Amino Acids in Entomopathogenic Fungi Cultured In Vitro


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Fungi Characterisation
2.1.2. Reagents
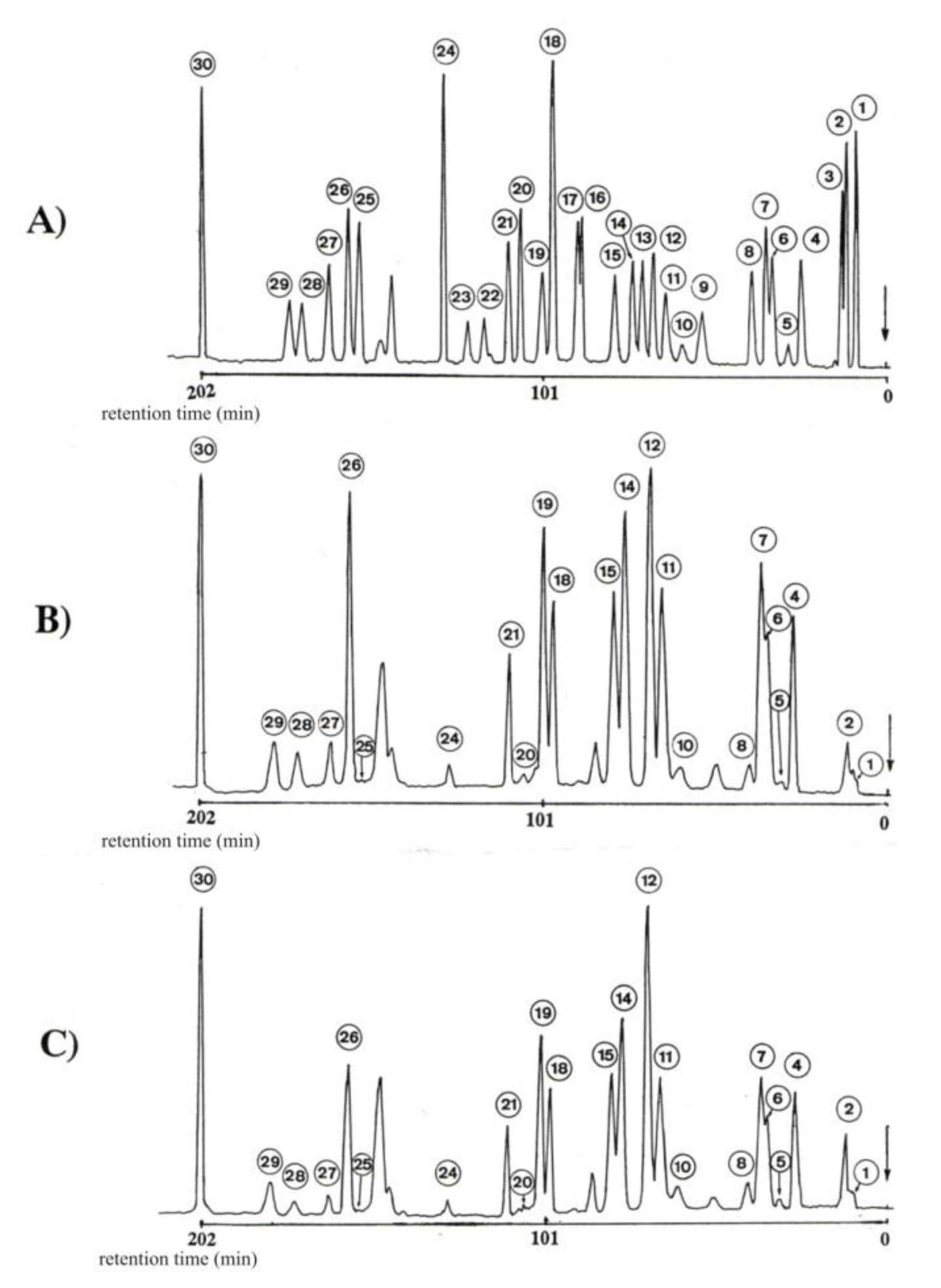
Determination of Bound Amino Acids in Hydrolysates Mycelium


2.2. Method for the Separation and Determination of Amino Acids in Mycelium Hydrolysates
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Integrated Pest Management (IPM). 2016. Available online: https://www2.ipm.ucanr.edu/What-is-IPM/ (accessed on 31 December 2006).
- Madden, U.A.; Stahr, H.M. Preliminary determination of mycotoxin binding to soil when leaching through soil with water. Int. Biodeterior. Biodegrad. 1993, 31, 265–275. [Google Scholar] [CrossRef]
- Liu, X.; Li, S. Fungal secondary metabolites in biological control of crop pests. In Handbook of Industrial Mycology; An, Z., Ed.; Marcel Dekker: New York, NY, USA, 2004; p. 732. [Google Scholar]
- Zimmermann, G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Meyer-Wolfarth, F.; Oldenburg, E.; Meiners, T.; Munoz, K.; Schrader, S. Farmer’s Little Helper-Earthworms’ and Collembolans’ Potential Role in Pathogen Repression and Reduction of Environmental Contaminants; Geophysical Research Abstracts; European Geosciences Union: Vienna, Austria, 2018; Volume 20, p. EGU2018-1782. [Google Scholar]
- Ryszkowski, L.; Karg, J. The effect of the structure of agricultural landscape on biomass of insect of the above-ground fauna. Ekol. Pol. 1991, 39, 171–179. [Google Scholar]
- Tkaczuk, C.; Krzyczkowski, T.; Wegensteiner, R. The occurrence of entomopathogenic fungi in soils from mid-field woodlots and adjacent small-scale arable fields. Acta Mycol. 2012, 47, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Steyn, P.S. The Biosynthesis of Mycotoxins—A Study in Secondary Metabolism; Academic Press: London, UK, 1980; p. 448. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the Surface: Entomopathogenic Fungi versus the Insect Cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Kodaira, Y. Biochemical studies on the muscardine fungi in the silkworms, Bombyx mori. J. Fac. Text. Sci. Technol. Shinshu Univ. Ser. A Biol. 1961, 29, 1–68. [Google Scholar]
- Stevenson, G. Ściana Komórki. Biologia Grzybów, Bakterii i Wirusów (Cell Wall. Biology of Fungi, Bacteria and Viruses); PWRiL: Warszawa, Poland, 1972; p. 278. (In Polish) [Google Scholar]
- Rose, A.H. Molecular architecture. In Chemical Microbiology, an Introduction to Microbial Physiology; Rose, A.H., Ed.; Butter Worths: London, UK, 1976; pp. 5–92. [Google Scholar]
- Vaziri-Tehrani, B.; Dick, M. Taxonomic significance of variations between ratios of cell wall amino acids in fungi, with special reference to Oomycetes. Trans. Br. Mycol. Soc. 1980, 74, 231–238. [Google Scholar] [CrossRef]
- Takatsuki, A.; Tamura, G.; Arima, K. Mode of action of xanthocillin × monomethylether on multiplication of newcastle disease virus in cultured cells. J. Antibiot. 1969, 22, 151–160. [Google Scholar] [CrossRef]
- Ho, P.K.; Walters, C.P. Specific inhibition of infuenza virus-induced ribonucleic acid polimerase by gliotoxin. Antimicrob. Agents Chemother. 1969, 29, 68–74. [Google Scholar]
- Trown, P. Antiviral activity of N,N′-dimethyl-epidithiapiperazinedione, a synthetic compound related to the gliotoxins, LL-S88α and β, chetomin and the sporidesmins. Biochem. Biophys. Res. Commun. 1968, 33, 402–407. [Google Scholar] [CrossRef]
- Kidd, J.G. Effects of an Antibiotic From Aspergillus fumigatus Fresenius on Tumor Cells In Vitro, and Its Possible Identity with Gliotoxin. Science 1974, 105, 511–513. [Google Scholar] [CrossRef]
- Liu, C.-X.; Zhang, J.; Xiang, W.; Qian, P.-T.; Wang, J.-D.; Gao, Y.-M.; Yan, Y.-J.; Zhang, S.-Z.; Xu, P.-F.; Li, W.; et al. Antifungal Activity of Borrelidin Produced by a Streptomyces Strain Isolated from Soybean. J. Agric. Food Chem. 2012, 60, 1251–1257. [Google Scholar] [CrossRef]
- Valiante, V. The Cell Wall Integrity Signaling Pathway and Its Involvement in Secondary Metabolite Production. J. Fungi 2017, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Li, F.; Zhang, Y. Risks of Mycotoxins from Mycoinsecticides to Humans. BioMed Res. Int. 2016, 12, 3194321. [Google Scholar] [CrossRef] [Green Version]
- Machowicz-Stefaniak, Z. Wpływ stałych podłoży hodowlanych na wzrost i rozwój entomopatogennego grzyba Paecilomyces farinosus (Dicks. Ex Fr.) Brown et Smith (The impact of solid culture media on the growth and development of the fungus enthomopathogenic Paecilomyces farinosus (Dicks. Ex Fr.) Brown et Smith). Roczn. Nauk Roln. 1988, 18, 171–177. (In Polish) [Google Scholar]
- Wang, Z.; Meng, H.; Zhuang, Z.; Chen, M.; Xie, L.; Huang, B. Molecular cloning of a novel subtilisin-like protease (Pr1A) gene from the biocontrol fungus Isaria farinosa. Appl. Entomol. Zool. 2013, 48, 477–487. [Google Scholar] [CrossRef]
