Introgression of Resistance to Leafminer (Liriomyza cicerina Rondani) from Cicer reticulatum Ladiz. to C. arietinum L. and Relationships between Potential Biochemical Selection Criteria


 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Description of the Location
2.3. Experiments, Agronomic Practices and Monitoring of Leafminer
2.4. Screening for Resistance to the Leafminer
2.5. Agro-Morphological Traits
2.6. Estimation of Heritability
2.7. Analyses of Organic Acids by High-Performance Liquid Chromatography (HPLC)
2.8. Statistical Analyses
3. Results
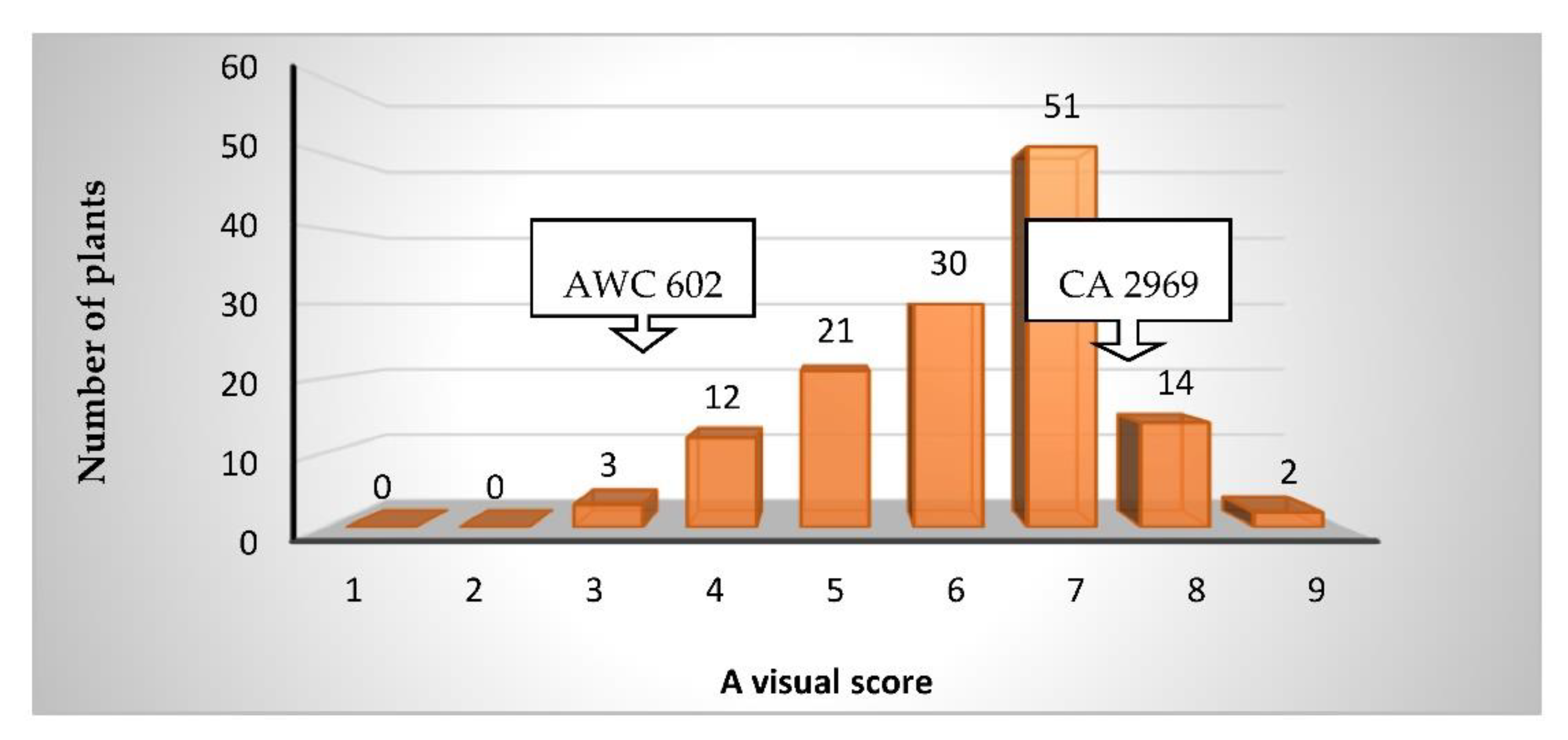
3.1. Resistance to the Leafminer
3.2. Agro-Morphological Traits
3.3. Relationships between Resistance to the Leafminer and Agro-Morphological Traits
3.4. Inheritance Pattern for Resistance to the Leafminer
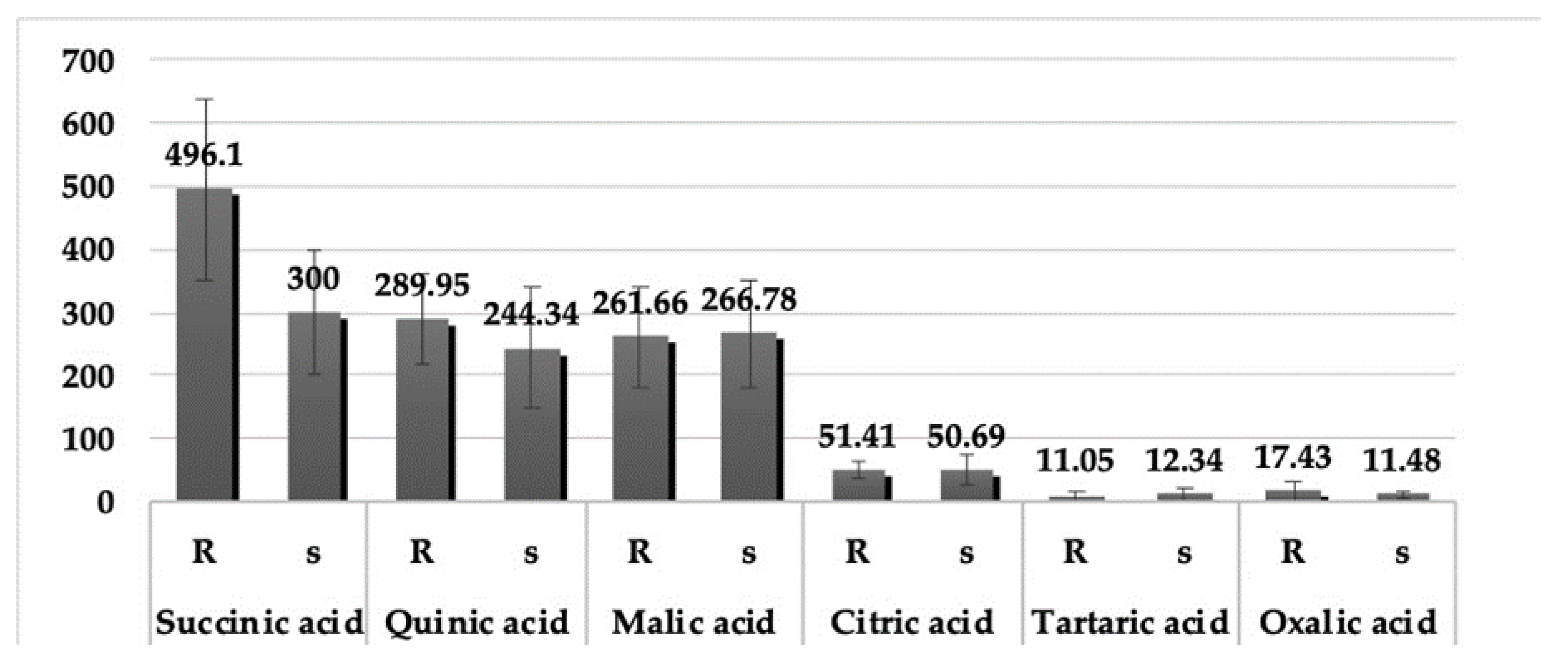
3.5. Organic Acids
3.6. Differences between Organic Acids in Resistant and Susceptible Genotypes of Chickpea
3.7. Organic Acids in Chickpea Genotype in Relation to Expression of Resistance to Leaf Miner
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, H.C.; Gowda, C.L.L.; Stevenson, P.C.; Ridsdill-Smith, T.J.; Clement, S.L.; Ranga Rao, G.V.; Romeis, J.; Miles, M.; El-Bouhssini, M. Host Plant Resistance and Insect Pest Management in Chickpea; Yadav, S.S., Redden, B., Chen, W., Sharma, B., Eds.; CAB International: Wallingford, Oxon, UK, 2007; pp. 520–537. [Google Scholar]
- Reed, W.; Caradona, C.; Sithanantham, S.; Lateef, S.S. Chickpea Insect Pest and Their Control. In The Chickpea; Saxena, M.C., Singh, K.B., Eds.; CAB International: Wallingford, Oxon, UK, 1987; pp. 283–318. [Google Scholar]
- Soltani, A.; Beyareslan, A.; Gencer, L.; Hamdi, S.H.; Bousselmi, A.; Amri, M.; Mediouni Ben Jemaa, J.M. Parasitoids of chickpea leaf miner Liriomyza cicerina (Diptera: Agromyzidae) and their parasitism rate on chickpea fields in North Tunisia. J. Asia Pac. Entomol. 2018, 21, 1215–1221. [Google Scholar] [CrossRef]
- Cikman, E.; Civelek, H.S.; Weintraub, P.G. The parasitoid complex of Liriomyza cicerina on chickpea (Cicer arietinum). Phytoparasitica 2008, 36, 211–216. [Google Scholar] [CrossRef]
- Weigand, S. Insects pest of chickpea in the Mediterranean area and possibilities for resistance. Cah. Options Mediterr. 1990, 9, 73–76. [Google Scholar]
- Toker, C.; Erler, F.; Çanci, H.; Ceylan, F.O. Severity of leaf miner (Liriomyza cicerina Rond.) damage in relation to leaf type in chickpea. Turk. J. Entomol. 2010, 34, 211–226. [Google Scholar]
- Singh, K.B.; Weigand, S. Registration of three leaf miner-resistant chickpea germplasm lines: ILC 3800, ILC 5901, and ILC 7738. Crop Sci. 1996, 36, 472. [Google Scholar] [CrossRef]
- Singh, K.B.; Weigand, S. Identification of resistant sources in Cicer species to Liriomyza cicerina. Genet. Resour. Crop Evol. 1994, 41, 75–79. [Google Scholar] [CrossRef]
