


The Effect of Neutral Salt and Alkaline Stress with the Same Na+ Concentration on Root Growth of Soybean (Glycine max (L.) Merr.) Seedlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials, Growth, and Treatment Conditions
2.2. Biomass and Morphological Measurement
2.3. Measurement of Root Activity and Relative Electrical Conductivity
2.4. Measurement of Antioxidant Enzyme Activities and ROS Content
2.5. Measurement of Osmotic Regulator Content
2.6. DNA Extraction and RAPD Analysis
2.7. RNA Extraction, First-Strand cDNA Synthesis, and qRT-PCR Analysis
2.8. Statistical Analysis
3. Results
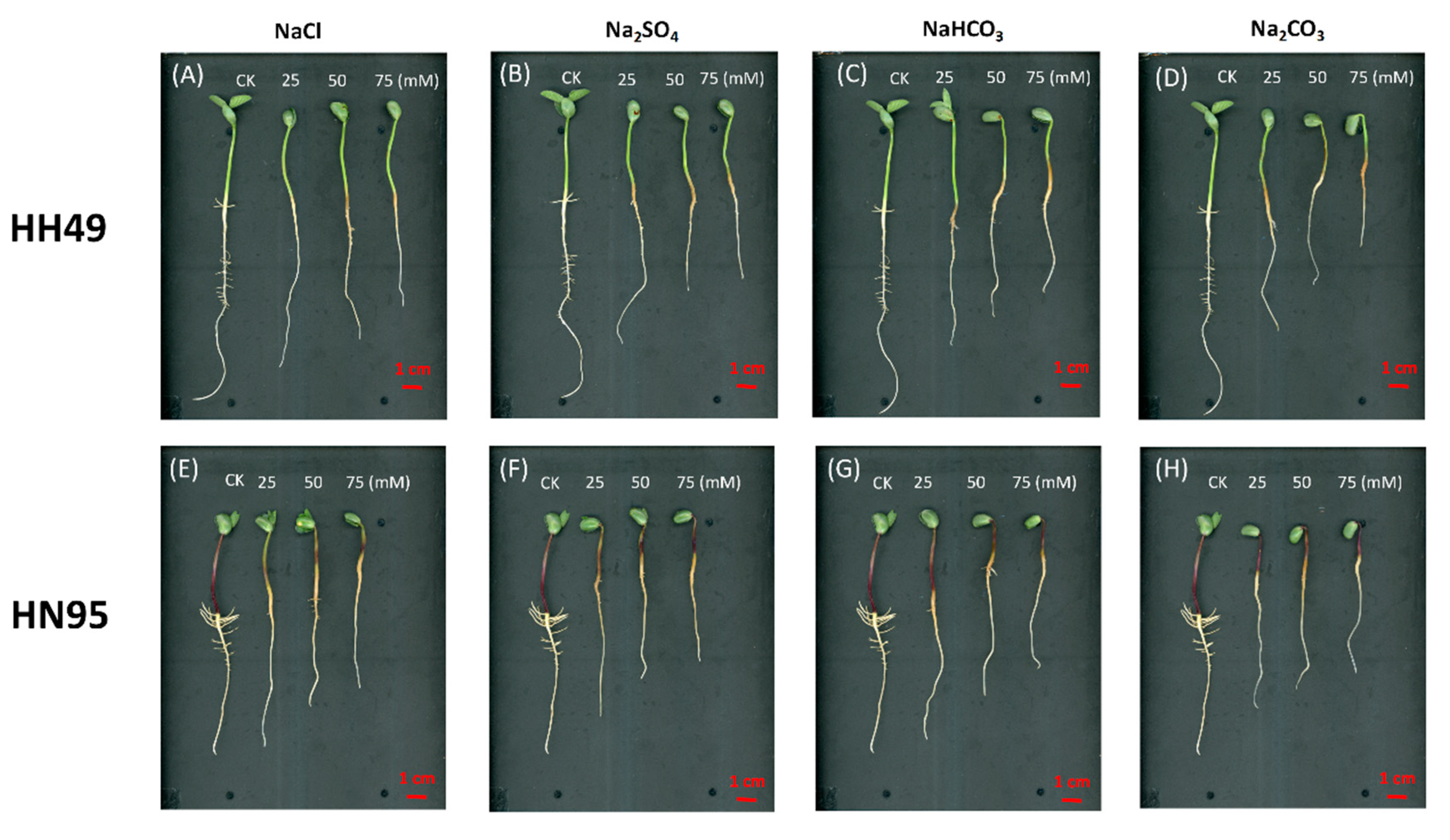