- Cantone, F.A.; Vandenberg, J.D. Intraspecific diversity in Paecilomyces fumosoroseus. Mycol. Res. 1998, 102, 209–215. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Jackson, M.A.; Wright, M.S. A foam formulation ofPaecilomyces fumosoroseus, an entomopathogenic biocontrol agent. Biocontrol Sci. Technol. 2007, 17, 513–523. [Google Scholar] [CrossRef]
- Hoy, M.A.; Singh, R.; Rogers, M.E. Evaluations of a Novel Isolate of Isaria fumosorosea for Control of the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae). Fla. Entomol. 2010, 93, 24–32. [Google Scholar] [CrossRef]
- Ownley, B.H.; Gwinn, K.; Vega, F.E. Endophytic fungal entomopathogens with activity against plant pathogens: Ecology and evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Cerkowniak, M.; Ostachowska, A.; Naczk, A.M.; Boguś, M.I.; Stepnowski, P. Effect of Conidiobolus coronatus on the Cuticular and Internal Lipid Composition of Tettigonia viridissima Males. Chem. Biodivers. 2016, 13, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Paszkiewicz, M.; Tyma, M.; Ligęza-Żuber, M.; Włóka, E.; Boguś, M.; Monika, P. Trichothecenes Production by Entomopathogenic Fungus Conidiobolus Coronatus. Adv. Toxicol. Toxic Eff. 2016, 1, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Bałazy, S.; Miętkiewski, R.; Majchrowicz, I. Mikozy mszyc-ich znaczenie i perspektywy wykorzystania w ochronie roślin (Mycosis of aphids—Their importance and utilization ways in the plant protection). Zesz. Probl. Post. Nauk Roln. 1990, 392, 35–56. (In Polish) [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi; Academic Press: London, UK; New York, NY, USA; Toronto, ON, Canada; Sydney, Australia; San Francisco, CA, USA, 1980; Volume 1, p. 859. [Google Scholar]
- Tkaczuk, C.; Król, A.; Majchrowska-Safaryan, A.; Nicewicz, Ł. The occurrence of entomopathogenic fungi in soils from fields cultivated in a conventional and organic system. J. Ecol. Eng. 2014, 15, 137–144. [Google Scholar] [CrossRef]
- Bałazy, S.; Wrzosek, M.; Sosnowska, D.; Tkaczuk, C.; Muszewska, A. Laboratory trials to infect insects and nematodes by some acaropathogenic Hirsutella strains (Mycota: Clavicipitaceous anamorphs). J. Invertebr. Pathol. 2008, 97, 103–113. [Google Scholar] [CrossRef]
- Bałazy, S.; Miętkiewski, R.; Tkaczuk, C.; Wegensteiner, R.; Wrzosek, M. Diversity of acaropathogenic fungi in Poland and other European countries. Exp. Appl. Acarol. 2008, 46, 53–70. [Google Scholar] [CrossRef]
- Szajdak, L.W.; Österberg, R. Amino acids present in humic acids from soils under different cultivations. Environ. Int. 1996, 22, 331–336. [Google Scholar] [CrossRef]
- Roberts, D.W.; Gupta, S.; St. Leger, R.J. Metabolite production by entomopathogenic fungi. Pes. Agropec. Bras. Brasilia 1992, 27, 325–347. [Google Scholar]
- Cole, R.J.; Jarvis, B.B.; Schweikert, M.A. Handbook of Secondary Fungal Metabolites; Academic Press: New York, NY, USA; Elsevier Science: New York, NY, USA, 2003; p. 2498. [Google Scholar]
- Elmholt, S. Mycotoxins in the Soil Environment. In Secondary Metabolites in Soil Ecology; Karlovsky, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 141–167. [Google Scholar]
- Zeilinger, S.; Garcìa-Estrada, C.; Martìn, J.F. Fungal secondary metabolites in “OMICS” era. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Zeilinger, S., Martin, J.F., Garcia-Estrada, C., Eds.; Springer: New York, NY, USA, 2015; pp. 1–12. [Google Scholar]
- Paszkiewicz, M.; Tyma, M.; Ligęza-Żuber, M.; Włóka, E.; Boguś, M.I.; Stepnowski, P. Mycotoxin production by entomopathogenic fungus Conidiobolus coronatus. Int. J. Environ. Agric. Res. 2017, 3, 33–40. [Google Scholar]
- Hoffmann, K.; Pawłowska, J.; Walther, G.; Wrzosek, M.; De Hoog, G.; Benny, G.; Kirk, P.; Voigt, K. The family structure of the Mucorales: A synoptic revision based on comprehensive multigene-genealogies. Persoonia 2013, 30, 57–76. [Google Scholar] [CrossRef] [Green Version]
- Koval, E.Z. Opredelited Entomofilnych Gribov SSSR; Naukova Dumka: Kiev, Ukraine, 1970; p. 260. (In Russian) [Google Scholar]
- Clark, R.V. Influence of some nitrogen and vitamin sources on growth, sporulation, and pathogenicity of Cochliobolus sativus. Can. J. Bot. 1971, 49, 2175–2186. [Google Scholar] [CrossRef]
- Nolan, R.A. A simplified, defined medium for growth of Entomophaga aulicae protoplasts. Can. J. Microbiol. 1988, 34, 45–51. [Google Scholar] [CrossRef]
- Grace, T.D.C. Establishment of Four Strains of Cells from Insect Tissues Grown In Vitro. Nature 1962, 195, 788–789. [Google Scholar] [CrossRef] [PubMed]
- Charnley, A.K. Host invasion by insect pathogenic fungi. In Proceedings of the VIth International Colloquium on Invertebrate Pathology and Microbial Control, Montpellier, France, 28 August–2 September 1994; Volume 1, pp. 31–37. [Google Scholar]
