Genome-Wide Identification and Expression Analysis of MAPK and MAPKK Gene Family in Pomegranate (Punica Granatum L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Data Sources
2.2. Identification of PgMAPKs and PgMAPKKs in Pomegranate
2.3. Phylogenetic Relationship and Conserved Motif Analysis
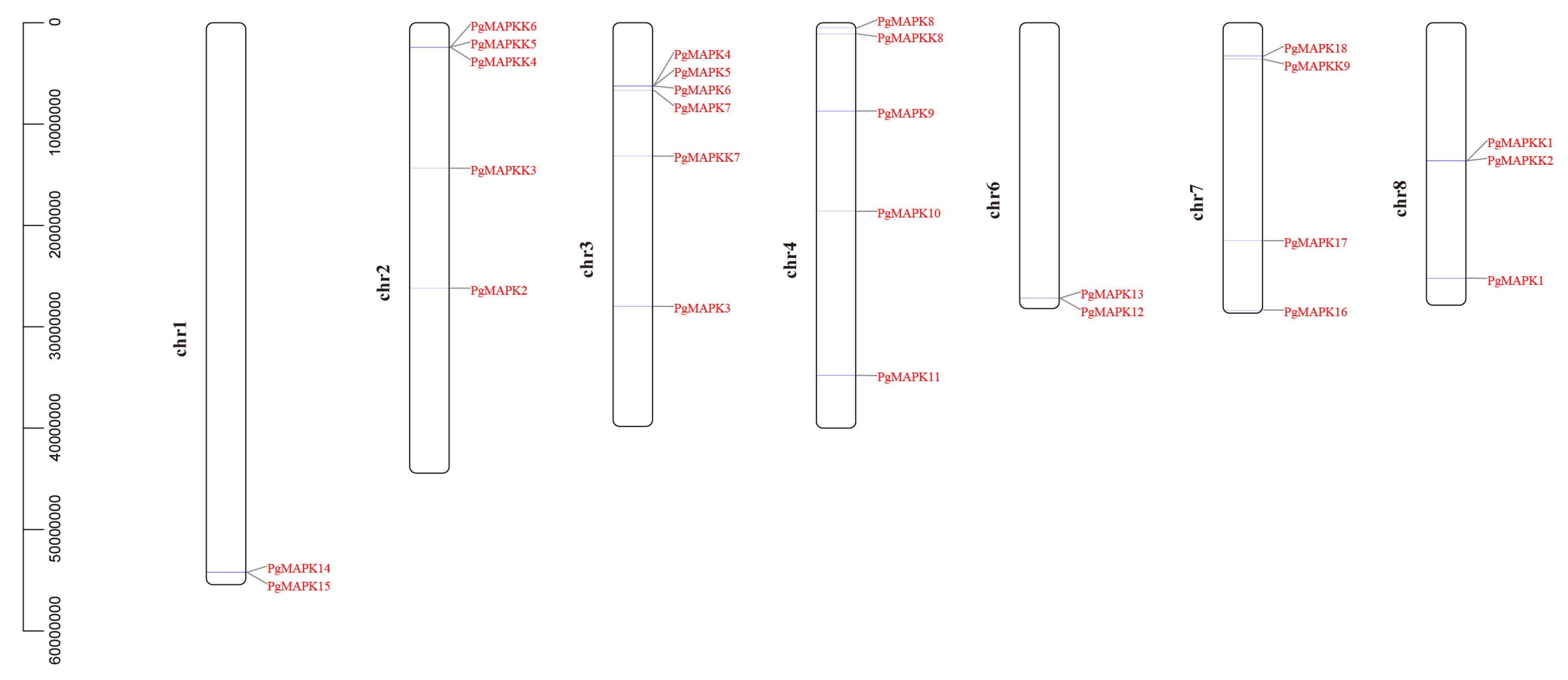
2.4. Chromosome Localization and Expression Profiles
3. Results
3.1. Identification and Sequence Analysis of PgMAPKs and PgMAPKKs
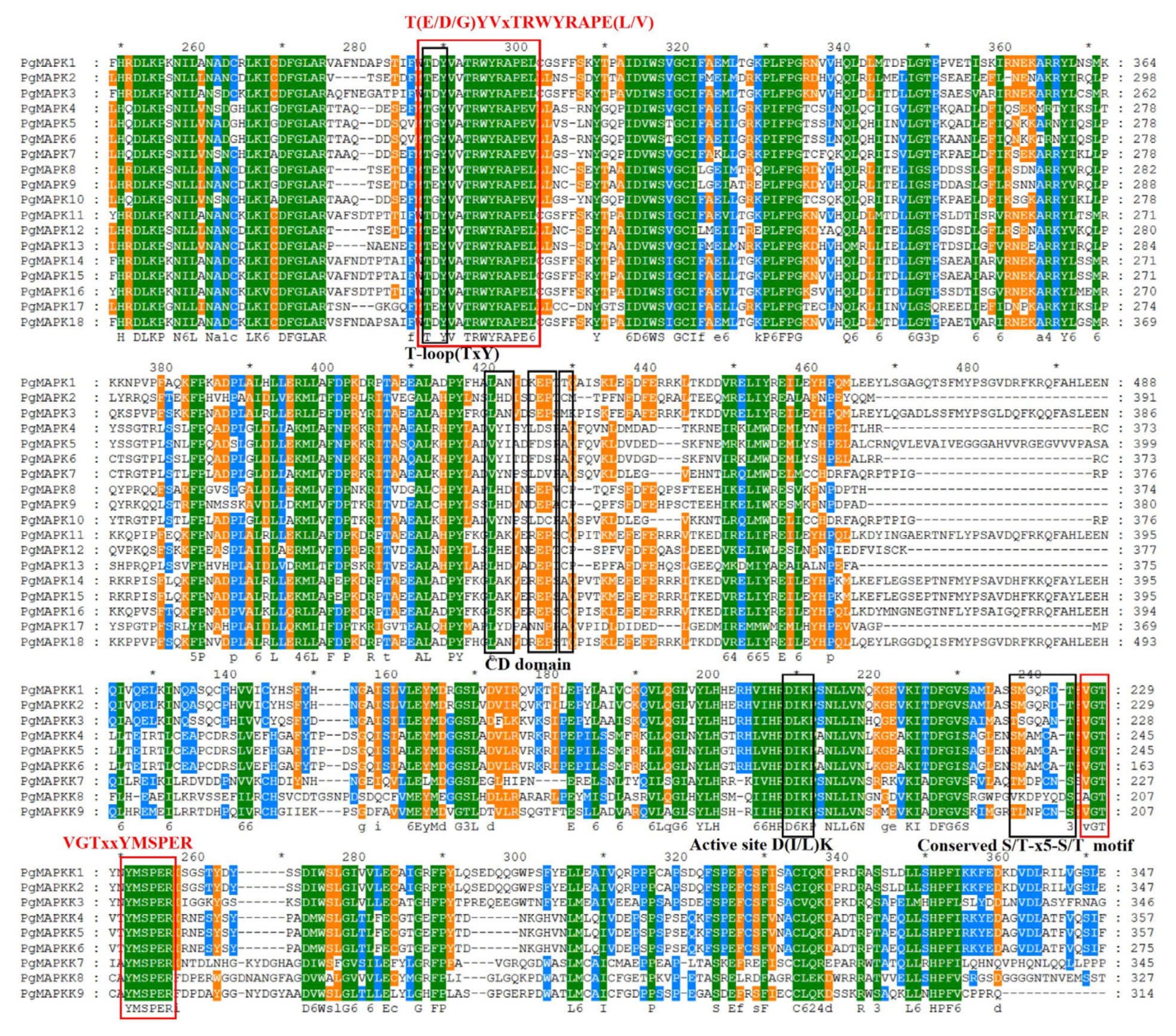
3.2. Conserved Motif Analysis of PgMAPK and PgMAPKK Gene Family Members
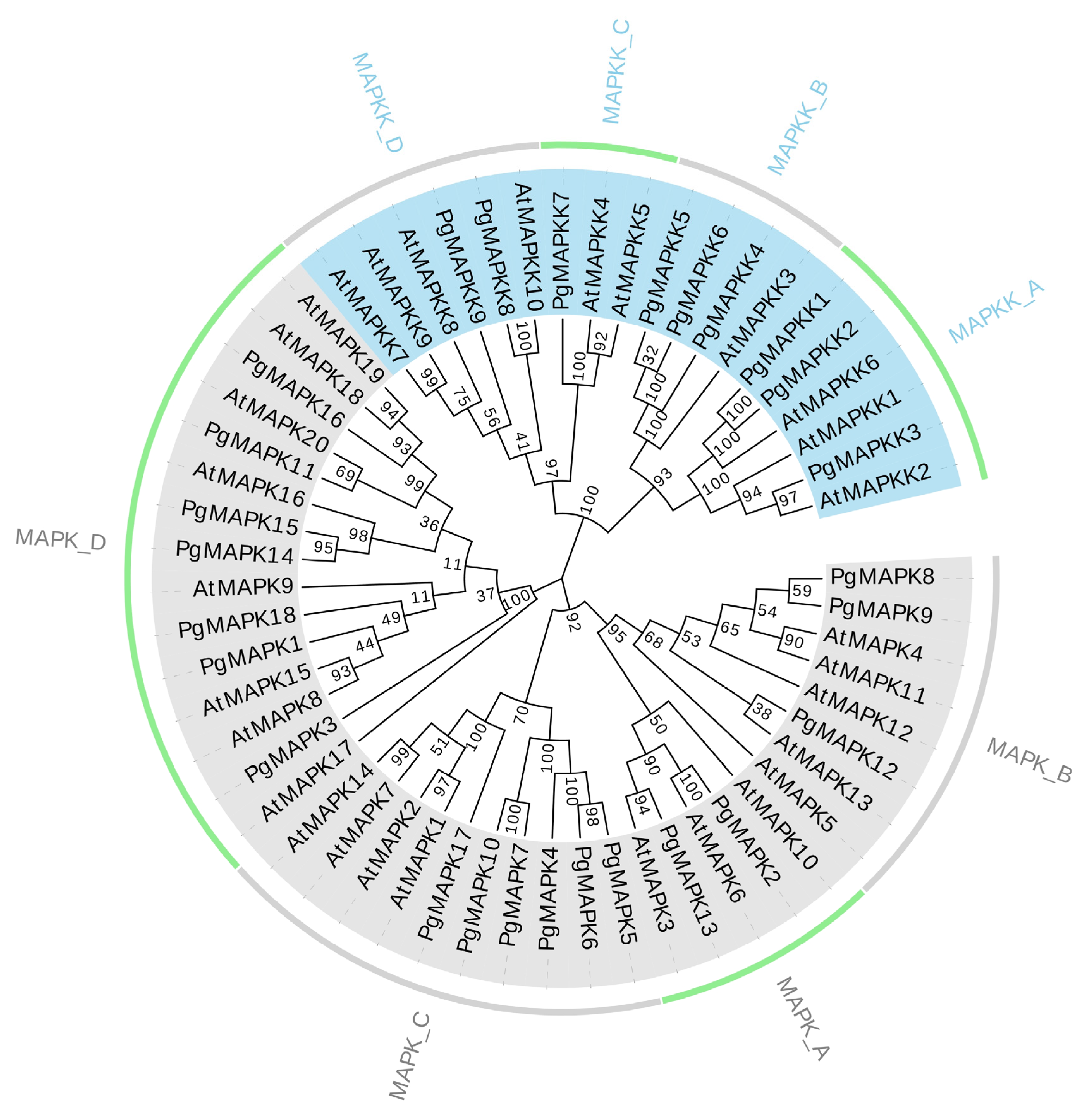
3.3. Phylogenetic Relationship of PgMAPKs and PgMAPKKs
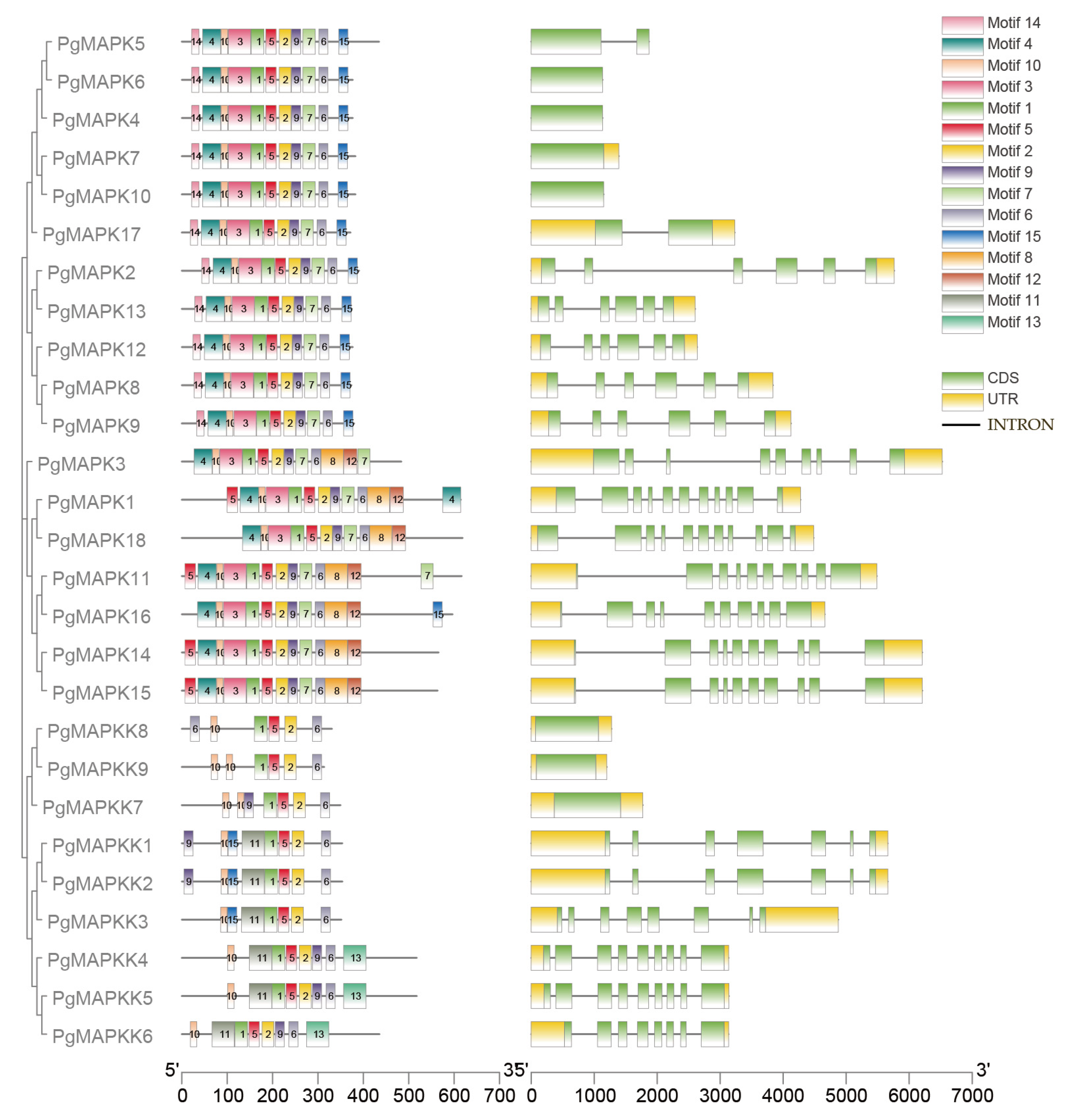
3.4. Gene Structure Analysis of PgMAPKs and PgMAPKKs
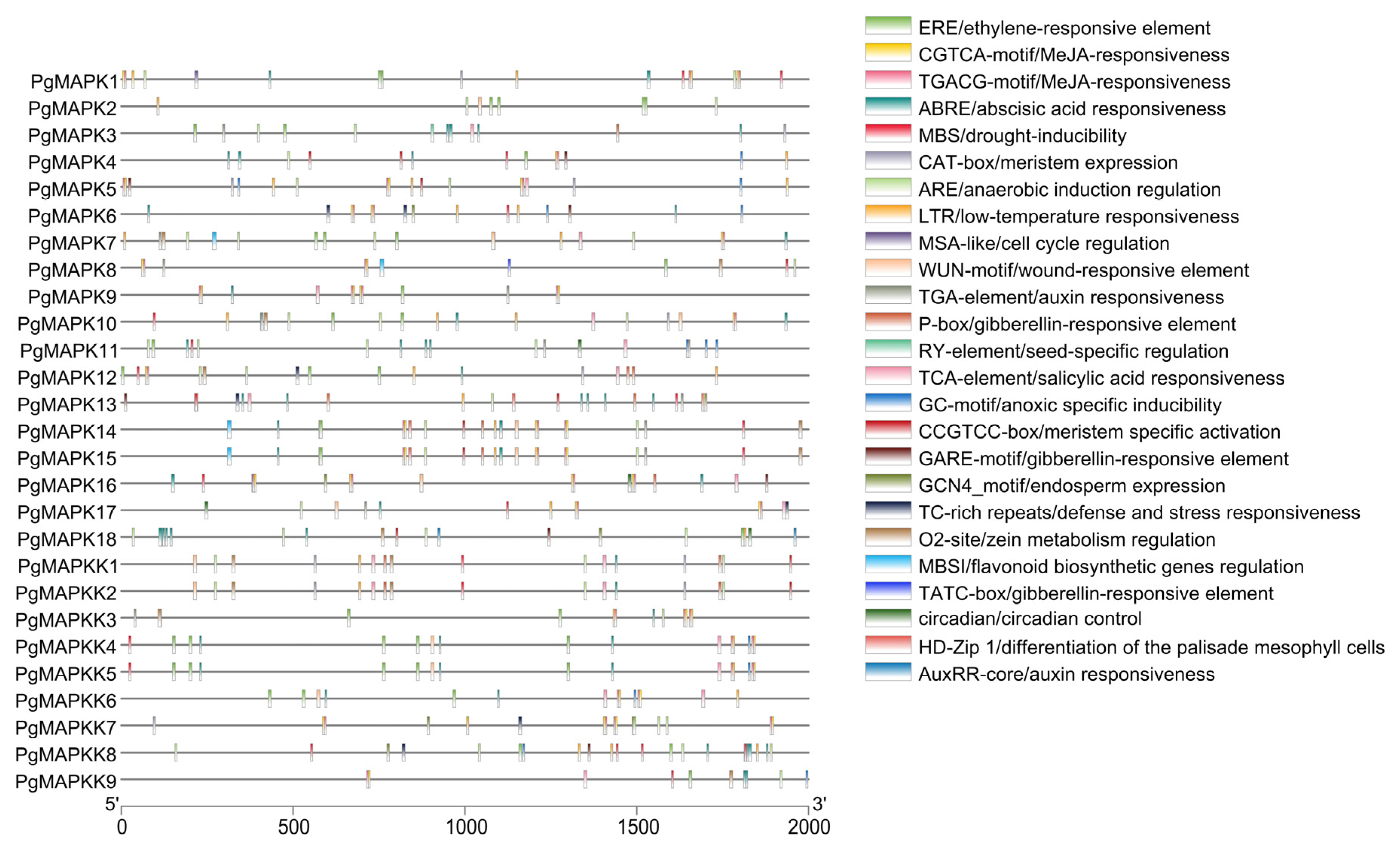
3.5. Cis-Acting Element Prediction of PgMAPKs and PgMAPKKs
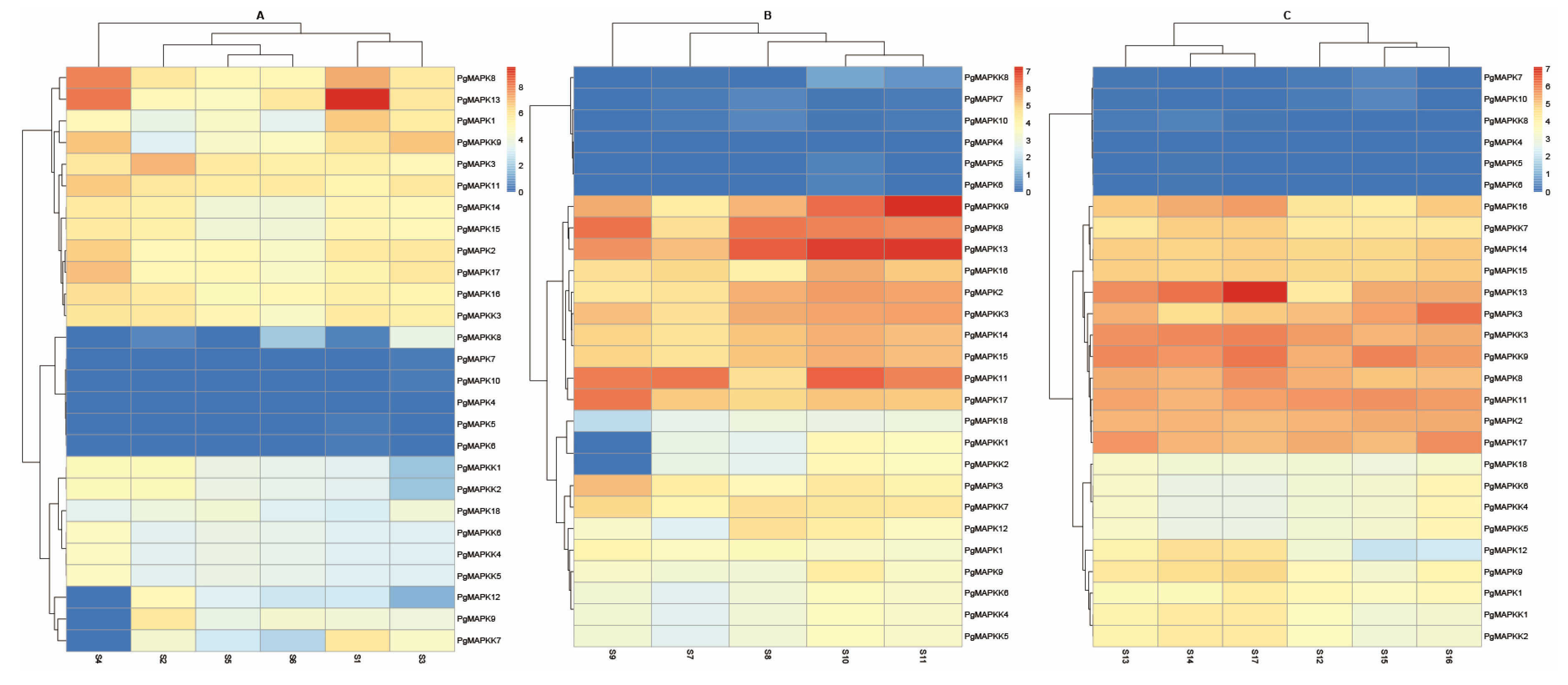
3.6. Expression Pattern of PgMAPKs and PgMAPKKs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nakagami, H.; Pitzschke, A.; Hirt, H. Emerging MAP kinase pathways in plant stress signalling. Trends Plant Sci. 2005, 10, 339–346. [Google Scholar] [CrossRef]
- Zhan, H.; Hong, Y.; Zhao, X.; Wang, M.; Weining, S.; Nie, X. Genome-wide identification and analysis of MAPK and MAPKK gene families in bread wheat (Triticum aestivum L.). Genes 2017, 8, 10. [Google Scholar] [CrossRef]
- Shen, H.; Liu, C.; Zhang, Y.; Meng, X.; Zhou, X.; Chu, C.; Wang, X. OsWRKY30 is activated by MAP kinases to confer drought tolerance in rice. Plant Mol. Biol. 2012, 80, 241–253. [Google Scholar] [CrossRef]
- Raina, S.K.; Wankhede, D.P.; Jaggi, M.; Singh, P.; Jalmi, S.K.; Raghuram, B.; Sheikh, A.H.; Sinha, A.K. CrMPK3, a mitogen activated protein kinase from Catharanthus roseusand its possible role in stress induced biosynthesis of monoterpenoid indole alkaloids. BMC Plant Biol. 2012, 12, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tena, G.; Asai, T.; Chiu, W.; Sheen, J. Plant mitogen-activated protein kinase signaling cascades. Curr. Opin. Plant Biol. 2001, 4, 392–400. [Google Scholar] [CrossRef]
- Zhou, H.; Ren, S.; Han, Y.; Zhang, Q.; Qin, L.; Xing, Y. Identification and analysis of mitogen-activated protein kinase (MAPK) cascades in Fragaria vesca. Int. J. Mol. Sci. 2017, 18, 1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Pan, J.; Zhang, D.; Jiang, S.; Cai, G.; Wang, L.; Li, D. Identification of mitogen-activated protein kinase kinase gene family and MKK–MAPK interaction network in maize. Biochem. Biophys. Res. Commun. 2013, 441, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Jonak, C.; Ökrész, L.; Bögre, L.; Hirt, H. Complexity, cross talk and integration of plant MAP kinase signalling. Curr. Opin. Plant Biol. 2002, 5, 415–424. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, Y.; Yang, T.; Zhang, L.; Xu, S.; Xue, L.; An, L. Diverse signals converge at MAPK cascades in plant. Plant Physiol. Biochem. 2006, 44, 274–283. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, L.; Xue, C.; Fang, H.; Zhao, J.; Liu, M. Genome-wide identification and analysis of MAPK and MAPKK gene family in Chinese jujube (Ziziphus jujuba Mill.). BMC Gen. 2017, 18, 855. [Google Scholar] [CrossRef] [Green Version]