- Singh, K.B.; Robertson, L.D.; Ocampo, B. Diversity for abiotic and biotic stress resistance in the wild annual Cicer species. Genet. Resour. Crop Evol. 2004, 45, 9–17. [Google Scholar] [CrossRef]
- Ceylan, F.O.; Sarı, H.; Sarı, D.; Adak, A.; Erler, F.; Toker, C. Revealing of resistant sources in Cicer species to chickpea leaf miner, Liriomyza cicerina (Rondani). Phytoparasitica 2018, 46, 635–643. [Google Scholar] [CrossRef]
- Ikten, C.; Ceylan, F.O.; Toker, C. Improvement of leaf miner [Liriomyza cicerina Rond.(Diptera: Agromyzidae)] resistance in Cicer species. Turk. Entomol. Derg. 2015, 39, 171–178. [Google Scholar] [CrossRef]
- Toker, C. Mutagenesis for resistance to abiotic stresses: Chickpea as model crop. In Mutagenesis: Exploring Novel Genes and Pathways; Tomlekova, N.B., Kozgar, M.I., Wani, M.R., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2014; pp. 215–238. [Google Scholar]
- Rembold, H. Malic acid in chickpea exudates. A marker for Heliothis resistance. Int Chickpea Newsl. 1981, 4, 18–19. [Google Scholar]
- Toker, C.; Karhan, M.; Ulger, S. Endogenous organic acid variations in different chickpea (Cicer arietinum L.) genotypes. Acta Agric. Scand B Soil Plant Sci. 2006, 1, 42–44. [Google Scholar] [CrossRef]
- Cagirgan, M.I.; Toker, C.; Karhan, M.; Aksu, M.; Ulger, S.; Canci, H. Assessment of endogenous organic acid levels in Ascochyta Blight [Ascochyta rabiei (Pass.) Labr.] Susceptible and resistant chickpeas (Cicer arietinum L.). Turk. J. Field Crops 2011, 16, 121–124. [Google Scholar]
- Yoshida, M.; Cowgill, S.E.; Wightman, J.A. Mechanism of resistance to Helicoverpa armigera (Lepidoptera: Noctuidae) in chickpea: Role of oxalic acid in leaf exudate as an antibiotic factor. J. Econ. Entomol. 1995, 88, 1783–1786. [Google Scholar] [CrossRef]
- Yoshida, M.; Cowgill, S.E.; Wightman, J.A. Roles of oxalic acid and malic acids in chickpea trichome exudates in host-plant resistance to Helicoverpa armigera. J. Chem. Ecol. 1995, 23, 1195–1210. [Google Scholar] [CrossRef] [Green Version]
- Soltani, A.; Zouali, Y.; Haoual-Hamdi, S.; Saadouni, D.; Amri, M.; Carapelli, A.; Jemâa, J.M.B. Relationship between secondary metabolites and infestations caused by chickpea leafminer Liriomyza cicerina (Diptera: Agromyzidae). Int. J. Trop. Insect Sci. 2020, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Copeland, L. Role of malonate in chickpeas. Phytochemistry 2000, 54, 585–589. [Google Scholar] [CrossRef]
- Carapelli, A.; Soltani, A.; LeoVitale, M.; Amri, M.; Ben Jemâa, J. Cryptic diversity hidden within the leaf miner genus Liriomyza (Diptera: Agromyzidae). Genes 2018, 9, 554. [Google Scholar] [CrossRef] [Green Version]
- Ceylan, F.O.; Adak, A.; Sari, D.; Sari, H.; Toker, C. Unveiling of suppressed genes in interspecific and backcross populations derived from mutants of Cicer species. Crop Pasture Sci. 2019, 70, 254–262. [Google Scholar] [CrossRef]