- Smith, R.J.; Grula, E. Toxic components on the larval surface of the corn earworm (Heliothis zea) and their effects on germination and growth of Beauveria bassiana. J. Invertebr. Pathol. 1982, 39, 15–22. [Google Scholar] [CrossRef]
- Bidochka, M.J.; Khachatourians, G.G. The implication of metabolic acids produced by Beauveria bassiana in pathogenesis of the migratory grasshopper, Melanoplus sanguinipes. J. Invertebr. Pathol. 1991, 58, 106–117. [Google Scholar] [CrossRef]
- Bidochka, M.J.; Khachatourians, G.G. Growth of the entomopathogenic fungus Beauveria bassiana on cuticular components from the migratory grasshopper, Melanoplus sanguinipes. J. Invertebr. Pathol. 1992, 59, 165–173. [Google Scholar] [CrossRef]
- Bidochka, M.J.; Khachatourians, G.G. Basic Proteases of Entomopathogenic Fungi Differ in Their Adsorption Properties to Insect Cuticle. J. Invertebr. Pathol. 1994, 64, 26–32. [Google Scholar] [CrossRef]
- Majchrowicz, I. Lipolytic activity of seven isolates of entomopathogenic fungi. Committee of Plant protection under auspices of Polish Academy of Science. In Proceedings of the Conference on Actual and Potential Use of Biological Pest Control On Plants, Skierniewice, Poland, 22–23 November 1995; pp. 100–103. [Google Scholar]
- Lunkenbein, S.; Takenberg, M.; Nimtz, M.; Berger, R.G. Characterization of a hydrophobin of the ascomycete Paecilomyces farinosus. J. Basic Microbiol. 2011, 51, 404–414. [Google Scholar] [CrossRef]
- Hong, I.-P.; Nam, S.-H.; Sung, G.-B.; Chung, I.-M.; Hur, H.; Lee, M.-W.; Kim, M.-K.; Guo, S.-X. Chemical Components of Paecilomyces tenuipes (Peck) Samson. Mycobiology 2007, 35, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Moorthi, V.; Balasubramanian, P.C.; Avery, P.B.; Selvarani, S.; Kubendran, T.; Rathinakumar, T.; Banu, A.N. Pathogenicity and proteome production of Isaria fumosorosea (=Paecilomyces Fumosoroseus) (WISE) isolates against lemon butterfly, Papilio demoleus (Papilionidae: Lepidoptera). Afr. J. Biotechnol. 2014, 13, 4176–4182. [Google Scholar] [CrossRef] [Green Version]
- Phadatare, S.U.; Srinivasan, M.C.; Deshpande, V.V. Evidence for controlled autoproteolysis of alkaline protease. A mechanism for physiological regulation of conidial discharge in Conidiobolus coronatus. JBIC J. Biol. Inorg. Chem. 1992, 205, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Putri, S.P.; Kinoshita, H.; Ihara, F.; Igarashi, Y.; Nihira, T. Farinomalein, a Maleimide-Bearing Compound from the Entomopathogenic Fungus Paecilomyces farinosus. J. Nat. Prod. 2009, 72, 1544–1546. [Google Scholar] [CrossRef]
- Miles, W.H.; Yan, M. Synthesis of farinomalein. Tetrahedron Lett. 2010, 51, 1710–1712. [Google Scholar] [CrossRef]
- El Amrani, M.; Debbab, A.; Aly, A.H.; Wray, V.; Dobretsov, S.; Müller, W.E.; Lin, W.; Lai, D.; Proksch, P. Farinomalein derivatives from an unidentified endophytic fungus isolated from the mangrove plant Avicennia marina. Tetrahedron Lett. 2012, 53, 6721–6724. [Google Scholar] [CrossRef]
- Xu, J.; Lacoske, M.H.; Theodorakis, E.A. Neurotrophic Natural Products: Chemistry and Biology. Angew. Chem. Int. Ed. Engl. 2014, 53, 956–987. [Google Scholar] [CrossRef] [Green Version]
- King, B.C. Species diversity and production of antimicrobial compounds by Pacific Northwestern Clavicipitalean Entomogenous Fungi (Cordyceps spp.). Master’s Thesis, University of British Columbia, Vancouver, BC, Canada, 2006. [Google Scholar]
- Elavarasi, A.; Peninal, S.; Priya, R.S.; Kalaiselvam, M. Cytotoxic and antibacterial activities of endophytic fungi isolated from mangroves. World J. Pharm. Res. 2018, 7, 954–961. [Google Scholar] [CrossRef]
- Shima, A.; Fukushima, K.; Arai, T.; Terada, H. Dual inhibitory effects of the peptide antibiotics leucinostatins on oxidative phosphorylation in mitochondria. Cell Struct. Funct. 1990, 15, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Che, J.-H.; Yun, J.-W.; Cho, E.-Y.; Kim, S.-H.; Kim, Y.-S.; Kim, W.H.; Park, J.-H.; Son, W.-C.; Kim, M.-K.; Kang, B.-C. Toxicologic assessment of Paecilomyces tenuipes in rats: Renal toxicity and mutagenic potential. Regul. Toxicol. Pharmacol. 2014, 70, 527–534. [Google Scholar] [CrossRef]
- Asai, T.; Chung, Y.-M.; Sakurai, H.; Ozeki, T.; Chang, F.-R.; Yamashita, K.; Oshima, Y. Tenuipyrone, a Novel Skeletal Polyketide from the Entomopathogenic Fungus, Isaria tenuipes, Cultivated in the Presence of Epigenetic Modifiers. Org. Lett. 2012, 14, 513–515. [Google Scholar] [CrossRef]
- Nam, K.S.; Jo, Y.S.; Kim, Y.H.; Hyun, J.W.; Kim, H.W. Cytotoxic activities of acetoxyscirpenediol and ergosterol peroxide from Paecilomyces tenuipes. Life Sci. 2001, 69, 229–237. [Google Scholar] [CrossRef]
- Evaluation Report ERC2011-07. Paecilomyces fumosoroseus Strain FE 9901; Health Canada Pest Management Regulatory Agency: Ottawa, ON, Canada, 2011.