- Kazuya, I.; Ichimura, K.; Shinozaki, K.; Tena, G.; Sheen, J.; Henry, Y.; Champion, A.; Kreis, M.; Zhang, S.; Hirt, H.; et al. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar] [CrossRef]
- Moustafa, K.; AbuQamar, S.; Jarrar, M.; Al-Rajab, A.J.; Trémouillaux-Guiller, J. MAPK cascades and major abiotic stresses. Plant Cell Rep. 2014, 33, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Andreasson, E.; Ellis, B. Convergence and specificity in the Arabidopsis MAPK nexus. Trends Plant Sci. 2010, 15, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.; Roux, M.; Petersen, M.; Mundy, J. MAP kinase cascades in Arabidopsis innate immunity. Front. Plant Sci. 2012, 3, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandhari, W.D.; Misra, M.; Singh, P.; Sinha, A.K.; Provart, N.J. Rice mitogen activated protein kinase Kinase and mitogen activated protein kinase interaction network revealed by in-silico docking and yeast two-hybrid approaches. PLoS ONE 2013, 8, e65011. [Google Scholar] [CrossRef] [Green Version]
- Rao, K.P.; Tambi, R.; Kundan, K.; Badmi, R.; Krishna, S.A. In silico analysis reveals 75 members of mitogen-activated protein kinase kinase kinase gene family in rice. DNA Res. 2010, 3, 139–153. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.; Liu, F.; Zou, J.; Xing, G.; Deng, P.; Song, W.; Tong, W.; Nie, X. Genome-wide identification, evolution, and co-expression network analysis of mitogen-activated protein kinase kinase kinases in Brachypodium distachyon. Front. Plant Sci. 2016, 7, 1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, D.; Wang, L.; Li, D. Genome-wide analysis of mitogen-activated protein kinase gene family in maize. Plant Mol. Biol. Rep. 2013, 31, 1446–1460. [Google Scholar] [CrossRef]
- Cui, L.; Yang, G.; Yan, J.; Pan, Y.; Nie, X. Genome-wide identification, expression profiles and regulatory network of MAPK cascade gene family in barley. BMC Gen. 2019, 20, 750. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Yang, K.Y.; Li, G.J.; Liu, Y.; Zhang, S. Activation of Ntf4, a tobacco mitogen-activated protein kinase, during plant defense response and its involvement in hypersensitive response-like cell death. Plant Physiol. 2006, 141, 1482–1493. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, K.; Casais, C.; Peck, S.C.; Shinozaki, K.; Shirasu, K. MEKK1 is required for MPK4 activation and regulates tissue-specific and temperature-dependent cell death in Arabidopsis. J. Biol. Chem. 2006, 281, 36969–36976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez-Rodriguez, M.C.; Adams-Phillips, L.; Liu, Y.; Wang, H.; Su, S.; Jester, P.J.; Zhang, S.; Bent, A.F.; Krysan, P.J. MEKK1 is required for flg22-induced MPK4 activation in Arabidopsis plants. Plant Physiol. 2007, 143, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef] [PubMed]
- De Zelicourt, A.; Colcombet, J.; Hirt, H. The role of MAPK modules and ABA during abiotic stress signaling. Trends Plant Sci. 2016, 21, 677–685. [Google Scholar] [CrossRef]
- Wang, J.; Ding, H.; Zhang, A.; Ma, F.; Cao, J.; Jiang, M. A novel mitogen-activated protein kinase gene in maize (Zea mays), ZmMPK3, is involved in response to diverse environmental cues. J. Integr. Plant Biol. 2010, 52, 442–452. [Google Scholar] [CrossRef]