3.1. Effects of Different Salt Stress Treatments on the Root Growth of Soybean Seedlings
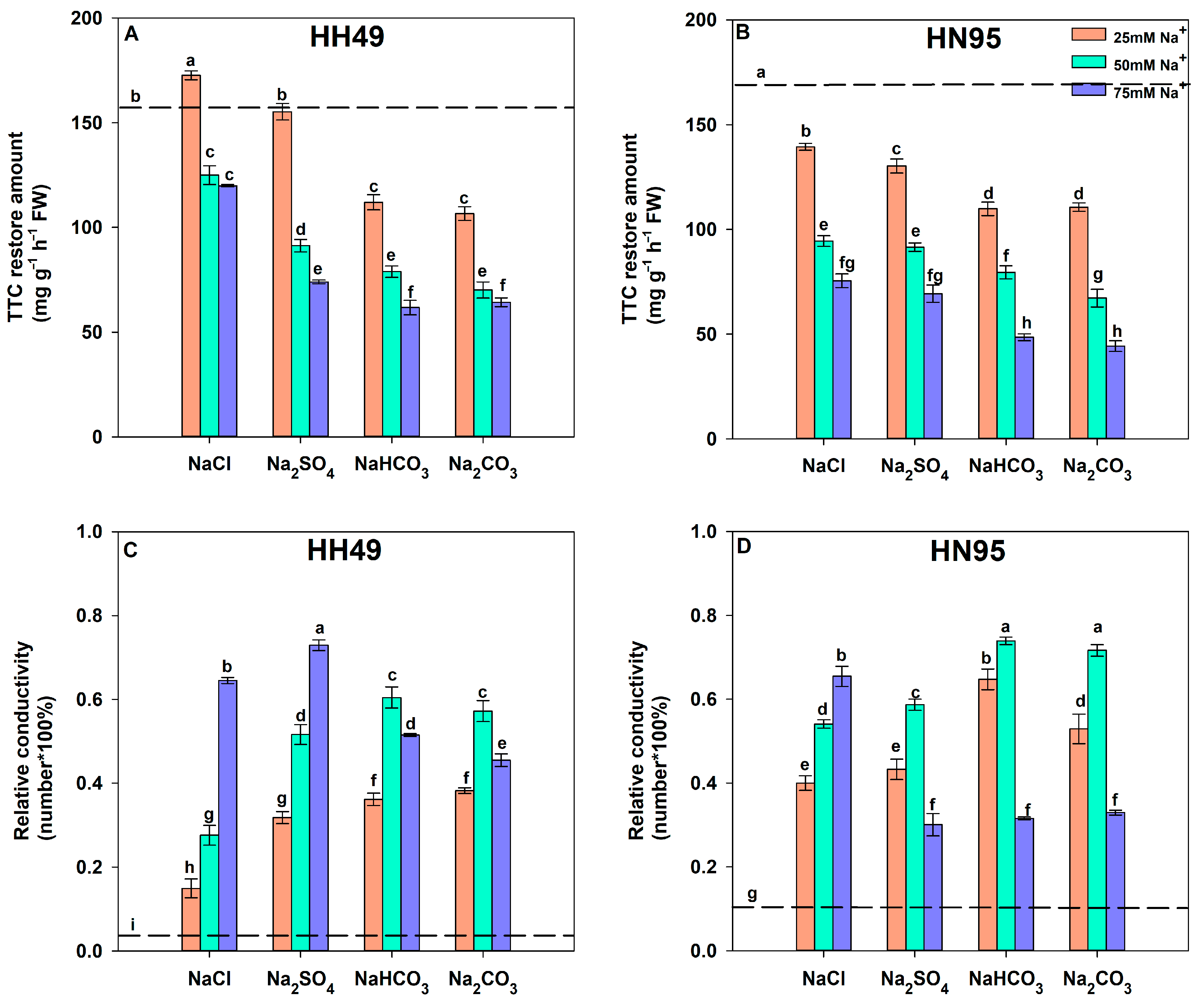
3.2. Effects of Different Salt Stress Treatments on the Root Activity and Relative