- Kivrak, K.G.; Eker, T.; Sari, H.; Sari, D.; Akan, K.; Aydinoglu, B.; Catal, M.; Toker, C. Integration of Extra-Large-Seeded and Double-Podded Traits in Chickpea (Cicer arietinum L.). Agronomy 2020, 10, 901. [Google Scholar] [CrossRef]
- Sari, H.; Sari, D.; Eker, T.; Aydinoglu, B.; Canci, H.; Ikten, C.; Gokturk, R.S.; Zeybek, A.; Bakir, M.; Smykal, P.; et al. Inheritance and expressivity of neoplasm trait in crosses between domestic pea (Pisum sativum subsp. sativum) and the tall wild pea (Pisum sativum subsp. elatius). Agronomy 2020, 10, 1869. [Google Scholar] [CrossRef]
- Rasmusson, D.C.; Lambert, J.W. Variety × environment interactions in barley variety tests. Crop Sci. 1961, 1, 261–262. [Google Scholar] [CrossRef]
- Dewey, D.L.; Lu, K.H. A correlation and path-coefficient analysis of components of crested wheatgrass seed production. Agron. J. 1959, 51, 515–518. [Google Scholar] [CrossRef]
- Cattel, R.B. Factor analysis: An introduction to essentials. I. The purpose and underlying models. Biometrics 1965, 21, 190–215. [Google Scholar] [CrossRef]
- Cikman, E.; Civelek, H.S. Population densities of Liriomyza cicerina (Rondani, 1875) (Diptera: Agromyzidae) on Cicer arietinum L. (Leguminosae: Papilionoidea) in different irrigated conditions. Turk. J. Entomol. 2006, 30, 3–10. [Google Scholar]
- Mahesh, P.; Srikanth, J.; Chandran, K.; Nisha, M. Damage pattern and status of the leaf miner Aphanisticus aeneus Kerremans (Coleoptera: Buprestidae) in Saccharum spp. Int. J. Pest Manag. 2014, 61, 36–46. [Google Scholar] [CrossRef]
- Tran, D.H. Agromyzid leafminers and their parasitoids on vegetable in Vietnam. J. ISSAAS 2009, 15, 21–33. [Google Scholar]
- Koseoglu, K.; Adak, A.; Sari, D.; Sari, H.; Ceylan, F.O.; Toker, C. Transgressive segregations for yield criteria in reciprocal interspecific crosses between Cicer arietinum L. and C. reticulatum Ladiz. Euphytica 2017, 213, 1–16. [Google Scholar] [CrossRef]
- Adak, A.; Sari, D.; Sari, H.; Toker, C. Gene effects of Cicer reticulatum on qualitative and quantitative traits in the cultivated chickpea. Plant Breed. 2017, 136, 939–947. [Google Scholar] [CrossRef]
- Vega, U.; Frey, K.J. Transgressive segregation in inter and intraspecific crosses of barley. Euphytica 1980, 29, 585–594. [Google Scholar] [CrossRef]
- Singh, K.B.; Ocampo, B. Exploitation of wild Cicer species for yield improvement in chickpea. Theor. Appl. Genet. 1997, 95, 418–423. [Google Scholar] [CrossRef]
- Singh, S.; Gumber, R.K.; Joshi, N.; Singh, K. Introgression from wild Cicer reticulatum to cultivated chickpea for productivity and disease resistance. Plant Breed. 2005, 124, 477–480. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, K.; Bisht, I.S.; Dutta, M.; Rana, M.K.; Rana, J.C.; Bansal, C.K.; Sarker, A. Exploitation of wild annual Cicer species for widening the gene pool of chickpea cultivars. Plant Breed. 2015, 134, 186–192. [Google Scholar] [CrossRef]
- Sithanantham, S.; Reed, W. Preliminary observations on Heliothis and other insect pests on chickpea in Syria. Int. Chick News. 1980, 2, 15. [Google Scholar]
