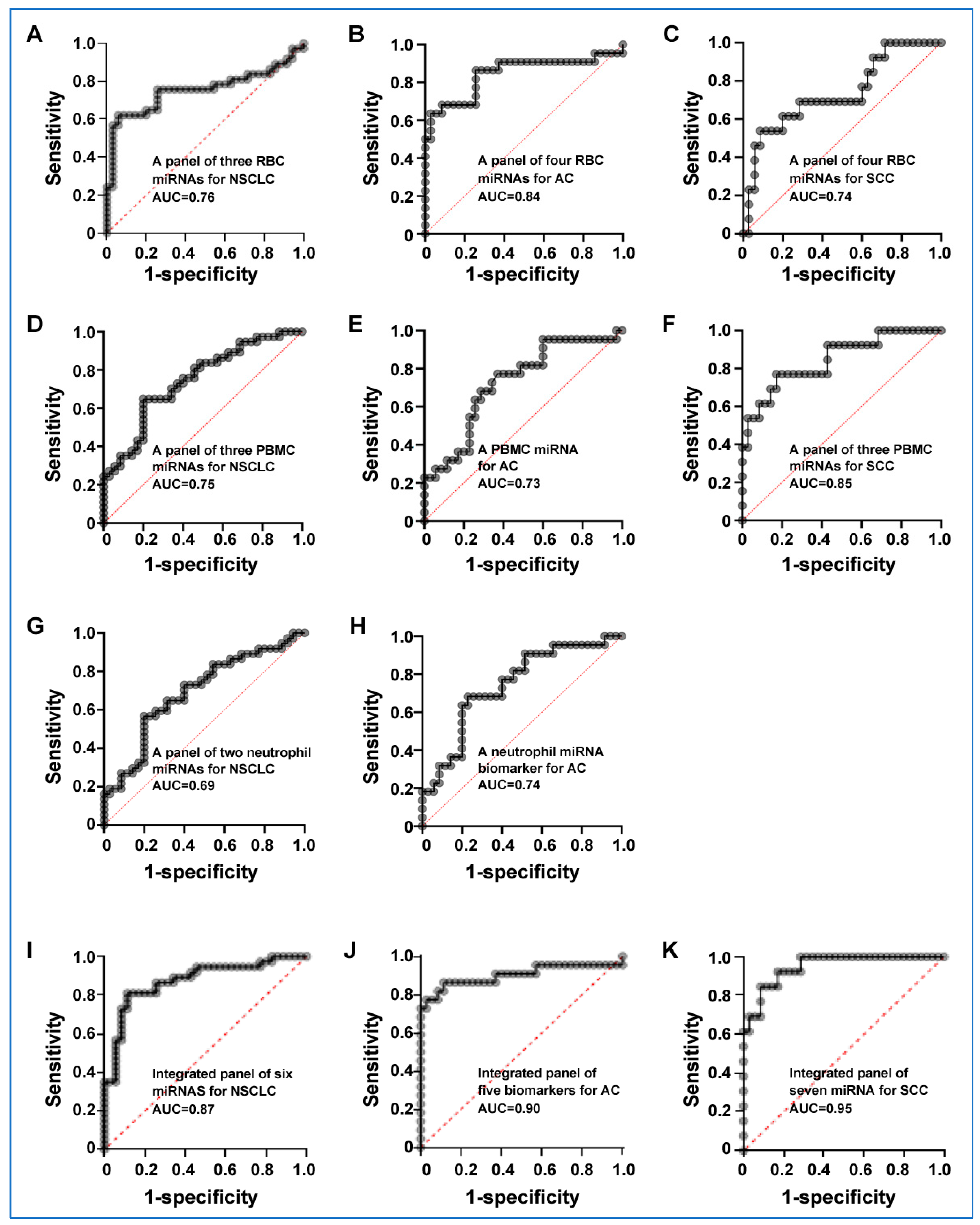
3.5. Integrated miRNAs from Distinct Blood Cell Types Exhibit Synergistic Effects on Lung Cancer Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marmor, H.N.; Zorn, J.T.; Deppen, S.A.; Massion, P.P.; Grogan, E.L. Biomarkers in Lung Cancer Screening: A Narrative Review. Curr. Chall. Thorac. Surg. 2023, 5, 5. [Google Scholar] [CrossRef]
- Moldovan, L.; Batte, K.E.; Trgovcich, J.; Wisler, J.; Marsh, C.B.; Piper, M. Methodological challenges in utilizing miRNAs as circulating biomarkers. J. Cell Mol. Med. 2014, 18, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.L.; Schmittgen, T.D.; et al. Detection of microRNA expression in human peripheral blood microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Pereira-Veiga, T.; Schneegans, S.; Pantel, K.; Wikman, H. Circulating tumor cell-blood cell crosstalk: Biology and clinical relevance. Cell Rep. 2022, 40, 111298. [Google Scholar] [CrossRef]
- Ruffell, B.; Au, A.; Rugo, H.S.; Esserman, L.J.; Hwang, E.S.; Coussens, L.M. Leukocyte composition of human breast cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 2796–2801. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Guo, E.; Mao, X.; Miao, S. Emerging roles of platelets in cancer biology and their potential as therapeutic targets. Front. Oncol. 2022, 12, 939089. [Google Scholar] [CrossRef]
- Ma, J.; Lin, Y.; Zhan, M.; Mann, D.L.; Stass, S.A.; Jiang, F. Differential miRNA expressions in peripheral blood mononuclear cells for diagnosis of lung cancer. Lab. Investig. 2015, 95, 1197–1206. [Google Scholar] [CrossRef]
- Ma, J.; Li, N.; Lin, Y.; Gupta, C.; Jiang, F. Circulating Neutrophil MicroRNAs as Biomarkers for the Detection of Lung Cancer. Biomark. Cancer 2016, 8, BIC.S37333-7. [Google Scholar] [CrossRef]
- Pritchard, C.C.; Kroh, E.; Wood, B.; Arroyo, J.D.; Dougherty, K.J.; Miyaji, M.M.; Tait, J.F.; Tewari, M. Blood cell origin of circulating microRNAs: A cautionary note for cancer biomarker studies. Cancer Prev. Res. 2012, 5, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.H.; Yi, H.S.; Kim, Y.; Kroh, E.M.; Chien, J.W.; Eaton, K.D.; Goodman, M.T.; Tait, J.F.; Tewari, M.; Pritchard, C.C. Plasma processing conditions substantially influence circulating microRNA biomarker levels. PLoS ONE 2013, 8, e64795. [Google Scholar] [CrossRef] [PubMed]
- Morozova, N.; Zinovyev, A.; Nonne, N.; Pritchard, L.L.; Gorban, A.N.; Harel-Bellan, A. Kinetic signatures of microRNA modes of action. RNA 2012, 18, 1635–1655. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA profiling: Approaches and considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef]