- Liu, L.; Zhang, J.; Chen, C.; Teng, J.; Wang, C.; Luo, D. Structure and biosynthesis of fumosorinone, a new protein tyrosine phosphatase 1B inhibitor firstly isolated from the entomogenous fungus Isaria fumosorosea. Fungal Genet. Biol. 2015, 81, 191–200. [Google Scholar] [CrossRef]
- Sood, S.; Sandhu, S.S.; Mukherjee, T.K. Pharmacological and Therapeutic Potential of Beauvericin: A Short Review. J. Proteom. Bioinform. 2017, 10, 18–23. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the risks to human and animal health related to the presence of beauvericin and enniantins in food and feed. Eur. Food Saf. Auth. J. 2014, 12, 3802. [Google Scholar]
- Zhang, J.; Meng, L.-L.; Wei, J.-J.; Fan, P.; Liu, S.-S.; Yuan, W.-Y.; Zhao, Y.; Luo, D. PTP1B Inhibitors from the Entomogenous Fungi Isaria fumosorosea. Molecules 2017, 22, 2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Dong, T. Non-ribosomal peptides from entomogenous fungi. In Biocontrol of Lepidopteran Pests. Use of Soil Microbes and Their Metabolites; Sree, K.S., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 169–206. [Google Scholar] [CrossRef]
- Watanabe, N.; Hattori, M.; Yokoyama, E.; Isomura, S.; Ujita, M.; Hara, A. Entomogenous fungi that produce 2,6-pyridine dicarboxylic acid (dipicolinic acid). J. Biosci. Bioeng. 2006, 102, 365–368. [Google Scholar] [CrossRef]
- Dipicolinic Acid. Jubilant Organosys Limited. India. Uttar Pradesh. Product Categories 499-83-2. Available online: https://www.google.com/search?client=firefox-b-d&q=Dipicolinic+acid.+Jubilant+Organosys+Limited.+India (accessed on 2 October 2020).
- Veselovsky, V.V.; Danilov, L.L.; Maltsev, S.D.; Pronin, A.V.; Narovlyansky, A.N.; Sanin, A.V.; Deyeva, A.V.; Amchenkova, A.M. Dicarboxylic Germanium Complex and Its Therapeutic Use. U.S. Patent 6,204,276, 20 March 2001. [Google Scholar]
- Baral, B. Entomopathogenicity and Biological Attributes of Himalayan Treasured Fungus Ophiocordyceps sinensis (Yarsagumba). J. Fungi 2017, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Yue, K.; Ye, M.; Zhou, Z.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2012, 65, 474–493. [Google Scholar] [CrossRef]
- Firáková, S.; Proksa, B.; Sturdíkova, M. Biosynthesis and biological activity of enniatins. Die Pharm. 2007, 62, 563–568. [Google Scholar]
- Boucias, D.G.; Pendland, J.C. Priciples of Insect Pathology; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 1–537. [Google Scholar]
- Jirakkakul, J.; Punya, J.; Pongpattanakitshote, S.; Paungmoung, P.; Vorapreeda, N.; Tachaleat, A.; Klomnara, C.; Tanticharoen, M.; Cheevadhanarak, S. Identification of the nonribosomal peptide synthetase gene responsible for bassianolide synthesis in wood-decaying fungus Xylaria sp. BCC1067. Microbiology 2008, 154, 995–1006. [Google Scholar] [CrossRef] [Green Version]
- Czogalla, A. Oral cyclosporine A—The current picture of its liposomal and other delivery systems. Cell. Mol. Biol. Lett. 2009, 14, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Datta, R. Hydroxycarboxylic Acids. In Encyclopedia of Chemical Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2004. [Google Scholar] [CrossRef]
- Ishidoh, K.-I.; Kinoshita, H.; Igarashi, Y.; Ihara, F.; Nihira, T. Cyclic lipodepsipeptides verlamelin A and B, isolated from entomopathogenic fungus Lecanicillium sp. J. Antibiot. 2014, 67, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Bycroft, B.W.; Higton, A.A.; Roberts, A.D. Dictionary of Antibiotics and Related Substances; Chapman and Hall: London, UK, 1988. [Google Scholar]
- Demirci, S.; Göksu, S.; Boztas, M.; Tümer, F.; Secen, H. Synthesis of vertilecanin C and two new derivatives of vertilecanin A via nicotinic acid. Turk. J. Chem. 2008, 32, 287–295. [Google Scholar]
- Soman, A.G.; Gloer, J.B.; Angawi, R.F.; Wicklow, D.T.; Dowd, P.F. Vertilecanins: New Phenopicolinic Acid Analogues from Verticillium lecanii. J. Nat. Prod. 2001, 64, 189–192. [Google Scholar] [CrossRef]
- Tumer, F.; Goksu, S.; Secen, H. First sysnthesis of (±)-vertilecanin A. Russ. Chem. Bull. (Int. Ed.) 2005, 54, 2466–2467. [Google Scholar] [CrossRef]
- Gindin, G.; Barash, I.; Harari, N.; Raccah, B. Effect of endotoxic compounds isolated from Verticillium lecanii on the sweetpotato whitefly, Bemisia tabaci. Phytoparasitica 1994, 22, 189–196. [Google Scholar] [CrossRef]