- Toker, C.; Adak, A.; Sari, D.; Sari, H.; Ceylan, F.O.; Canci, H.; Ikten, C.; Erler, F.; Upadhyaya, H.D. Registration of AWC 612M Chickpea Mutant Germplasm Line Resistant to Leaf Miner (Liriomyza cicerina). J. Plant Regist. 2019, 13, 87–90. [Google Scholar] [CrossRef]
- Malhotra, R.S.; El-Bouhssini, M.; Joubi, A. Registration of seven improved chickpea breeding lines resistant to leaf miner. J.Plant Regist. 2007, 1, 145–146. [Google Scholar] [CrossRef] [Green Version]
- Clement, S.L.; El-Din, S.; El-Dine, N.; Weigand, S.; Lateef, S.S. Research achievements in plant resistance to insect pests of cool season food legumes. Euphytica 1994, 73, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Edwards, O.; Singh, K.B. Resistance to insect pests: What do legumes have to offer? Euphytica 2006, 147, 273–285. [Google Scholar] [CrossRef]
- Kambrekar, D.N.; Balikai, R.A.; Giraddi, R.S.; Aruna, J.; Neelakanth, A. Host plant resistance in the management of chickpea pod borer, Helicoverpa armigera (Hubner). J. Agric. Sci. 2016, 29, 157–171. [Google Scholar]
- Ibanda, A.P.; Malinga, G.M.; Tanzito, G.A.; Ocan, D.; Badji, A.; Mwila, N.; Rubaihayo, P.R. Combining ability and heritability of soybean resistance to groundnut leaf miner. Euphytica 2018, 214, 192. [Google Scholar] [CrossRef]
- Weigand, S.; Tahhan, O. Chickpea insect pests in the Mediterranean zones and new approaches to their management. In Proceedings of the Second International Workshop on Chickpea Improvement, Patancheru, India, 4–8 December 1989; pp. 169–175. [Google Scholar]
- Rees, T.A.P. Carbon metabolism in mitochondria. In Plant Physiology; Dennis, D.T., Turpin, D.H., Eds.; Biochemistry and Molecular Biology, Longman: London, UK; New York, NY, USA, 1990; pp. 106–143. [Google Scholar]
- Lopez-Bucio, J.; Nieto-Jacobo, M.F.; Ramirez-Rodrıguez, V.; Herrera-Estrella, L. Organic acid metabolism in plants: From adaptive physiology to transgenic varieties for cultivation in extreme soils. Plant Sci. 2000, 160, 1–13. [Google Scholar] [CrossRef]
- Michaeli, S.; Fromm, H. Closing the loop on the GABA shunt in plants: Are GABA metabolism and signaling entwined? Plant Sci. 2015, 6, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouche, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Ramputh, A.I.; Bown, A.W. Rapid g-aminobutyric acid synthesis and the inhibition of the growth and development of oblique-banded leaf-roller larvae. Plant Physiol. 1996, 111, 1349–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bown, A.W.; Hall, D.E.; Mac Gregor, K.B. Insect footsteps on leaves stimulate the accumulation of 4-aminobutyrate and can be visualized through increased chlorophyll fluorescence and superoxide production. Plant Physiol. 2002, 129, 1430–1434. [Google Scholar] [CrossRef] [Green Version]
- GRDC. GRDC GrowNotes Chickpea Southern Region. (Australia: Grains Research and Development Corporation); GRDC: Canberra, Australia, 2018. [Google Scholar]