- Helwa, R.; Heller, A.; Knappskog, S.; Bauer, A.S. Tumor cells interact with red blood cells via galectin-4—A short report. Cell. Oncol. 2017, 40, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Hernandez, A.; Rodriguez, M.C.; Lopez-Revuelta, A.; Sanchez-Gallego, J.I.; Shnyrov, V.; Llanillo, M.; Sanchez-Yague, J. Alterations in erythrocyte membrane protein composition in advanced non-small cell lung cancer. Blood Cells Mol. Dis. 2006, 36, 355–363. [Google Scholar] [CrossRef]
- Amezaga, J.; Arranz, S.; Urruticoechea, A.; Ugartemendia, G.; Larraioz, A.; Louka, M.; Uriarte, M.; Ferreri, C.; Tueros, I. Altered Red Blood Cell Membrane Fatty Acid Profile in Cancer Patients. Nutrients 2018, 10, 1853. [Google Scholar] [CrossRef]
- Valles, J.; Santos, M.T.; Aznar, J.; Martinez, M.; Moscardo, A.; Pinon, M.; Broekman, M.J.; Marcus, A.J. Platelet-erythrocyte interactions enhance alpha(IIb)beta(3) integrin receptor activation and P-selectin expression during platelet recruitment: Down-regulation by aspirin ex vivo. Blood 2002, 99, 3978–3984. [Google Scholar] [CrossRef]
- Helms, C.C.; Marvel, M.; Zhao, W.; Stahle, M.; Vest, R.; Kato, G.J.; Lee, J.S.; Christ, G.; Gladwin, M.T.; Hantgan, R.R.; et al. Mechanisms of hemolysis-associated platelet activation. J. Thromb. Haemost. 2013, 11, 2148–2154. [Google Scholar] [CrossRef]
- Lam, L.K.M.; Murphy, S.; Kokkinaki, D.; Venosa, A.; Sherrill-Mix, S.; Casu, C.; Rivella, S.; Weiner, A.; Park, J.; Shin, S.; et al. DNA binding to TLR9 expressed by red blood cells promotes innate immune activation and anemia. Sci. Transl. Med. 2021, 13, eabj1008. [Google Scholar] [CrossRef]
- Liang, N.; Jiao, Z.; Zhang, C.; Wu, Y.; Wang, T.; Li, S.; Wang, Y.; Song, T.; Chen, J.Q.; Liang, H.; et al. Mature Red Blood Cells Contain Long DNA Fragments and Could Acquire DNA from Lung Cancer Tissue. Adv. Sci. 2023, 10, e2206361. [Google Scholar] [CrossRef]
- Hotz, M.J.; Qing, D.; Shashaty, M.G.S.; Zhang, P.; Faust, H.; Sondheimer, N.; Rivella, S.; Worthen, G.S.; Mangalmurti, N.S. Red Blood Cells Homeostatically Bind Mitochondrial DNA through TLR9 to Maintain Quiescence and to Prevent Lung Injury. Am. J. Respir. Crit. Care Med. 2018, 197, 470–480. [Google Scholar] [CrossRef]
- Dobkin, J.; Mangalmurti, N.S. Immunomodulatory roles of red blood cells. Curr. Opin. Hematol. 2022, 29, 306–309. [Google Scholar] [CrossRef]
- Karsten, E.; Breen, E.; McCracken, S.A.; Clarke, S.; Herbert, B.R. Red blood cells exposed to cancer cells in culture have altered cytokine profiles and immune function. Sci. Rep. 2020, 10, 7727. [Google Scholar] [CrossRef] [PubMed]