Conductivity of Soybean Seedling Roots
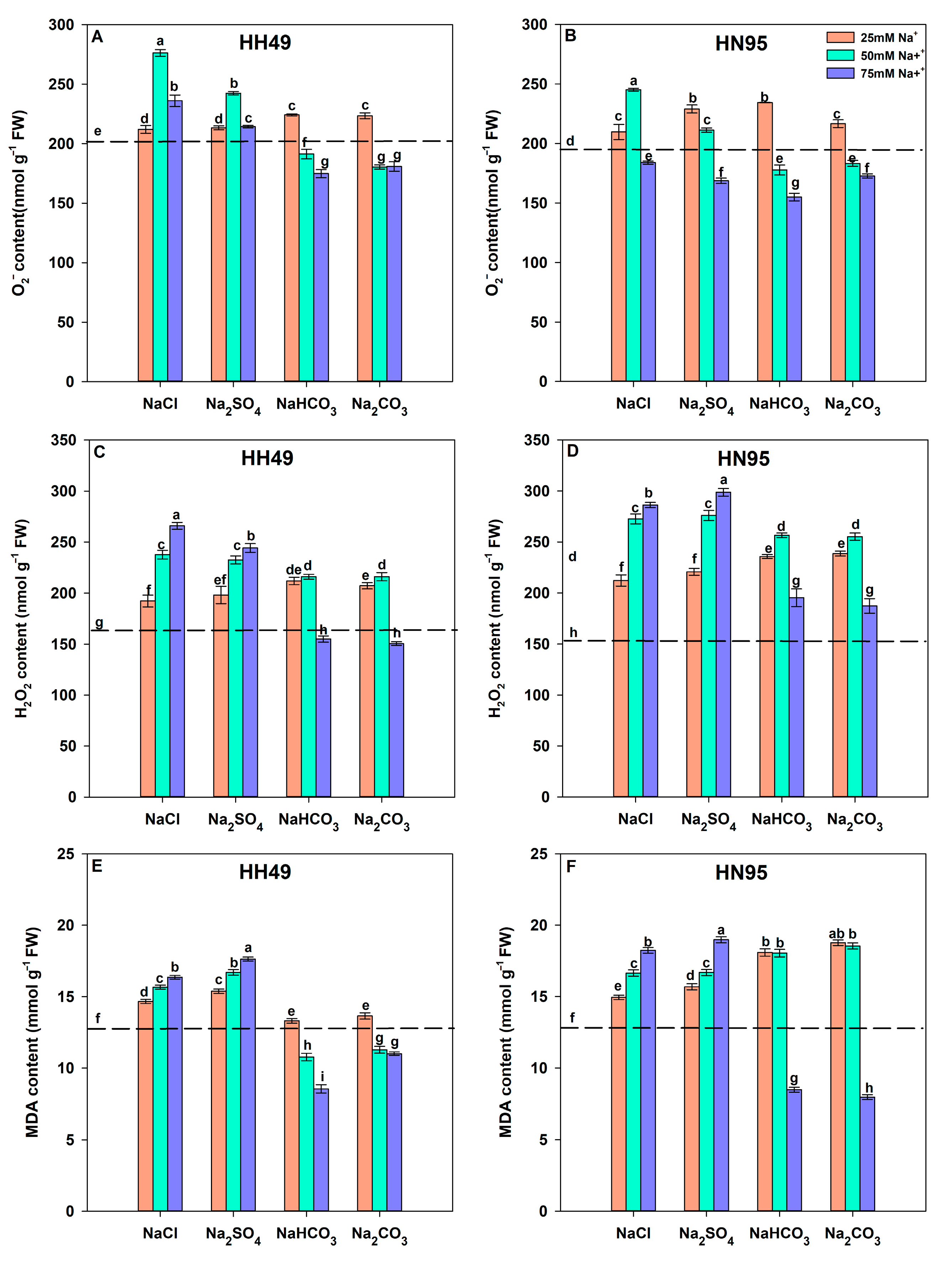
3.3. Effects of Different Salt Stress Treatments on ROS Accumulation in Soybean Seedling Roots
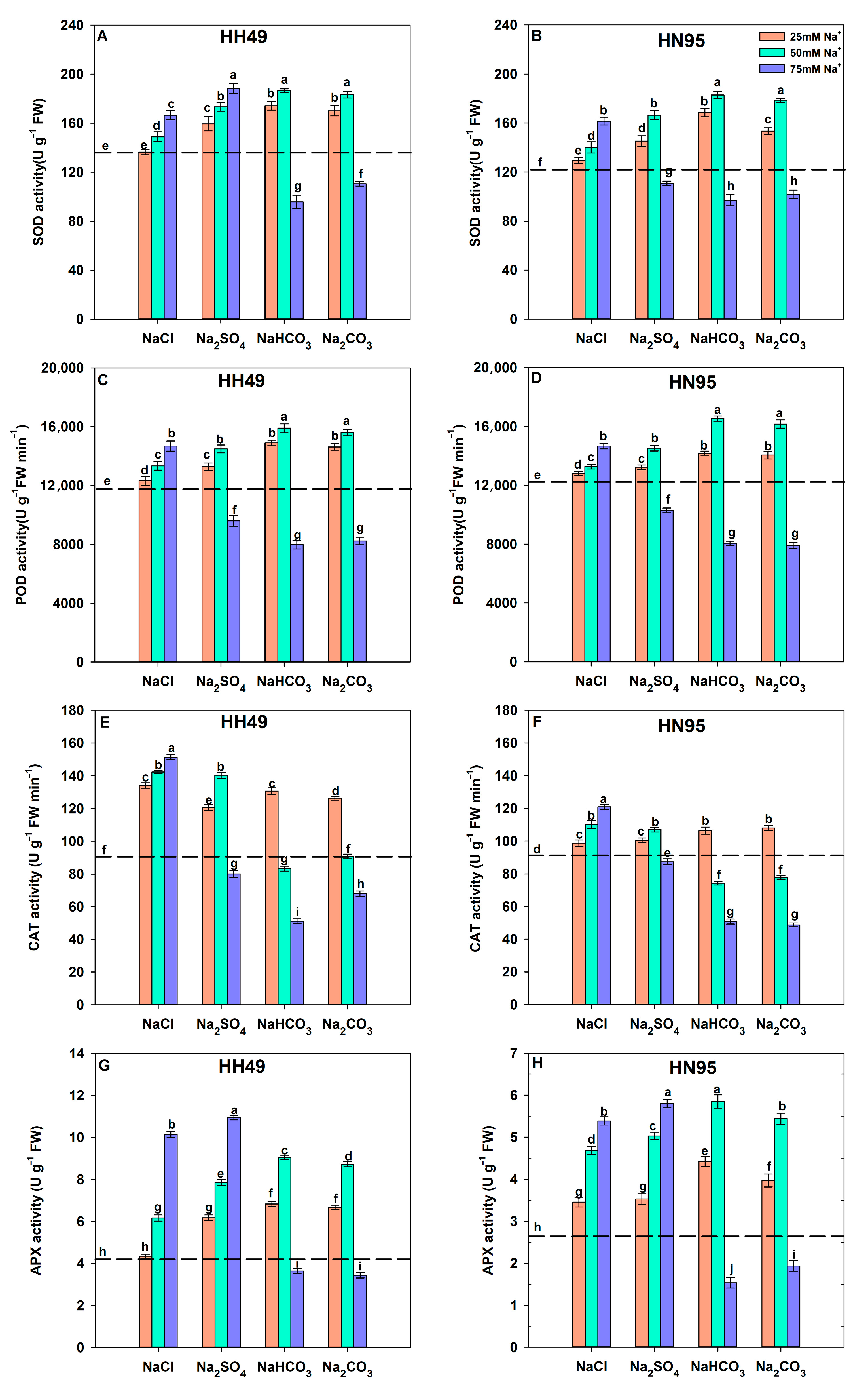