- Bhagwat, V.R.; Aherkar, S.K.; Satpute, U.S.; Thakare, H.S. Screening of chickpea (Cicer arietinum L.) genotypes for resistance to gram pod borer, Heliothis armigera (Hubner) and its relationship with malic acid in leaf exudates. J. Entomol. Res. 1995, 19, 249–253. [Google Scholar]
- Rembold, H.; Schroth, A.; Lateef, S.S.; Weigner, C.H. Semiochemicals and host-plant selection by Helicoverpa armigera: basic studies in the laboratory for the field. In Proceedings of the First Consultative Group Meeting on Host Selection Behavior of Helicoverpa armigera, Patancheru, India, 5–7 March 1990; ICRISAT: Patancheru, India, 1990; pp. 23–26. [Google Scholar]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Shahid, M.A.; Babar, M.A. Role of sugars, amino acids and organic acids in improving plant abiotic stress tolerance. Pak. J. Bot. 2020, 52, 1–9. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. ABA-based chemical signaling: The coordination of responses to stress in plants. Plant Cell Environ. 2002, 25, 195–210. [Google Scholar] [CrossRef]
- Stafford, H.A. Distribution of Tartaric Acid in the Leaves of Certain Angiosperms. Am. J. Bot. 1959, 46, 347–352. [Google Scholar] [CrossRef]
- Wang, X.; Kaçar, G.; Daane, K.M. Temporal Dynamics of Host Use by Drosophila suzukii in California’s San Joaquin Valley: Implications for Area-Wide Pest Management. Insects 2019, 10, 206. [Google Scholar] [CrossRef] [Green Version]
- Etienne, A.; Genard, M.; Lobit, P.; Mbeguie, A.; Mbeguie, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [Green Version]
- Darandeeh, N.; Hadavi, E. Effects of pre-harvest foliar application of citric acid and malic acid on chlorophyll II content and post-harvest vase life of lilium cv brunello. Front. Plant. Sci. 2012, 2, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timpa, I.D.; Burke, J.B.; Quisenberry, J.E.; Wendt, C.W. Effects of Water Stress on the Organic Acid and Carbohydrate Compositions of Cotton Plants. Plant Physiol. 1986, 82, 724–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Shi, L.X.; Jiao, Y.; Li, M.X.; Zhong, X.L.; Gu, F.X.; Liu, Q.; Xia, X.; Li, H.R. Metabolic responses to drought stress in the tissues of drought-tolerant and drought-sensitive wheat genotype seedlings. AoBP 2018, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Eaton, F.M.; Engle, D.R. Organic acids of the cotton plant. Plant Physiol. 1949, 24, 373–386. [Google Scholar]
- Barros, J.; Serk, H.; Granlundz, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, F.C.; Del Campo, M.L.; Grant, J.B.; Weibel, D.B.; Smedley, S.R.; Bolton, K.L.; Meinwald, J.; Eisner, T. Pinoresinol: A lignol of plant origin serving for defense in a caterpillar. Proc. Natl. Acad. Sci. USA 2006, 103, 15497–15501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miedes, E.; Vanholme, R.; Boerjan, W.; Molina, A. The role of the secondary cell wall in plant resistance to pathogens. Front. Plant Sci. 2014, 5, 1–13. [Google Scholar]