- Mintz, J.; Vedenko, A.; Rosete, O.; Shah, K.; Goldstein, G.; Hare, J.M.; Ramasamy, R.; Arora, H. Current Advances of Nitric Oxide in Cancer and Anticancer Therapeutics. Vaccines 2021, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Ellingsen, T.S.; Lappegard, J.; Skjelbakken, T.; Braekkan, S.K.; Hansen, J.B. Impact of red cell distribution width on future risk of cancer and all-cause mortality among cancer patients—The Tromso Study. Haematologica 2015, 100, e387–e389. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.H.; Chacko, B.K.; Kevil, C.G.; Patel, R.P. The red blood cell and vascular function in health and disease. Antioxid. Redox Signal 2004, 6, 992–999. [Google Scholar] [CrossRef]
- Miao, Y.; Yang, Y.; Guo, L.; Chen, M.; Zhou, X.; Zhao, Y.; Nie, D.; Gan, Y.; Zhang, X. Cell Membrane-Camouflaged Nanocarriers with Biomimetic Deformability of Erythrocytes for Ultralong Circulation and Enhanced Cancer Therapy. ACS Nano 2022, 16, 6527–6540. [Google Scholar] [CrossRef]
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef]
- Papadopoulos, C. Immunosuppressive function of intratumor red blood cells. An immunometabolic perspective. Curr. Cancer Ther. Rev. 2022, 4, 224–226. [Google Scholar] [CrossRef]
- Allantaz, F.; Cheng, D.T.; Bergauer, T.; Ravindran, P.; Rossier, M.F.; Ebeling, M.; Badi, L.; Reis, B.; Bitter, H.; D’Asaro, M.; et al. Expression profiling of human immune cell subsets identifies miRNA-mRNA regulatory relationships correlated with cell type specific expression. PLoS ONE 2012, 7, e29979. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ma, J.; Guarnera, M.A.; Fang, H.; Cai, L.; Jiang, F. Digital PCR quantification of miRNAs in sputum for diagnosis of lung cancer. J. Cancer Res. Clin. Oncol. 2014, 140, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, N.; Guarnera, M.; Jiang, F. Quantification of Plasma miRNAs by Digital PCR for Cancer Diagnosis. Biomark. Insights 2013, 8, 127–136. [Google Scholar] [CrossRef]
- Ma, J.; Mannoor, K.; Gao, L.; Tan, A.; Guarnera, M.A.; Zhan, M.; Shetty, A.; Stass, S.A.; Xing, L.; Jiang, F. Characterization of microRNA transcriptome in lung cancer by next-generation deep sequencing. Mol. Oncol. 2014, 8, 1208–1219. [Google Scholar] [CrossRef]
- Su, J.; Liao, J.; Gao, L.; Shen, J.; Guarnera, M.A.; Zhan, M.; Fang, H.; Stass, S.A.; Jiang, F. Analysis of small nucleolar RNAs in sputum for lung cancer diagnosis. Oncotarget 2016, 7, 5131–5142. [Google Scholar] [CrossRef]
- Gupta, C.; Su, J.; Zhan, M.; Stass, S.A.; Jiang, F. Sputum long non-coding RNA biomarkers for diagnosis of lung cancer. Cancer Biomark. 2019, 26, 219–227. [Google Scholar] [CrossRef]
- Liao, J.; Yu, L.; Mei, Y.; Guarnera, M.; Shen, J.; Li, R.; Liu, Z.; Jiang, F. Small nucleolar RNA signatures as biomarkers for non-small-cell lung cancer. Mol. Cancer 2010, 9, 198. [Google Scholar] [CrossRef]
- Shen, J.; Todd, N.W.; Zhang, H.; Yu, L.; Lingxiao, X.; Mei, Y.; Guarnera, M.; Liao, J.; Chou, A.; Lu, C.L.; et al. Plasma microRNAs as potential biomarkers for non-small-cell lung cancer. Lab. Investig. 2011, 91, 579–587. [Google Scholar] [CrossRef]