- Xiong, L.; Yang, Y. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid-inducible mitogen-activated protein kinase. Plant Cell 2003, 15, 745–759. [Google Scholar] [CrossRef] [Green Version]
- Ghawana, S.; Kumar, S.; Ahuja, P.S. Early low-temperature responsive mitogen activated protein kinases RaMPK1 and RaMPK2 from Rheum australe D. Don respond differentially to diverse stresses. Mol. Biol. Rep. 2009, 37, 933. [Google Scholar] [CrossRef]
- Danquah, A.; de Zélicourt, A.; Boudsocq, M.; Neubauer, J.; Frei Dit Frey, N.; Leonhardt, N.; Pateyron, S.; Gwinner, F.; Tamby, J.; Ortiz-Masia, D.; et al. Identification and characterization of an ABA-activated MAP kinase cascade in Arabidopsis thaliana. Plant J. 2015, 82, 232–244. [Google Scholar] [CrossRef]
- Hahn, A.; Harter, K. Mitogen-activated protein kinase cascades and ethylene: Signaling, biosynthesis, or both? Plant Physiol. 2009, 149, 1207–1210. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.; Cho, Y.; Tena, G.; Xiong, Y.; Sheen, J. Dual control of nuclear EIN3 by bifurcate MAPK cascades in C2H4 signaling. Nature 2008, 451, 789–795. [Google Scholar] [CrossRef]
- Harel-Beja, R.; Sherman, A.; Rubinstein, M.; Eshed, R.; Bar-Ya Akov, I.; Trainin, T.; Ophir, R.; Holland, D. A novel genetic map of pomegranate based on transcript markers enriched with QTLs for fruit quality traits. Tree Genet. Genomes 2015, 11, 109. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, Y.; Zhao, X.; Wang, J.; Gu, M.; Yuan, Z. Transcriptomic profiling of pomegranate provides insights into salt tolerance. Agronomy 2020, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Qin, G.; Liu, C.; Li, J.; Qi, Y.; Gao, Z.; Zhang, X.; Yi, X.; Pan, H.; Ming, R.; Xu, Y. Diversity of metabolite accumulation patterns in inner and outer seed coats of pomegranate: Exploring their relationship with genetic mechanisms of seed coat development. Hortic. Res. 2020, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, M.; Zhao, X.; Zhou, J.; Huo, Y.; Ding, Y.; Yuan, Z. The complete chloroplast genomes of Punica granatum and a comparison with other species in lythraceae. Int. J. Mol. Sci. 2019, 20, 2886. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Fang, Y.; Zhang, T.; Fei, Z.; Han, F.; Liu, C.; Liu, M.; Xiao, W.; Zhang, W.; Wu, S.; et al. The pomegranate (Punica granatum L.) genome provides insights into fruit quality and ovule developmental biology. Plant Biotechnol. J. 2018, 16, 1363–1374. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yuan, Z.; Feng, L.; Fang, Y. Cloning and expression of anthocyanin biosynthetic genes in red and white pomegranate. J. Plant Res. 2015, 128, 687–696. [Google Scholar] [CrossRef]
- Thomma, B.P.; Penninckx, I.A.; Cammue, B.P.; Broekaert, W.F. The complexity of disease signaling in Arabidopsis. Curr. Opin. Immunol. 2001, 13, 63–68. [Google Scholar] [CrossRef]
- Luo, X.; Li, H.; Wu, Z.; Yao, W.; Zhao, P.; Cao, D.; Yu, H.; Li, K.; Poudel, K.; Zhao, D.; et al. The pomegranate (Punica granatum L.) draft genome dissects genetic divergence between soft- and hard-seeded cultivars. Plant Biotechnol. J. 2019, 18, 955–968. [Google Scholar] [CrossRef] [Green Version]