3.4. Effects of Different Salt Stress Treatments on Antioxidant Enzyme Activity in Soybean Seedling Roots
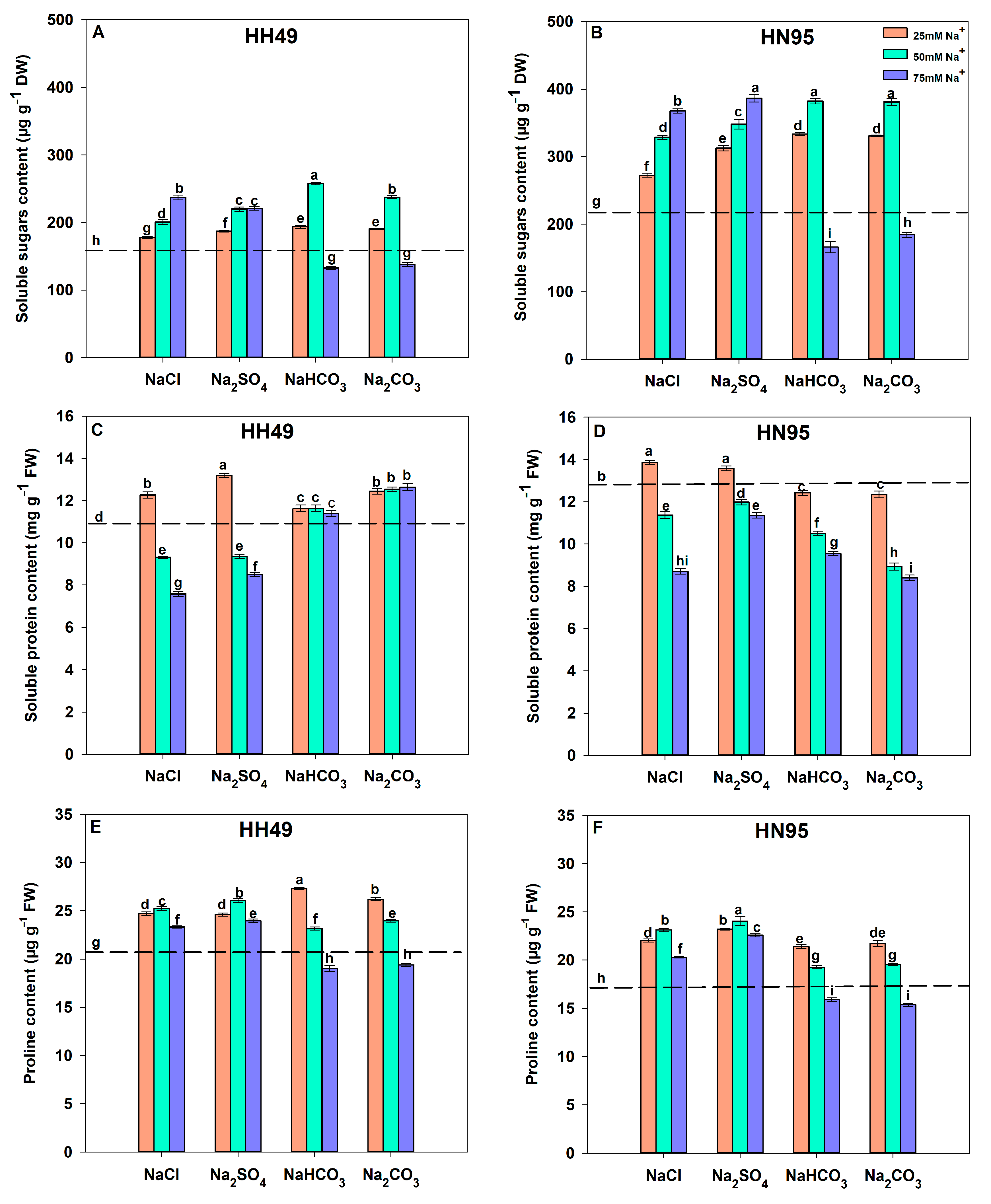
3.5. Effects of Different Salt Stress Treatments on Osmotic Regulator