- Zaporozhchenko, I.A.; Morozkin, E.S.; Skvortsova, T.E.; Ponomaryova, A.A.; Rykova, E.Y.; Cherdyntseva, N.V.; Polovnikov, E.S.; Pashkovskaya, O.A.; Pokushalov, E.A.; Vlassov, V.V.; et al. Plasma miR-19b and miR-183 as Potential Biomarkers of Lung Cancer. PLoS ONE 2016, 11, e0165261. [Google Scholar] [CrossRef]
- Zaporozhchenko, I.A.; Morozkin, E.S.; Ponomaryova, A.A.; Rykova, E.Y.; Cherdyntseva, N.V.; Zheravin, A.A.; Pashkovskaya, O.A.; Pokushalov, E.A.; Vlassov, V.V.; Laktionov, P.P. Profiling of 179 miRNA Expression in Blood Plasma of Lung Cancer Patients and Cancer-Free Individuals. Sci. Rep. 2018, 8, 6348. [Google Scholar] [CrossRef]
- Yang, W.; Bai, J.; Liu, D.; Wang, S.; Zhao, N.; Che, R.; Zhang, H. MiR-93-5p up-regulation is involved in non-small cell lung cancer cells proliferation and migration and poor prognosis. Gene 2018, 647, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Li, H.X.; Li, P.P.; Guo, Z.J.; Yang, Y. MicroRNA-449b-3p inhibits epithelial-mesenchymal transition by targeting IL-6 and through the JAK2/STAT3 signaling pathway in non-small cell lung cancer. Exp. Ther. Med. 2020, 19, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Jin, K.; Dong, R.; Gao, C.; Si, L.; Feng, Z.; Zhang, H.; Tian, H. Negatively Regulated by miR-29c-3p, MTFR1 Promotes the Progression and Glycolysis in Lung Adenocarcinoma via the AMPK/mTOR Signalling Pathway. Front. Cell Dev. Biol. 2021, 9, 771824. [Google Scholar] [CrossRef]
- Ni, Y.; Yang, Y.; Ran, J.; Zhang, L.; Yao, M.; Liu, Z. miR-15a-5p inhibits metastasis and lipid metabolism by suppressing histone acetylation in lung cancer. Free Radic. Biol. Med. 2020, 161, 150–162. [Google Scholar] [CrossRef]
- Liang, L.; Xu, W.Y.; Shen, A.; Cen, H.Y.; Chen, Z.J.; Tan, L.; Zhang, L.M.; Zhang, Y.; Fu, J.J.; Qin, A.P.; et al. Promoter methylation-regulated miR-148a-3p inhibits lung adenocarcinoma (LUAD) progression by targeting MAP3K9. Acta Pharmacol. Sin. 2022, 43, 2946–2955. [Google Scholar] [CrossRef]
- Greenawalt, E.J.; Edmonds, M.D.; Jain, N.; Adams, C.M.; Mitra, R.; Eischen, C.M. Targeting of SGK1 by miR-576-3p Inhibits Lung Adenocarcinoma Migration and Invasion. Mol. Cancer Res. 2019, 17, 289–298. [Google Scholar] [CrossRef]
- Li, Z.L.; Li, D.; Yin, G.Q. MiR-19b-3p promotes tumor progression of non-small cell lung cancer via downregulating HOXA9 and predicts poor prognosis in patients. Histol. Histopathol. 2022, 37, 779–789. [Google Scholar] [CrossRef]
- Ma, J.; Huang, W.; Zhu, C.; Sun, X.; Zhang, Q.; Zhang, L.; Qi, Q.; Bai, X.; Feng, Y.; Wang, C. miR-423-3p activates FAK signaling pathway to drive EMT process and tumor growth in lung adenocarcinoma through targeting CYBRD1. J. Clin. Lab. Anal. 2021, 35, e24044. [Google Scholar] [CrossRef]