- Kersey, P.J.; Allen, J.E.; Allot, A.; Barba, M.; Boddu, S.; Bolt, B.J.; Carvalho-Silva, D.; Christensen, M.; Davis, P.; Grabmueller, C.; et al. Ensembl Genomes 2018: An integrated omics infrastructure for non-vertebrate species. Nucleic Acids Res. 2017, 46, D802–D808. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Birney, E.; Cerruti, L.; Ettwiller, L.; Eddy, S.; Griffiths-Jones, S.; Howe, K.; Marshall, M.; Sonnhammer, E. The pfam protein families database. Nucleic Acids Res. 2002, 30, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; Mcgettigan, P.; McWilliam, H.; Valentin, F.; Wallace, I.; Wilm, A.; López, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, K.B.; Nicholas, H.; Deerfield, D. GeneDoc: Analysis and visualization of genetic variation. Embnew. News 1996, 4, 14. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Magali, L.; Patrice, D.; Gert, T.; Kathleen, M.; Yves, M.; Yves, V.D.P.; Pierre, R.; Stephane, R. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Racine, J.S. RStudio: A platform-independent IDE for R and sweave. J. Appl. Econ. 2012, 27, 167–172. [Google Scholar] [CrossRef]
- Ophir, R.; Sherman, A.; Rubinstein, M.; Eshed, R.; Sharabi-Schwager, M.; Harel Beja, R.; Bar-Ya’Akov, I.; Holland, D. Single-nucleotide polymorphism markers from de-novo assembly of the pomegranate transcriptome reveal germplasm genetic diversity. PLoS ONE 2014, 9, e88998. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhang, J.; Li, H.; Niu, J.; Xue, H.; Liu, B.; Wang, Q.; Luo, X.; Zhang, F.; Zhao, D.; et al. Transcriptomic analysis reveals candidate genes for female sterility in pomegranate flowers. Front. Plant Sci. 2017, 8, 1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, N.N.; Britton, M.T.; Fass, J.N.; Nicolet, C.M.; Lin, D.; Tian, L. Exploring the transcriptome landscape of pomegranate fruit peel for natural product biosynthetic gene and SSR marker discovery. Integr. Plant Biol. 2011, 53, 800–813. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Lovato, A.; Polverari, A.; Wang, M.; Liang, Y.; Ma, Y.; Cheng, Z. Genome-wide identification and analysis of mitogen activated protein kinase kinase kinase gene family in grapevine (Vitis vinifera). BMC Plant Biol. 2014, 14, 219. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, W.; Tie, W.; Ding, Z.; Ding, X.; Liu, Y.; Yan, Y.; Wu, C.; Peng, M.; Xu, B.; et al. The MAPKKK and MAPKK gene families in banana: Identification, phylogeny and expression during development, ripening and abiotic stress. Sci. Rep. 2017, 7, 1159. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xu, R.; Luo, X.; Jiang, Z.; Shu, H. Genome-wide identification and expression analysis of MAPK and MAPKK gene family in Malus domestica. Gene 2013, 531, 377–387. [Google Scholar] [CrossRef]
- Bohnert, H.; Nelson, D.; Jensen, R. Adaptations to environmental stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef]