- Machado, A.R.T.; Campos, V.A.C.; Da Silva, W.J.R. Metabolic profiling in the roots of coffee plants exposed to the coffee root-knot nematode, Meloidogyne exigua. Eur. J. Plant Pathol. 2012, 134, 431–441. [Google Scholar] [CrossRef]
- Leuschner, C.; Herrmann, K.M.; Schultz, G. The metabolism of quinate in pea roots. Plant Physiol. 1995, 108, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, S.C.; Logan, B.A. Energy dissipation and radical scavenging by the plant phenylpropanoid pathway. Philos. Trans. R Soc. Lond. B Biol. Sci. 2000, 355, 1499–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sree Latha, E.; Sharma, H.C. Antixenosis Mechanism of Resistance to Helicoverpa armigera (Hub.) in Chickpea (Cicer arietinum Linn.). Int. J. Curr. Microbiol. App. Sci. 2018, 7, 1233–1238. [Google Scholar] [CrossRef] [Green Version]
- Nakata, P.A. An assessment of engineered calcium oxalate formation on plant growth and development as a step toward evaluating enhance plant defense. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Shivay, Y.S. Oxalic acid/oxalates in plants: From self-defence to phytoremediation. Curr. Sci. 2017, 112, 1665–1667. [Google Scholar] [CrossRef]
- ICARDA. ICARDA Annual Report. Mega-Project 2. Integrated Gene Management: Conservation, Enhancement and Utilization of Agro-Biodiversity in Dry Areas. Cereal Leafminer: An Emerging Threat; ICARDA: Aleppo, Syria, 2005; Volume 34. [Google Scholar]
- Golla, S.K.; Rajasekhar, P.; Sharma, S.P.; Prasad, K.H.; Sharma, H.C. Antixenosis and antibiosis mechanisms of resistance to pod borer, Helicoverpa armigera in wild relatives of chickpea, Cicer arietinum. Euphytica. 2018, 214, 1–16. [Google Scholar] [CrossRef]
- Narayanamma, V.L.; Sharma, H.C.; Vijay, P.M.; Gowda, C.L.L.; Sriramulu, M. Expression of resistance to the pod borer Helicoverpa armigera (Lepidoptera: Noctuidae), in relation to high performance liquid chromatography fingerprints of leaf exudates of chickpea. Int. J. Trop. 2013, 33, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Patnaik, H.P.; Senapati, B. Comparative tolerance of chickpea cultivars against Helicoverpa armigera. Ann. Plant Prot. Sci. 2001, 2, 324–325. [Google Scholar]
- Srivastava, C.P.; Srivastava, R.P. Screening for resistance to the gram pod borer H. armigera in chickpea genotypes and obviations on its mechanisms of resistance in India. Insect Sci. Appl. 1989, 10, 255–258. [Google Scholar]
- Selvanarayanan, V.; Narayanasamy, P. Assessment of tomato germplasm for resistance to fruit borer Helicoverpa armigera (Hubner). Int. J. Veg. Sci. 2006, 12, 71–79. [Google Scholar] [CrossRef]
- Toker, C.; Cagirgan, M.I. The use of phenotypic correlations and factor analysis in determining characters for grain yield selection in Chickpea (Cicer arietinum L.). Hereditas 2004, 140, 226–228. [Google Scholar] [CrossRef]