- World Health Organization. Environmental Health Criteria 219, Fumonisin B1; World Health Organization: Geneva, Switzerland, 2000; p. 168. [Google Scholar]
- De Souza Diniz, S.P.S. Mycotoxins, Biochemical Approach; Albatroz: Rio de Janeiro, Brazil, 2016; p. 188. [Google Scholar]
- Fan, J.-Q.; Chen, X.-R.; Hu, Q.-B. Effects of Destruxin A on Hemocytes Morphology of Bombyx mori. J. Integr. Agric. 2013, 12, 1042–1048. [Google Scholar] [CrossRef]
- Fung, F.; Clark, R.F. Health Effects of Mycotoxins: A Toxicological Overview. J. Toxicol. Clin. Toxicol. 2004, 42, 217–234. [Google Scholar] [CrossRef]
- Cummings, N.J. Entomopathogenic Fungi in New Zealand Native Forests: The Genera Beauveria and Isaria. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2009. [Google Scholar]



{kind=link}
{kind=link}
| Amino Acid | Amount Per 1 L (g) |
|---|---|
| Cysteic acid | 234 |
| Taurine | 156.4 |
| Phosphoetanoloamine | 176.38 |
| Aspargig acid | 166.37 |
| Hydroxyproline | 163.9 |
| Threonine | 148.87 |
| Serine | 131.37 |
| Asparagine hydrate | 187.7 |
| Glutamic acd | 184 |
| Glutamine | 182.7 |
| α-amino adipic acid | 201.5 |
| Proline | 143.87 |
| Glycine | 93.87 |
| Alanine | 111.38 |
| Citruline | 219 |
| α-aminobutyric acid | 128.9 |
| Valine | 146.37 |
| Cystine | 150.19 |
| Methionine | 186.5 |
| Cystationine | 277.86 |
| Isoleucine | 164 |
| Leucine | 164 |
| Norleucine | 131.2 |
| Tyrosine | 226.5 |
| Phenylalanine | 203.13 |
| β-alanine | 111.4 |
| β-aminoisobutyric acid | 128.9 |
| γ-aminobutyric acid | 128.9 |
| Ornithine | 210.75 |
| Lysine | 228.37 |
| 1-methyl histidine | 234 |
| 3-methyl histidine | 211.5 |
| Tryptophane | 255.3 |
| Arginine | 217.8 |
| 1 | Lithium citrate buffer at pH = 2.9: 9.4000 citric acid, 9.3880 g lithium chloride, 2.3000 g lithium citrate, 2 mL 30% Brij 35®, 0.1 mL caprylic acid, 2.5 mL thiodiglycol (Pierce Chemical Co., Dallas, TX, USA) |
| 2 | Lithium citrate buffer at pH = 3.1: 9.7000 g citric acid, 9.5700 g lithium chloride, 3.9000 g lithium citrate 2 mL 30% Brij 35®, 0.1 mL caprylic acid, 2.5 mL tiodiglycol (Pierce Chemical Co., USA); |
| 3 | Lithium citrate buffer at pH = 3.35: 10.4980 g citric acid, 17.5000 g lithium chloride, 5.6500 g lithium citrate, 2 mL 30% Brij 35®, 0.1 mL caprylic acid, 2.5 mL tiodigliycol (Pierce Chemical Co., USA); |
| 4 | Lithium citrate buffer at pH = 4.05: 9.5000 g citric acid, 10.0000 g lithium chloride, 15.4500 g lithium citrate, 2 mL 30% Brij 35®, 0.1 mL caprylic acid, 2.5 mL tiodiglikolu (Pierce Chemical Co., USA); |
| 5 | Lithium citrate buffer at pH = 4.9: 8.5000 g citric acid, 39.9500 g lithium chloride, 50.6300 g lithium citrate, 2 mL 30% Brij 35®, 0.1 mL caprylic acid. |
| Amino Acids | Isaria farinosa | Isaria tenuipes | Isaria fumosorosea | Lecanicillium lecanii | Conidiobolus coronatus | Isaria coleopterorum |
|---|---|---|---|---|---|---|
| Cysteic acid | 67.6 ± 2.5 1.3 | 32.4 ± 1.2, 0.2 | 27.4 ± 1.0 0.3 | 14.6 ± 0.5 0.2 | 24.8 ± 0.9 6.4 | 17.3 ± 0.6 2.5 |
| Taurine | 28.7 ± 1.0 0.5 | 141.3 ± 5.3 0.9 | 131.0 ± 4.9 1.3 | 49.9 ± 1.6 0.5 | 1.7 ± 0.1 0.4 | 1.7 ± 0.1 0.3 |
| Phosphoethanolamine | - | - | - | - | 37.9 ± 1.4 9.7 | 37.8 ± 1.2 5.5 |
| Aspargic acid | 32.0 ± 1.2 0.6 | 757.9 ± 28.5 5.1 | 558.0 ± 21.0 5.5 | 441.5 ± 16.6 4.7 | 15.8 ± 0.6 4.1 | 45.5 ± 1.7 6.7 |
| Hydroksyproline | 1196.9 ± 44.9 22.5 | 322.4 ± 12.1 2.2 | 110.5 ± 4.2 1.1 | 139.9 ± 5.3 2.1 | 30.7 ± 1.2 7.9 | 46.0 ± 1.7 6.7 |
| Threonine | - | 705.0 ± 26.5 4.7 | 380.0 ± 14.3 3.6 | 445.0 ± 16.7 4.7 | 26.4 ± 1.0 6.8 | 24.1 ± 0.9 3.5 |
| Serine | 26.6 ± 1.0 0.5 | 865 ± 32.6 5.8 | 523.21 ± 9.7 4.9 | 530.21 ± 9.9 5.7 | 5.7 ± 0.2 1.5 | 21.0 ± 0.8 3.1 |
| Glutamic acid | 294.4 ± 11.0 5.5 | 194.4 ± 7.3 1.3 | 210.0 ± 7.9 2.0 | 111.1 ± 4.0 1.2 | - | - |
| Proline | 210.2 ± 7.7 3.9 | 604.4 ± 20.8 4.0 | 614.4 ± 23.1 5.8 | 496.2 ± 18.5 5.3 | 38.2 ± 1.4 9.8 | 15.9 ± 0.6 2.3 |
| Glycine | 389.7 ± 14.7 7.33 | 1190.4 ± 44.8 7.9 | 801.6 ± 30.1 7.6 | 599.1 ± 22.5 6.4 | 19.1 ± 0.7 4.9 | 53.8 ± 2.0 7.9 |
| Alanine | 450.7 ± 16.9 8.5 | 1270 ± 46.2 8.5 | 1475.9 ± 55.5 14.0 | 912.6 ± 34.3 9.7 | 15.3 ± 0.6 3.9 | 28.4 ± 1.1 4.2 |
| α-aminobutyric acid | - | 1280.6 ± 45.9 8.7 | 863.7 ± 32.5 8.2 | 945.9 ± 35.5 10.1 | 19.0 ± 0.7 4.9 | 28.0 ± 1.2 4.1 |
| Valine | 453.3 ± 17.0 8.5 | 1046.0 ± 39.2 6.9 | 718 ± 27.0 6.8 | 777.5 ± 29.2 8.3 | 10.3 ± 0.4 2.7 | 15.8 ± 0.6 2.3 |