- Nicole, M.; Hamel, L.; Morency, M.; Beaudoin, N.; Ellis, B.E.; Séguin, A. MAP-ping genomic organization and organ-specific expression profiles of poplar MAP kinases and MAP kinase kinases. BMC Gen. 2006, 7, 223. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Wang, J.; Cheng, L.; Liu, S.; Wu, J.; Peng, Z.; Lu, G. Genome-wide analysis of the mitogen-activated protein kinase gene family in Solanum lycopersicum. Gene 2012, 499, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Liu, X.; Long, D.; Guo, Q.; Fang, Y.; Bian, C.; Zhang, D.; Zeng, Q.; Xiang, Z.; Zhao, A. Molecular cloning and expression analysis of mulberry MAPK gene family. Plant Physiol. Biochem. 2014, 77, 108–116. [Google Scholar] [CrossRef]
- Zhang, S.; Klessig, D.F. MAPK cascades in plant defense signaling. Trends Plant Sci. 2001, 6, 520–527. [Google Scholar] [CrossRef]
- Kovtun, Y.; Chiu, W.; Zeng, W.; Sheen, J. Suppression of auxin signal transduction by a MAPK cascade in higher plants. Nature 1998, 395, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Shi, L.; Liu, Y.; Tang, Q.; Shen, L.; Sheng, Y.; Cai, J.; Yu, H.; Wang, R.; Wen, J.; et al. Genome-wide identification and transcriptional expression analysis of mitogen-activated protein kinase and mitogen-activated protein kinase kinase genes in Capsicum annuum. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Pan, C.; Wang, Y.; Ye, L.; Wu, J.; Chen, L.; Zou, T.; Lu, G. Genome-wide identification of MAPK, MAPKK, and MAPKKK gene families and transcriptional profiling analysis during development and stress response in cucumber. BMC Gen. 2015, 16, 386. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Li, D.; Dai, Y.; Liu, S.; Huang, L.; Hong, Y.; Zhang, H.; Song, F. Characterization, expression patterns and functional analysis of the MAPK and MAPKK genes in watermelon (Citrullus lanatus). BMC Plant Biol. 2015, 15, 298. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gong, M.; Guo, J.; Xin, H.; Gao, Y.; Liu, C.; Dai, D.; Tang, L. Genome-wide identification of Jatropha curcas MAPK, MAPKK, and MAPKKK gene families and their expression profile under cold stress. Sci. Rep. 2018, 8, 16163. [Google Scholar] [CrossRef] [Green Version]
- Holub, E. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. 2001, 2, 516–527. [Google Scholar] [CrossRef]
- Goyal, R.; Tulpan, D.; Chomistek, N.; González-Peña Fundora, D.; West, C.; Ellis, B.; Frick, M.; Laroche, A.; Foroud, N. Analysis of MAPK and MAPKK gene families in wheat and related Triticeae species. BMC Gen. 2018, 19, 178. [Google Scholar] [CrossRef] [Green Version]
- Seyfferth, C.; Tsuda, K. Salicylic acid signal transduction: The initiation of biosynthesis, perception and transcriptional reprogramming. Front. Plant Sci. 2014, 5, 697. [Google Scholar] [CrossRef] [Green Version]
- Shariatipour, N.; Heidari, B. Meta-analysis of expression of the stress tolerance associated genes and uncover their cis-regulatory elements in rice (Oryza sativa L.). Open Biol. J. 2020, 13, 39–49. [Google Scholar] [CrossRef]
- Itzhaki, H.; Maxson, J.M.; Woodson, W.R. An ethylene-responsive enhancer element is involved in the senescence-related expression of the carnation glutathione-S-transferase (GST1) gene. Proc. Natl. Acad. Sci. USA 1994, 91, 8925–8929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubler, F.; Jacobsen, J. Gibberellin-responsive elements in the promoter of a barley high-pl [alpha]-amylase gene. Plant Cell 1992, 4, 1435–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiltinan, M.J.; Marcotte, W.R., Jr.; Quatrano, R.S. A plant leucine zipper protein that recognizes an abscisic acid response element. Science 1990, 250, 267–271. [Google Scholar] [CrossRef] [Green Version]