- Zhou, R.; Zhou, X.; Yin, Z.; Guo, J.; Hu, T.; Jiang, S.; Liu, L.; Dong, X.; Zhang, S.; Wu, G. MicroRNA-574-5p promotes metastasis of non-small cell lung cancer by targeting PTPRU. Sci. Rep. 2016, 6, 35714. [Google Scholar] [CrossRef]
- Liu, B.; Wu, X.; Wang, C.; Liu, Y.; Zhou, Q.; Xu, K. MiR-26a enhances metastasis potential of lung cancer cells via AKT pathway by targeting PTEN. Biochim. Biophys. Acta 2012, 1822, 1692–1704. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Wagner, P.D. The Early Detection Research Network: A National Infrastructure to Support the Discovery, Development, and Validation of Cancer Biomarkers. Cancer Epidemiol. Biomark. Prev. 2020, 29, 2401–2410. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| An Exploratory Set | A Validation Set | ||||
|---|---|---|---|---|---|
| NSCLC cases (n = 26) | Controls (n = 26) | NSCLC cases (n = 42) | Controls (n = 39) | ||
| Age | 66.73 (SD 11.25) | 65.782 (SD 10.39) | Age | 65.38 (SD 10.23) | 64.74 (SD 10.26) |
| Sex | Sex | ||||
| Female | 9 | 9 | Female | 12 | 11 |
| Male | 17 | 17 | Male | 30 | 28 |
| Race | Race | ||||
| AAs | 11 | 11 | AAs | 26 | 25 |
| WAs | 16 | 16 | WAs | 16 | 14 |
| Smoking pack-years (median) | 32.7 | 30.9 | Smoking pack-years (median) | 33.8 | 31.2 |
| Pulmonary nodule size (mm) | 21.26 (SD 10.25) | 6.72 (SD 3.58) | Pulmonary nodule size (mm) | 20.25 (SD 11.38) | 6.63 (SD 3.97) |
| Stage | Stage | ||||
| Stage I | 7 | Stage I | 10 | ||
| Stage II | 5 | Stage II | 8 | ||
| Stage III | 8 | Stage III | 9 | ||
| Stage IV | 6 | Stage IV | 12 | ||
| Unknown | 3 | ||||
| Histological type | Histological type | ||||
| Adenocarcinoma | 15 | AC | 26 | ||
| SCC | 11 | SCC | 16 | ||
| miRNAs | Mean Expression (Controls) | Mean Expression (Cancer Patients) | Mann–Whitney U Statistic | FDR-Adjusted p-Value |
|---|---|---|---|---|
| RBC miR-93-5p | 0.922 | 1.529 | 41 | <0.001 |
| RBC miR-449b-5p | −0.585 | 0.370 | 116 | <0.001 |
| RBC miR-29c-3p | −0.585 | −0.145 | 189 | 0.006 * |
| RBC miR-15a-5p | 2.039 | −1.041 | 10 | <0.001 * |
| RBC miR-449a | 2.431 | −1.299 | 0 | <0.001 * |
| RBC miR-148a-3p | 2.728 | −1.701 | 0 | <0.001 * |
| Neutrophil miR-423-3p | −0.464 | 1.550 | 75 | <0.001 |
| Neutrophil miR-574-3p | −0.255 | 1.014 | 2 | <0.001 |
| Neutrophil miR-26a-2-3p | 0.926 | −0.454 | 6 | <0.001 * |
| PBMC miR-576-3p | −0.754 | 1.476 | 16 | <0.001 |
| PBMC miR-19b-3p | −0.105 | 1.219 | 126 | <0.001 |
| PBMC miR-29b-3p | 1.739 | −0.528 | 0 | <0.001 * |
| miRNAs | Mean Expression (Controls) | Mean Expression (Cancer Patients) | Mann–Whitney U Statistic | FDR-Adjusted p-Value |
|---|---|---|---|---|