| Cysteine | 233.7 ± 8.8 4.4 | - | - | - | - | - |
| Methionine | 65.9 ± 2.5 1.2 | - | - | - | - | - |
| Cystatathionine | 175.6 ± 6.6 3.3 | 465.9 ± 17.5 3.1 | 352.3 ± 13.2 3.3 | 346.0 ± 13.0 3.7 | 5.7 ± 0.2 1.5 | 13.6 ± 0.5 1.9 |
| Isoleucine | 457.6 ± 17.2 8.6 | 1391.7 ± 52.3 9.3 | 968.7 ± 36.4 9.2 | 1000.8 ± 35.8 10.7 | 23.9 ± 0.9 6.1 | 75.0 ± 2.8 10.9 |
| Leucine | 30.6 ± 1.2 0.6 | 52.2 ± 1.9 0.4 | 54.3 ± 2.0 0.5 | 18.3 ± 0.7 0.2 | 5.7 ± 0.2 1.5 | - |
| Tyrosine | 206.1 ± 7.8 3.9 | 603.3 ± 22.7 4.0 | 427.5 ± 16.1 4.1 | 467.0 ± 17.6 4.9 | 16.8 ± 0.6 4.3 | 42.1 ± 1.9 6.2 |
| β-alanine | 8.3 ± 0.3 0.2 | - | - | - | - | - |
| β-aminobutyric acid | 35.1 ± 1.3 0.7 | - | - | - | - | - |
| γ-aminobutyric acid | 516.4 ± 19.4 9.7 | 53.6 ± 2.0 0.4 | 38.7 ± 1.5 0.4 | 41.3 ± 1.5 0.4 | 3.6 ± 0.1 0.9 | 0.9 ± 0.03 0.1 |
| Ornithine | 21.5 ± 0.8 0.4 | 17.6 ± 0.7 0.1 | 16.0 ± 0.6 0.2 | - | - | - |
| Lysine | 194.4 ± 7.3 3.7 | 937.3 ± 35.2 6.3 | 485.7 ± 18.3 4.6 | 412.7 ± 15.5 4.4 | 14.8 ± 0.6 3.8 | 30.6 ± 1.2 4.5 |
| Histidine | 69.7 ± 2.6 1.3 | 232.8 ± 8.8 1.6 | 83.9 ± 3.2 0.8 | 53.0 ± 1.9 0.5 | 4.1 ± 0.2 1.1 | 4.1 ± 0.2 0.6 |
| 1-methylohistidine | - | 447.9 ± 16.8 3.0 | 150.9 ± 5.6 1.4 | 173.4 ± 6.5 1.9 | 9.0 ± 0.3 2.3 | 25.7 ± 1.0 3.9 |
| 3-methylohistidine | 32.2 ± 1.2 0.6 | 587.2 ± 22.1 3.9 | 407 ± 15.3 3.9 | 318.0 ± 11.0 3.4 | 7.9 ± 0.3 2.0 | 28.5 ± 1.1 4.2 |
| Arginine | 121.7 ± 4.6 2.3 | 1762.4 ± 66.2 11.8 | 1129.3 ± 42.3 10.7 | 1095.6 ± 41.2 11.7 | 53.6 ± 2.0 13.7 | 127.1 ± 4.8 18.6 |
| Total amount | 5317.9 | 14,963.0 | 10,530.41 | 9390.51 | 390.0 | 683.9 |
| Fungus Name | Amino Acids | |||||||
|---|---|---|---|---|---|---|---|---|
| Sulfuric | Acidic | Neutral | Basic | |||||
| (µg g−1d.m.) | (%) | (µg g−1d.m.) | (%) | (µg g−1d.m.) | (%) | (µg g−1d.m.) | (%) | |
| Isaria farinose | 510.6 | 4.9 | 1781.3 | 16.9 | 5963.5 | 56.6 | 2271.4 | 21.6 |
| Isaria tenuipes | 410.5 | 4.4 | 1667.6 | 17.8 | 5259.6 | 56.0 | 2052.8 | 21.9 |
| Isaria fumosorosea | 639.5 | 4.3 | 2845.3 | 19.0 | 7492.7 | 50.1 | 3985.2 | 26.6 |
| Lecanicillium lecanii | 32.2 | 8.3 | 116.6 | 29.9 | 151.9 | 38.9 | 89.4 | 22.9 |
| Conidiobolus coronatus | 32.7 | 4.8 | 174.4 | 25.5 | 259.7 | 37.9 | 216.9 | 31.7 |
| Isaria coleopterorum | 571.4 | 10.8 | 1255.5 | 23.6 | 3051.5 | 57.4 | 439.5 | 8.3 |
| Fungus Name | Amino Acids | |||
|---|---|---|---|---|
| Hydrophilic | Hydrophobic | |||
| (µg g−1s.m.) | (%) | (µg g−1s.m.) | (%) | |
| Isaria farinose | 4214.0 | 40.0 | 6315.8 | 59.9 |
| Isaria tenuipes | 3784.9 | 40.3 | 5605.6 | 59.7 |
| Isaria fumosorosea | 7004.3 | 46.8 | 7958.7 | 53.2 |
| Lecanicillium lecanii | 232.3 | 59.6 | 157.6 | 40.4 |
| Conidiobolus coronatus | 410.4 | 60.0 | 273.3 | 39.7 |
| Isaria coleopterorum | 1791.2 | 33.68 | 3526.7 | 66.32 |
| Amino Acids | Isaria farinose | Isaria tenuipes | Isaria fumosorosea | Lecanicillium lecanii | Conidiobolus coronatus | Isaria coleopterorum | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Our Study | [51] | Our Study | [52] | Our Study | [53] | Our Study | [49] | Our Study | [54] | Our Study | |
| Alanine | + | + | + | + | + | + | + | + | + | + | |
| β-alanine | + | ||||||||||
| α-aminobutyric acid | + | + | + | + | + | ||||||
| β-aminobutyric acid | + | + | |||||||||
| γ-aminobutyric acid | + | + | + | + | + | + | |||||
| Arginine | + | + | + | + | + | + | + | + | |||
| Asparagine | + | + | + | ||||||||
| Aspartic acid | + | + | + | + | + | + | + | ||||
| Cystathionine | + | + | + | + | + | + | |||||
| Cysteic acid | + | + | + | + | + | + | |||||
| Cysteine | + | + | |||||||||
| Glutamic acid | + | + | + | + | + | + | + | ||||
| Glutamine | + | ||||||||||
| Glycine | + | + | + | + | + | + | + | + | + | ||
| Histidine | + | + | + | + | + | + | + | + | + | + | |
| Hydroksyproline | + | + | + | + | + | + | |||||
| Isoleucine | + | + | + | + | + | + | + | + | + | ||
| Leucine | + | + | + | + | + | + | + | + | |||
| Lysine | + | + | + | + | + | + | + | + | + | + | |
| Methionine | + | + | + | + | |||||||
| 1-methylohistidine | + | + | + | + | + | ||||||
| 3-methylohistidine | + | + | + | + | + | + | |||||
| Ornithine | + | + | + | ||||||||
| Phenylalanine | + | + | + | ||||||||
| Phosphoethanolamine | + | + | |||||||||
| Proline | + | + | + | + | + | + | + | + | + | + | |
| Serine | + | + | + | + | + | + | + | + | + | + | |
| Taurine | + | + | + | + | + | + | |||||