- Goldsbrough, A.P.; Albrecht, H.; Stratford, R. Salicylic acid-inducible binding of a tobacco nuclear protein to a 10 bp sequence which is highly conserved amongst stress-inducible genes. Plant J. 1993, 3, 563–571. [Google Scholar] [CrossRef]
- Krysan, P.; Colcombet, J. Cellular complexity in MAPK signaling in plants: Questions and emerging tools to answer them. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Chardin, C.; Schenk, S.; Hirt, H.; Colcombet, J.; Krapp, A. Review: Mitogen-activated protein kinases in nutritional signaling in Arabidopsis. Plant Sci. 2017, 260. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Y.; Hu, F.; Zhang, S.Y.; Wang, K.; Zhang, C.R.; Liu, T. MAPKs regulate root growth by influencing auxin signaling and cell cycle-related gene expression in cadmium-stressed rice. Environ. Sci. Pollut. Res. 2013, 20, 5449–5460. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Cultivars | Sample Type | Library | Platform | Reference |
|---|---|---|---|---|---|
| SRR5279396 | “Dabenzi” | Root | Paired end | Illumina HiSeq 4000 | [33] |
| SRR5279397 | “Dabenzi” | Leaf | Paired end | Illumina HiSeq 4000 | [33] |
| SRR5279395 | “Dabenzi” | Flower | Paired end | Illumina HiSeq 4000 | [33] |
| SRR5279391 | “Dabenzi” | Inner seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [33] |
| SRR5279388 | “Dabenzi” | Outer seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [33] |
| SRR5279394 | “Dabenzi” | Pericarp (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [33] |
| SRR5678820 | “Tunisia” | Inner seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [33] |
| SRR5678819 | “Baiyushizi” | Inner seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [33] |
| SRR1055290 | “nana” | Mixed samples of leaves, flowers, fruit and roots | Single end | 454 GS FLX Titanium | [53] |
| SRR1054190 | “Black127” | Mixed samples of root, leaf, flower and fruit | Single end | 454 GS FLX Titanium | [53] |
| SRR5446598 | “Tunisia” | Functional male flower (3.0−5.0 mm) | Paired end | Illumina HiSeq 2500 | [54] |
| SRR5446595 | “Tunisia” | Functional male flower (5.1–13.0 mm) | Paired end | Illumina HiSeq 2500 | [54] |
| SRR5446592 | “Tunisia” | Functional male flower (13.1–25.0 mm) | Paired end | Illumina HiSeq 2500 | [54] |
| SRR5446607 | “Tunisia” | Female sterility flower (3.0–5.0 mm) | Paired end | Illumina HiSeq 2500 | [54] |
| SRR5446604 | “Tunisia” | Female sterility flower (5.1–13.0 mm) | Paired end | Illumina HiSeq 2500 | [54] |
| SRR5446601 | “Tunisia” | Female sterility flower (13.1–25.0 mm) | Paired end | Illumina HiSeq 2500 | [54] |
| SRR080723 | “Wonderful” | Pericarp | Paired end | Illumina HiSeq 2000 | [55] |
| Gene Name | Protein ID | Genomic Position | Size (aa) | MW (Da) | pIs | Types | Group |
|---|---|---|---|---|---|---|---|
| PgMAPK1 | XP_031372466.1 | chr8:25215947…25219536 | 615 | 68,952.19 | 7.69 | TDY | D |