- Khaliq, I.; Parveen, N.; Chowdhry, M.A. Correlation and path coefficient analyses in bread wheat. Int. J. Agric. Biol. 2004, 4, 633–635. [Google Scholar]
- Dragon, K. Application of factor analysis to determine the anthropogenic changes in groundwater quality. Prz. Geol. 2002, 50, 127–131. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Score | Response to Leaf Miner | Appearance of Plants |
|---|---|---|
| 1 | Very Highly Resistant | Free from any damage |
| 2 | Highly Resistant | Mines in less than 10% of the leaflets after careful observation |
| 3 | Resistant | Mines in less than 11–20% of the leaflets, no defoliation |
| 4 | Moderately Resistant | Mines present in 21 to 30% of the leaflets, no defoliation |
| 5 | Less susceptible | Mines present in 31 to 40% of the leaflets, some defoliation |
| 6 | Moderately susceptible | Mines in 41 to 50% of the leaflets, defoliation of 10% |
| 7 | Susceptible | Mines in 51 to 70% of the leaflets, defoliation 11–20% |
| 8 | Highly susceptible | Mines in 71 to 90% of the leaflets, defoliation 21–30% |
| 9 | Very highly susceptible | Mines in more than 91% of the leaflets and defoliation greater than 31% |
| Traits | Mean ± SE | Minimum | Maximum | Correlations |
|---|---|---|---|---|
| Resistance to leaf miner (RL) | 6.15 ± 0.8 | 3.00 | 9.00 | |
| Leaves per plant (LP) | 89.15 ± 1.5 | 31.00 | 176.50 | −0.141 * |
| Leaflets per leaf (LL) | 12.25 ± 0.6 | 10.00 | 14.00 | 0.018 |
| Leaflet length (LH) | 0.86 ± 0.01 | 0.50 | 1.30 | −0.267 ** |
| Leaflet width (LW) | 0.49 ± 0.01 | 0.30 | 1.00 | −0.234 ** |
| Plant height (PH) | 31.82 ± 0.3 | 21.50 | 48.00 | −0.314 ** |
| Pods per plant (PP) | 23.37 ± 0.7 | 6.00 | 63.00 | −0.071 |
| Biological yield (BY) | 7.27 ± 0.3 | 1.40 | 25.68 | −0.076 |
| Seed yield (SY) | 3.46 ± 0.1 | 0.25 | 11.81 | −0.213 ** |
| 100-seed weight (SW) | 16.76 ± 0.3 | 4.83 | 29.24 | −0.092 |
| Traits | LP | LL | LH | LW | PH | PP | BY | SY | SW | GH | PA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaves per plant (LP) | 0.012 | 0.028 | 0.060 | −0.014 | 0.324 * | 0.004 | −0.025 | 0.014 | −0.034 | −0.062 | 0.063 |
| Leaflets per leaf (LL) | 0.022 | −0.004 | −0.006 | 0.029 | 0.050 | 0.023 | 0.071 | −0.156 | 0.044 | −0.044 | 0.032 |
| Leaflet length (LH) | −0.034 | 0.089 | −0.058 | 0.847 | 0.264 * | 0.075 | −0.199 | 0.020 | 0.363 | −0.154 | −0.620 |
| Leaflet width (LW) | 0.165 | −0.019 | 0.743 | −0.146 | 0.002 | −0.109 | 0.182 | −0.098 | −0.137 | 0.144 | 0.563 |
| Plant height (PH) | 0.422 | 0.082 | 0.126 | 0.001 | −0.254 * | 0.032 | 0.015 | 0.188 * | 0.013 | −0.282 * | −0.128 |
| Pods per plant (PP) | 0.006 | 0.047 | 0.044 | −0.072 | 0.039 | 0.032 | 0.480 | 0.425 * | −0.452 | −0.015 | −0.103 |
| Biological yield (BY) | −0.036 | 0.129 | −0.106 | 0.110 | 0.016 | 0.440 | 0.073 | 0.199 | 0.341 | −0.231 * | −0.189 |
| Seed yield (SY) | 0.021 | −0.286 | 0.011 | −0.060 | 0.212 | 0.392 | 0.200 | −0.223 * | 0.349 | 0.083 | 0.209 |