| Threonine | + | + | + | + | + | + | + | + | |||
| Tryptophan | + | ||||||||||
| Tyrosine | + | + | + | + | + | + | + | + | + | ||
| Valine | + | + | + | + | + | + | + | + | + | ||
| Fungi | Compounds Molecular Formula (MF), Molecular Weight (MW) | Physiological Activity IC50—The Half Maximal Inhibitory Concentration, LD50—Lethal Dose, 50% |
|---|---|---|
| 1. Isaria farinosa | Farinomalein MF C10H13NO4, MW 211.21 g mol−1 | The first natural maleimide, pesticide of Phytophthora stem rot in soybean [55], activity comparable to antibiotic amphotericin B for inhibition Phytophthora sojae [56], selective inhibition of Phytophthora sojae with an MIC (Minimal Inhibitory Concentration) value of 5 µg disk−1 [18], IC50 = 4.4 µg mL−1 [55,57] |
| Farinosones A, B, C A-MF C25H27NO4, MW 405.49 g mol−1 B-MF C25H26NO5, MW 420.48 g mol−1 C-MF C19H25NO5, MW 347.41 g mol−1 | Classe as polyketide, cytotoxic in the PC-12 cell line [20], pyridone alkaloid, neurotrophic activities [58], antimicrobial, antitumor, IC50 = 1.1 µg mL−1 [59] | |
| Paecilosetin MF C22H31NO4, MW 373.49 g mol−1 | Tetramic acid derivative, antibacterial, ant-leucemia (P388 cell line) [59] antimicrobial, antitumor, IC50 = 3.2 µg mL−1 [60] | |
| Leucinostatin A MF C62H111N11O13, MW 1218.61 g mol−1 | Linear peptides, paecilotoxins, LD50 = 1.8 mg kg−1-mice, antimicrobial, anticancer [35], antibiotic [61] | |
| 2. Isaria tenuipes | Extract | LD50 = >2000 mg/kg of body weight, LD50 = >5 g kg−1 of body weight in rats and dogs [62] |
| Tenuipyrone MF C15H16O6, MW 292.28 g mol−1 | Polyketide with an unprecedented tetracyclic ring system bearing a spiroketal structural component [20,63] | |
| Acetoxyscirpenediol MF C17H24O6, MW 324.37 g mol−1 | Trichothecenes, cytotoxic, antitumor [59], metabolite of anguidine is to inhibit protein synthesis in rabbit reticulocytes, anguidine is antitumor agent which was in the process of phase II evaluation in gastrointestinal malignancy, central nervous system tumors, colorectal adenocarcinoma [64]) | |
| Spirotenuipesine A, B A-MF C15H22O4, MW 266.34 g mol−1 B-MF C15H22O5, MW 282.33 g mol−1 | Trichothecenes, neurotrophic factor biosynthesis [59] | |
| 3. Isaria fumosorosea | Extract (beauverolide 1, beauverolide 5, 2.6-pyridinedicarboxylic) | LD50 > 0.9 × 108 CFU/animal-pulmonary, LD50 > 108 CFU/animal-oral [65] |
| Fumosorinone MF C29H35NO5, MW 477.59 g mol−1 | Classe as polyketide, inhibits tyrosine phosphatase 1B (PTP1B) to treat type 2 diabetes mellitus (T2DM) [20], 2-pyridone alkaloid, insecticidal, IC50 = 14.04 IM [66] | |
| Beauvericin MF C45H57N3O9, MW 783.95 g mol−1 | Cytotoxic, cyclic peptide, insecticidal against mosquito larvae and blowflies, antiplasmodial [35,59], antibiotic, cyclohexadepsipeptide, anthermintic, antibacterial, antifungal, antimycobacterial and anticancer activities [39], herbicidal, antiretroviral, cytotoxic, antihaptotactic, anti-cholesterol, chemosensitizer, as well as repression of amyloid plaque formation in Alzheimer′s disease [67], LD50 = 100 mg kg−1-mice [68], IC50 = 0.59 µM [69], LC50 of 633 ppm (95% fiducial limits, 530–748 ppm) and LC90 of 1196 ppm (95% fiducial limits, 954–1863 ppm) [35] | |
| Beauverolide MF C29H45N3O5, MW 515.69 g mol−1 | Cyclic tetradepsipeptide [59], inhibits the adhesion, extension, and phagocytosis of plasmatocyte of Galleria mellonella [70] | |
| Pyridine-2,6-dicarboxylic acid (dipicolinic acid) MF C7H5NO4, MW 167.19 g mol−1 | Insecticidal [71], produces pharmaceuticals and metal salts for the application of nutritional supplements, act a chelating agent and an enzyme inhibitor [72], LD50 = 20 µg mice−1 intramuscularly on the day of infection, LD50 = 10.50 g kg−1 of body weight [73] | |
| Cepharosporolides C, E, F E-MF C10H14O4, MW 198.22 g mol−1 | Biosynthetic precursor of tenuipyrone, and cephalosporolide F [63,69] | |
| 2-carboxymethyl-4-(30-hydroxybutyl)furan | Insecticidal [74], biological and pharmacological activity, chemotherapeutic agents, IC50 = >1000 (µM) [69,75] | |
| Enniatins A, B, C (fusafungine) A, C-MF C36H63N3O9, MW 681.90 g mol−1 B-MF C34H59N3O9, MW 653.85 g mol−1 | Microbial, a cyclodepsipeptide alkali metal ionophore inhibitor of acyl-CoA: cholesterol acyltransferase (ACAT), antitumor (Sigma), antibiotic activity, toxins in low non-toxic concentrations are tested for application in human and veterinary medicine, phytotoxic, herbicide, IC50 = 10−4 M effect on seed germination of the parasitic weed Striga hermonthica [35,76], LD50 = 350 mg kg−1-mice [68] | |