| PgMAPK2 | XP_031382378.1 | chr2:26190661…26195979 | 391 | 44,872.36 | 5.57 | TEY | A |
| PgMAPK3 | XP_031384487.1 | chr3:27987357…27983062 | 484 | 55,597.84 | 8.79 | TDY | D |
| PgMAPK4 | XP_031387033.1 | chr3:6220495…6221628 | 377 | 42,844.83 | 8.90 | TGY | C |
| PgMAPK5 | XP_031387034.1 | chr3:6229201…6231072 | 435 | 48,521.45 | 7.66 | TGY | C |
| PgMAPK6 | XP_031387035.1 | chr3:6233547…6234680 | 377 | 42,552.40 | 9.23 | TGY | C |
| PgMAPK7 | XP_031388383.1 | chr3:6647361…6648512 | 383 | 43,266.50 | 9.26 | TGY | C |
| PgMAPK8 | XP_031389841.1 | chr4:494852…498050 | 374 | 42,897.79 | 6.11 | TEY | B |
| PgMAPK9 | XP_031389944.1 | chr4:8712267…8715865 | 380 | 43,399.50 | 6.20 | TEY | B |
| PgMAPK10 | XP_031392146.1 | chr4:18603577…18604728 | 383 | 43,164.35 | 9.36 | TGY | C |
| PgMAPK11 | XP_031392360.1 | chr4:34805345…34809858 | 617 | 70,420.74 | 9.19 | TDY | D |
| PgMAPK12 | XP_031399464.1 | chr6:27190634…27192917 | 377 | 43,002.99 | 4.94 | TEY | B |
| PgMAPK13 | XP_031400536.1 | chr6:27163862…27162051 | 375 | 42,968.25 | 5.78 | TEY | A |
| PgMAPK14 | XP_031402771.1 | chr1:54220111…54215519 | 566 | 64,371.77 | 8.76 | TDY | D |
| PgMAPK15 | XP_031402779.1 | chr1:54220111…54215513 | 564 | 64,243.64 | 8.76 | TDY | D |
| PgMAPK16 | XP_031404075.1 | chr7:28354650…28351083 | 597 | 68,130.90 | 9.35 | TDY | D |
| PgMAPK17 | XP_031404901.1 | chr7:21504171…21503431 | 372 | 42,781.79 | 6.92 | TEY | C |
| PgMAPK18 | XP_031406973.1 | chr7:3285226…3281535 | 619 | 69,387.45 | 8.06 | TDY | D |
| MAPKK1 | XP_031374856.1 | chr8:13616418…13612292 | 354 | 39,870.90 | 5.96 | DIK | A |
| MAPKK2 | XP_031374857.1 | chr8:13616418…13612292 | 354 | 39,870.90 | 5.96 | DIK | A |
| MAPKK3 | XP_031381620.1 | chr2:14357511…14354367 | 352 | 38,956.51 | 5.49 | DLK | A |
| MAPKK4 | XP_031383136.1 | chr2:2401332…2404194 | 518 | 58,182.26 | 5.76 | DIK | B |
| MAPKK5 | XP_031383137.1 | chr2:2401332…2404194 | 518 | 58,182.26 | 5.76 | DIK | B |
| MAPKK6 | XP_031383138.1 | chr2:2401664…2404194 | 436 | 49,232.33 | 6.04 | DIK | B |
| MAPKK7 | XP_031388194.1 | chr3:13155229…13156281 | 350 | 39,123.69 | 9.38 | DIK | C |
| MAPKK8 | XP_031393241.1 | chr4:1081579…1082574 | 331 | 36,995.33 | 8.31 | DIK | D |
| MAPKK9 | XP_031406836.1 | chr7:3593726…3594670 | 314 | 34,910.05 | 8.35 | DIK | D |
| Gene Family | Species | GroupA | GroupB | GroupC | GroupD | Total | Reference |
|---|---|---|---|---|---|---|---|
| MAPK | Pomegranate | 2 | 3 | 6 | 7 | 18 | |
| Chinese jujube | 2 | 1 | 2 | 5 | 11 | [10] | |
| Arabidopsis | 3 | 5 | 4 | 8 | 20 | [14] | |
| Rice | 2 | 1 | 2 | 10 | 15 | [15] | |
| Brachypodium distachyon | 2 | 2 | 3 | 9 | 16 | [17] | |
| Maize | 4 | 2 | 2 | 11 | 10 | [18] | |
| Apple | 5 | 6 | 5 | 10 | 26 | [58] | |
| Poplar | 4 | 4 | 4 | 9 | 21 | [60] | |
| Tomato | 3 | 4 | 2 | 7 | 16 | [61] | |
| Mulberry | 2 | 3 | 2 | 3 | 10 | [62] | |
| Bread wheat | 7 | 3 | 8 | 36 | 54 | [3] | |
| MAPKK | Pomegranate | 3 | 3 | 1 | 2 | 9 | |
| Chinese jujube | 2 | 1 | 0 | 2 | 5 | [10] | |
| Arabidopsis | 3 | 1 | 2 | 4 | 10 | [14] | |
| Rice | 2 | 1 | 2 | 3 | 8 | [15] | |
| Brachypodium distachyon | 2 | 3 | 2 | 5 | 12 | [17] | |
| Apple | 3 | 1 | 2 | 3 | 9 | [58] | |
| Poplar | 3 | 1 | 2 | 5 | 11 | [60] | |