| RBC miR-93-5p | 38.443 | 52.000 | 474 | 0.009 |
| RBC miR-449b-5p | 0.016 | 0.022 | 631 | 0.008 |
| RBC miR-29c-3p | 3.930 | 3.030 | 561 | 0.014 * |
| RBC miR-15a-5p | 151.015 | 112.129 | 506 | 0.003 * |
| PBMC miR-576-3p | 0.045 | 0.056 | 576 | 0.021 |
| PBMC miR-19b-3p | 24.834 | 27.016 | 515 | 0.004 |
| PBMC miR-29b-3p | 8.220 | 6.855 | 438 | 0.001 * |
| Neutrophil miR-574-3p | 0.605 | 0.790 | 532 | 0.010 |
| Neutrophil miR-26a-2-3p | 0.031 | 0.019 * | 397 | 0.003 * |
| AUC, % (95% CI) | Sensitivity, % (95% CI) | Specificity, % (95% CI) | |
|---|---|---|---|
| A panel of three RBC miRNA biomarkers for NSCLC | 0.76 (0.64 to 0.87) | 77.42% (58.90% to 90.41%) | 68.29% (51.91% to 81.92%) |
| A panel of four RBC miRNA biomarkers for AC | 0.84 (0.72 to 0.96) | 87.50% (61.65% to 98.45%) | 80.49% (65.13% to 91.18%) |
| A panel of four RBC miRNA biomarkers for SCC | 0.74 (0.57 to 0.90) | 71.43% (29.04% to 96.33%) | 80.49% (65.13% to 91.18%) |
| A panel of three PBMC miRNA biomarkers for NSCLC | 0.75 (0.63 to 0.86) | 70.59% (52.52% to 84.90%) | 65.79% (48.65% to 80.37%) |
| A PBMC miRNA biomarker for AC | 0.73 (0.59 to 0.86) | 57.14% (28.86% to 82.34%) | 67.44% (51.46% to 80.92%) |
| A panel of three PBMC miRNA biomarkers for SCC | 0.85 (0.72to 0.97) | 72.73% (39.03% to 93.98%) | 86.49% (71.23% to 95.46%) |
| A panel of two neutrophil miRNA biomarkers for NSCLC | 0.69 (0.57 to 0.82) | 64.10% (47.18% to 78.80%) | 63.64% (45.12% to 79.60%) |
| A neutrophil miRNA biomarker for AC | 0.74 (0.61 to 0.87) | 53.33% (26.59% to 78.73%) | 66.67% (50.45% to 80.43%) |
| Integrated panel of biomarkers for NSCLC | 0.87 (0.79to 0.96) | 8056%, (63.98% to 91.81%) | 83.33% (67.19% to 93.63% |
| Integrated panel of biomarkers for AC | 0.90 (0.79 to 1.00) | 85.00% (62.11% to 96.79%) | 86.49% (71.23% to 95.46%) |
| Integrated panel of biomarkers for SCC | 0.95 (0.89 to 1.00) | 81.82% (48.22% to 97.72%) | 89.19% (74.58% to 96.97%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, X.; Ma, J.; Dhilipkannah, P.; Jiang, F. MicroRNA Profiling of Red Blood Cells for Lung Cancer Diagnosis. Cancers 2023, 15, 5312. https://doi.org/10.3390/cancers15225312
Geng X, Ma J, Dhilipkannah P, Jiang F. MicroRNA Profiling of Red Blood Cells for Lung Cancer Diagnosis. Cancers. 2023; 15(22):5312. https://doi.org/10.3390/cancers15225312
Chicago/Turabian StyleGeng, Xinyan, Jie Ma, Pushpa Dhilipkannah, and Feng Jiang. 2023. "MicroRNA Profiling of Red Blood Cells for Lung Cancer Diagnosis" Cancers 15, no. 22: 5312. https://doi.org/10.3390/cancers15225312
APA StyleGeng, X., Ma, J., Dhilipkannah, P., & Jiang, F. (2023). MicroRNA Profiling of Red Blood Cells for Lung Cancer Diagnosis. Cancers, 15(22), 5312. https://doi.org/10.3390/cancers15225312