| 4. Lecanicillium lecanii | Bassianolide MF C48H84N4O12, MW 909.20 g mol−1 | Classe as nonribosomal peptide, insecticidal [20], toxic to lepidopteran larvae after infection or feeding [77], biological activities for cryptophycins, didemnins, dolastatins, PF1022, enniatin, destruxin, beauvericin, valinomycin and anti-plasmodial, antimycobacterial, antitumor activities, toxicity against bacteria: Staphylococcus aureus, Bacillus subtilis, Escherichia coli, Pseudomonas aeruginosa [78] |
| Cyclosporine MF C62H111N11O12, MW 1202.61 g mol−1 | Cyclic peptide, immunosuppressant, insecticidal (mosquito, but no Galleria mellonella) [38,59,79], restaining rejection following solid organ transplantations especially heart, lung, kidney, preventing and treating graft-versus-host disease after bone marrow transplants, the treatment of numerous autoimmune diseases, anti-inflammatory, anti-parasitic (anti-malaria), antifungal, antiviral (ant-HIV) [79], LD50 = 2329 mg kg−1-mice, 1480 mg kg−1-rats, >1000 mg kg−1-rabbits (DrugBank online) | |
| Pyridine-2,6-dicarboxylic acid (dipicolinic acid) | Look at Isaria fumosorosea [59,71] | |
| Hydroxycarboxilic acid | Group of chemicals use in processes such as biodegradable plastics for consumer products and nontoxic and easily degradable solvents, cleaning agents, plasticizers, etc., global commercial [59,80] | |
| Verlamelin A,B B-MF C44H69N7O11, MW 872.06 g mol−1 | Cyclic hexadepsipeptide lipopeptide, exhibits antifungal activity against plant pathogenic fungi [81], drugs [81,82] | |
| Vertilecanin A, B, C A-MF C13H11NO3, MW 229.21 g mol−1 | Phenopicolinic acid derivatives were synthesized from nicotinic acid, antibacterial activity against Bacillus subtilis [83,84], insecticidal activity against Helicoverpa zea [85] | |
| Phospholipids toxic | Insecticidal [86] | |
| 5. Conidiobolus coronatus | Fumonisin B1 MF C34H59NO15, MW 721.83 g mol−1 | Bioinsecticide [39] causes leukoencephalomalacia in horses, mycotoxin, hepatotoxic (Sigma), nephrotoxicity, immunotoxicity, reproductive toxicity, embryotoxicity, teratogenicity, mutagenicity, carcinogenicity, phytotoxic, nephrotoxic, pulmonary, oedema syndrome, hepatocarcinogenic, esophageal cancer [87], LD50 = 1.25 mg kg−1 [88] |
| Destruxin A MF C29H47N5O7, MW 577.71 g mol−1 | Cyclic depsipeptides consisting of 5 amino acids and D-α-hydroxy acid, insecticidal [35], antitumor, herbicidal, cytoxicity [20], bioinsecticide [39,89], phytotoxin, cancer cell, anti-immunity of Metarhizium anisopliae antimicrobial peptides [89], virustatic against, LD50 = 13.2–16.9 mg/kg-mice [3] | |
| Enniatin A, B (fusafungine) A-MF C36H63N3O9, MW 681.90 g mol−1 B-MF C33H57N3O9, MW 639.00 g mol−1 | Look at Isaria fumosorosea | |
| Beauvericin | Look at Isaria fumosorosea | |
| T-2 and HT-2 toxins metabolites from trichothecene group T-2-MF C24H34O9, MW 466.52 g mol−1 HT-2-MF C22H32O8, MW 424.49 g mol−1 trichothecene MF C15H22O2, MW 234.16 g mol−1 | Bioinsecticide [39], mycotoxin, antibacterial, antiviral, antifungal, cytostatic activity, anorexia, hematuria, leukocytosis, leukopenia, inhibitors of protein and DNA synthesis, natural intoxications of animals and man [36], sesquiterpenoid chemicals characterized by a tetracyclic 12,13-epoxytrichothecene, LD50 = 0.5–300 mg kg−1-animal, LD50 = 13–140 µg kg−1-rat brain [90] | |
| 6. Isaria coleopterorum | Pathogens of Lepidoptera and Coleoptera [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szajdak, L.W.; Bałazy, S.; Meysner, T. Amino Acids in Entomopathogenic Fungi Cultured In Vitro. Agronomy 2020, 10, 1899. https://doi.org/10.3390/agronomy10121899
Szajdak LW, Bałazy S, Meysner T. Amino Acids in Entomopathogenic Fungi Cultured In Vitro. Agronomy. 2020; 10(12):1899. https://doi.org/10.3390/agronomy10121899
Chicago/Turabian StyleSzajdak, Lech Wojciech, Stanisław Bałazy, and Teresa Meysner. 2020. "Amino Acids in Entomopathogenic Fungi Cultured In Vitro" Agronomy 10, no. 12: 1899. https://doi.org/10.3390/agronomy10121899
APA StyleSzajdak, L. W., Bałazy, S., & Meysner, T. (2020). Amino Acids in Entomopathogenic Fungi Cultured In Vitro. Agronomy, 10(12), 1899. https://doi.org/10.3390/agronomy10121899