| Bread wheat | 3 | 2 | 1 | 12 | 18 | [3] |
| Function | Promoter Name | Promoter Annotation | Total Number |
|---|---|---|---|
| Motifs related to stress response | ARE | cis-acting regulatory element essential for the anaerobic induction | 20 |
| LTR | cis-acting element involved in low-temperature responsiveness | 18 | |
| MBS | MYB binding site involved in drought-inducibility | 17 | |
| TC-rich repeats | cis-acting element involved in defense and stress responsiveness | 6 | |
| WUN-motif | wound-responsive element | 12 | |
| GC-motif | enhancer-like element involved in anoxic specific inducibility | 9 | |
| Motifs related to growth and development | CAT-box | cis-acting regulatory element related to meristem expression | 8 |
| GCN4_motif | cis-regulatory element involved in endosperm expression | 5 | |
| CCGTCC-box | cis-acting regulatory element related to meristem specific activation | 10 | |
| O2-site | cis-acting regulatory element involved in zein metabolism regulation | 11 | |
| HD-Zip 1 | element involved in differentiation of the palisade mesophyll cells | 4 | |
| RY-element | cis-acting element involved in seed-specific regulation | 1 | |
| MBSI | MYB binding site involved in flavonoid biosynthetic genes regulation | 5 | |
| MSA-like | cis-acting element involved in cell cycle regulation | 1 | |
| circadian | cis-acting regulatory element involved in circadian control | 5 | |
| Motifs related to hormone response | ABRE | cis-acting element involved in the abscisic acid responsiveness | 23 |
| AuxRR-core | cis-acting regulatory element involved in auxin responsiveness | 1 | |
| CGTCA-motif | cis-acting regulatory element involved in the MeJA-responsiveness | 20 | |
| TGACG-motif | cis-acting regulatory element involved in the MeJA-responsiveness | 19 | |
| ERE | ethylene-responsive element | 20 | |
| GARE-motif | gibberellin-responsive element | 7 | |
| P-box | gibberellin-responsive element | 8 | |
| TATC-box | cis-acting element involved in gibberellin-responsiveness | 1 | |
| TGA-element | auxin-responsive element | 11 | |
| TCA-element | cis-acting element involved in salicylic acid responsiveness | 15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Y.; Ge, D.; Dong, J.; Guo, L.; Yuan, Z. Genome-Wide Identification and Expression Analysis of MAPK and MAPKK Gene Family in Pomegranate (Punica Granatum L.). Agronomy 2020, 10, 1015. https://doi.org/10.3390/agronomy10071015
Ren Y, Ge D, Dong J, Guo L, Yuan Z. Genome-Wide Identification and Expression Analysis of MAPK and MAPKK Gene Family in Pomegranate (Punica Granatum L.). Agronomy. 2020; 10(7):1015. https://doi.org/10.3390/agronomy10071015
Chicago/Turabian StyleRen, Yuan, Dapeng Ge, Jianmei Dong, Linhui Guo, and Zhaohe Yuan. 2020. "Genome-Wide Identification and Expression Analysis of MAPK and MAPKK Gene Family in Pomegranate (Punica Granatum L.)" Agronomy 10, no. 7: 1015. https://doi.org/10.3390/agronomy10071015
APA StyleRen, Y., Ge, D., Dong, J., Guo, L., & Yuan, Z. (2020). Genome-Wide Identification and Expression Analysis of MAPK and MAPKK Gene Family in Pomegranate (Punica Granatum L.). Agronomy, 10(7), 1015. https://doi.org/10.3390/agronomy10071015