| 100-seed weight (SW) | −0.037 | 0.060 | 0.144 | −0.062 | 0.010 | −0.309 | 0.254 | 0.258* | 0.035 | 0.171 * | 0.164 |
| Growth habit (GH) | −0.056 | −0.051 | −0.051 | 0.054 | −0.196 | −0.009 | −0.144 | 0.052 | 0.143 | −0.172 * | −0.167 |
| Pods per axil (PA) | 0.060 | 0.038 | −0.217 | 0.224 | −0.094 | −0.062 | −0.124 | 0.137 | 0.145 | −0.176 * | 0.117 |
| Organic Acids | OA | QA | MA | CA | SA | TA | Correlations |
|---|---|---|---|---|---|---|---|
| Oxalic acid (OA) | −0.304 * | 0.629 * | 0.081 | 0.124 | −0.208 | −0.265 | −0.264 |
| Quinic acid (QA) | 0.302 | 0.034 | 0.283 | −0.174 | 0.353 * | 0.308 | −0.260 |
| Malic acid (MA) | 0.073 | 0.525 * | 0.253 | −0.104 | −0.069 | 0.043 | 0.047 |
| Citric acid (CA) | 0.135 | −0.393 | −0.127 | 0.293 | 0.429 * | 0.013 | 0.043 |
| Succinic acid (SA) | −0.179 | 0.633 * | −0.067 | 0.341 * | 0.760 ** | 0.030 | −0.587 ** |
| Tartaric acid (TA) | −0.243 | 0.587 * | 0.044 | 0.011 | 0.032 | 0.198 | 0.028 |
| Source of Variance | Factor 1 | Factor 2 | Factor 3 | Factor 4 | Communality |
|---|---|---|---|---|---|
| Quinic acid | 0.899 | 0.146 | 0.038 | −0.046 | 0.833 |
| Succinic acid | 0.649 | −0.624 | 0.260 | 0.093 | 0.887 |
| Malic acid | 0.646 | 0.482 | 0.002 | −0.211 | 0.694 |
| RL scores | −0.472 | 0.659 | 0.372 | −0.361 | 0.926 |
| Oxalic acid | 0.490 | 0.053 | −0.658 | −0.458 | 0.887 |
| Tartaric acid | 0.590 | 0.233 | 0.604 | 0.068 | 0.771 |
| Citric acid | −0.187 | −0.590 | 0.330 | −0.699 | 0.981 |
| Variance | 2.5 | 1.5 | 1.1 | 0.9 | 6.08 |
| Variance% | 35.6 | 21.2 | 15.9 | 12.7 | 85.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrigui, N.; Sari, D.; Sari, H.; Eker, T.; Cengiz, M.F.; Ikten, C.; Toker, C. Introgression of Resistance to Leafminer (Liriomyza cicerina Rondani) from Cicer reticulatum Ladiz. to C. arietinum L. and Relationships between Potential Biochemical Selection Criteria. Agronomy 2021, 11, 57. https://doi.org/10.3390/agronomy11010057
Chrigui N, Sari D, Sari H, Eker T, Cengiz MF, Ikten C, Toker C. Introgression of Resistance to Leafminer (Liriomyza cicerina Rondani) from Cicer reticulatum Ladiz. to C. arietinum L. and Relationships between Potential Biochemical Selection Criteria. Agronomy. 2021; 11(1):57. https://doi.org/10.3390/agronomy11010057
Chicago/Turabian StyleChrigui, Nesrine, Duygu Sari, Hatice Sari, Tuba Eker, Mehmet Fatih Cengiz, Cengiz Ikten, and Cengiz Toker. 2021. "Introgression of Resistance to Leafminer (Liriomyza cicerina Rondani) from Cicer reticulatum Ladiz. to C. arietinum L. and Relationships between Potential Biochemical Selection Criteria" Agronomy 11, no. 1: 57. https://doi.org/10.3390/agronomy11010057
APA StyleChrigui, N., Sari, D., Sari, H., Eker, T., Cengiz, M. F., Ikten, C., & Toker, C. (2021). Introgression of Resistance to Leafminer (Liriomyza cicerina Rondani) from Cicer reticulatum Ladiz. to C. arietinum L. and Relationships between Potential Biochemical Selection Criteria. Agronomy, 11(1), 57. https://doi.org/10.3390/agronomy11010057