Content of Soybean Seedling Roots
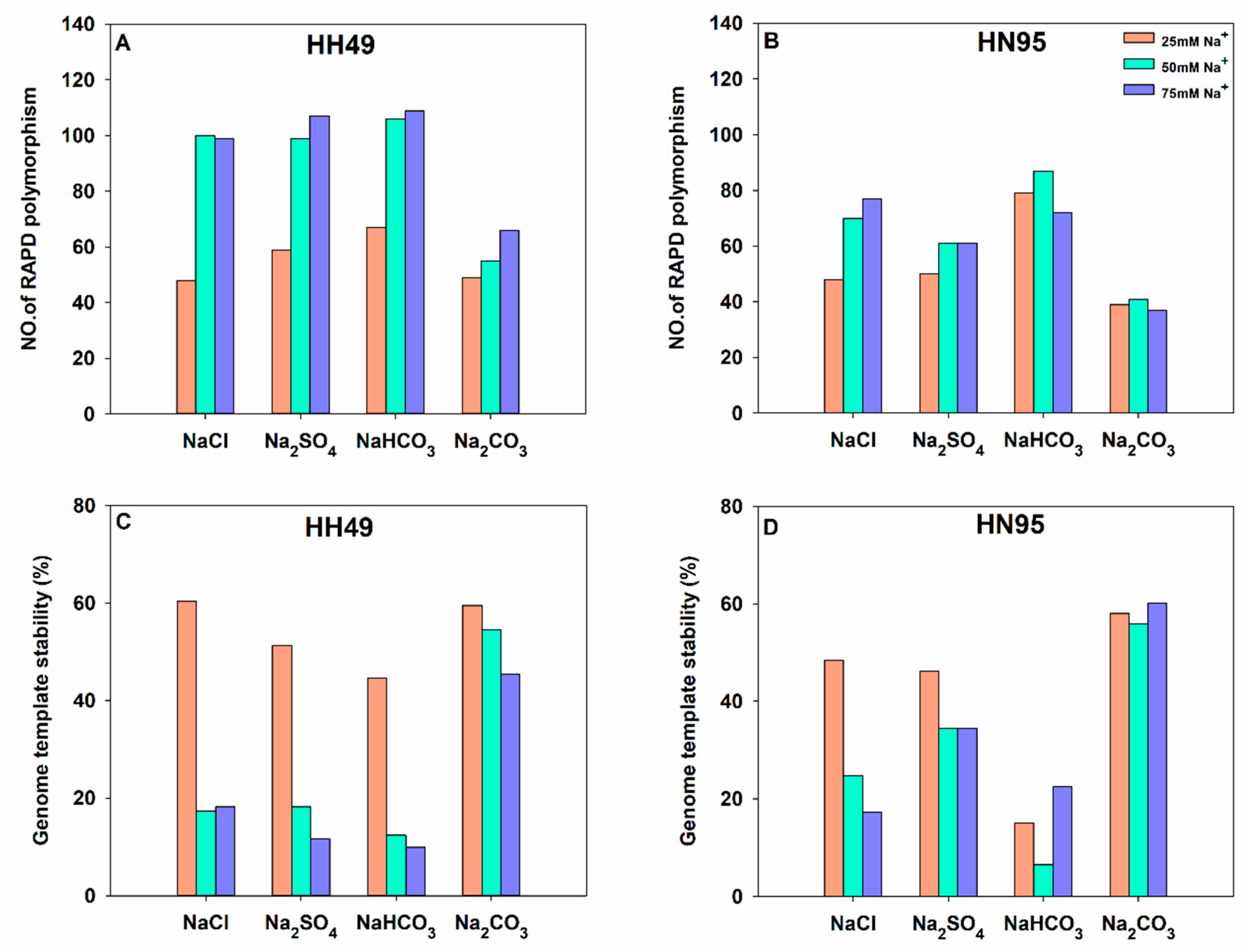
3.6. Effects of Different Salt Stress Treatments on DNA Oxidative Damage of Soybean Seedling Roots
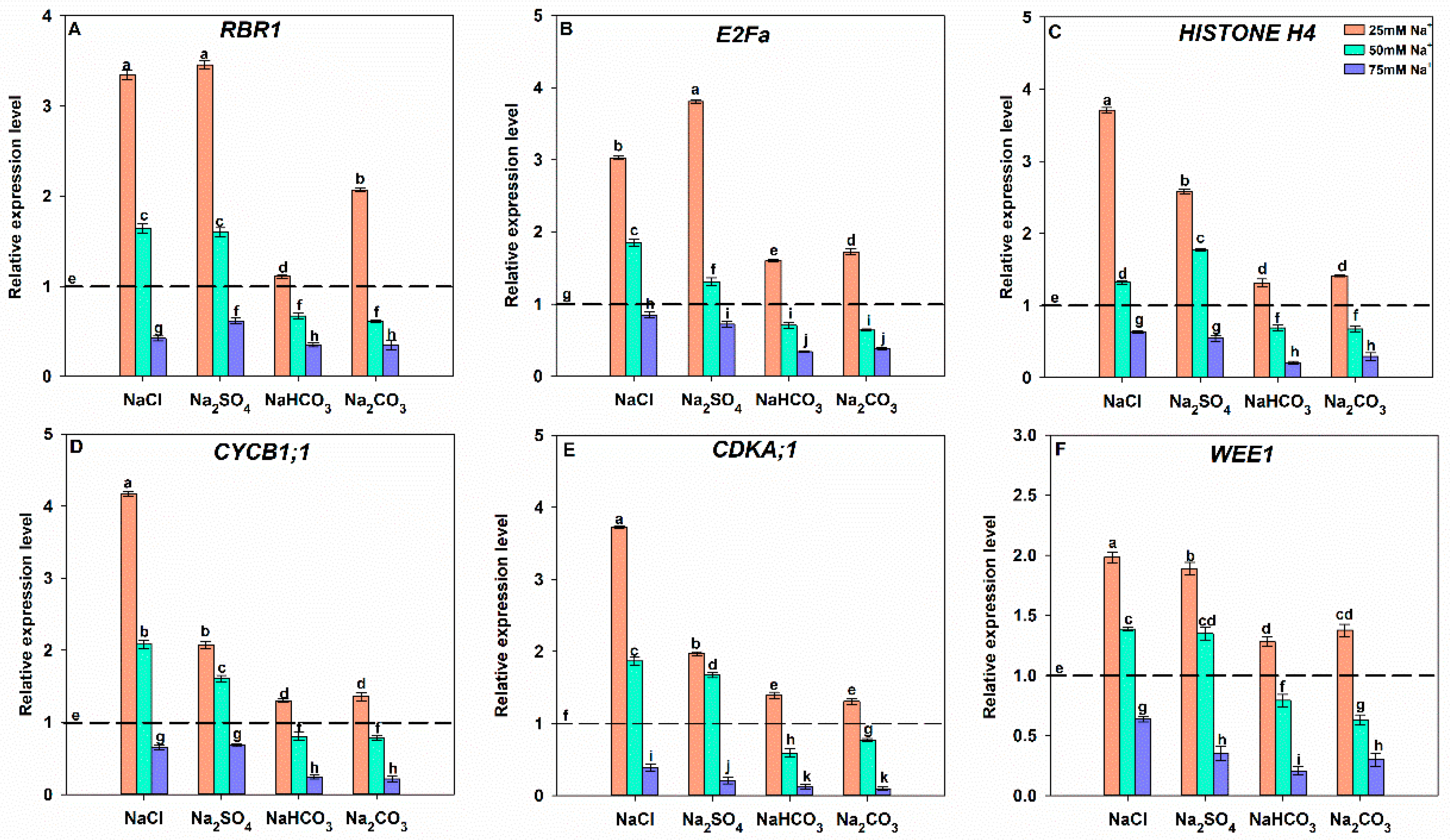
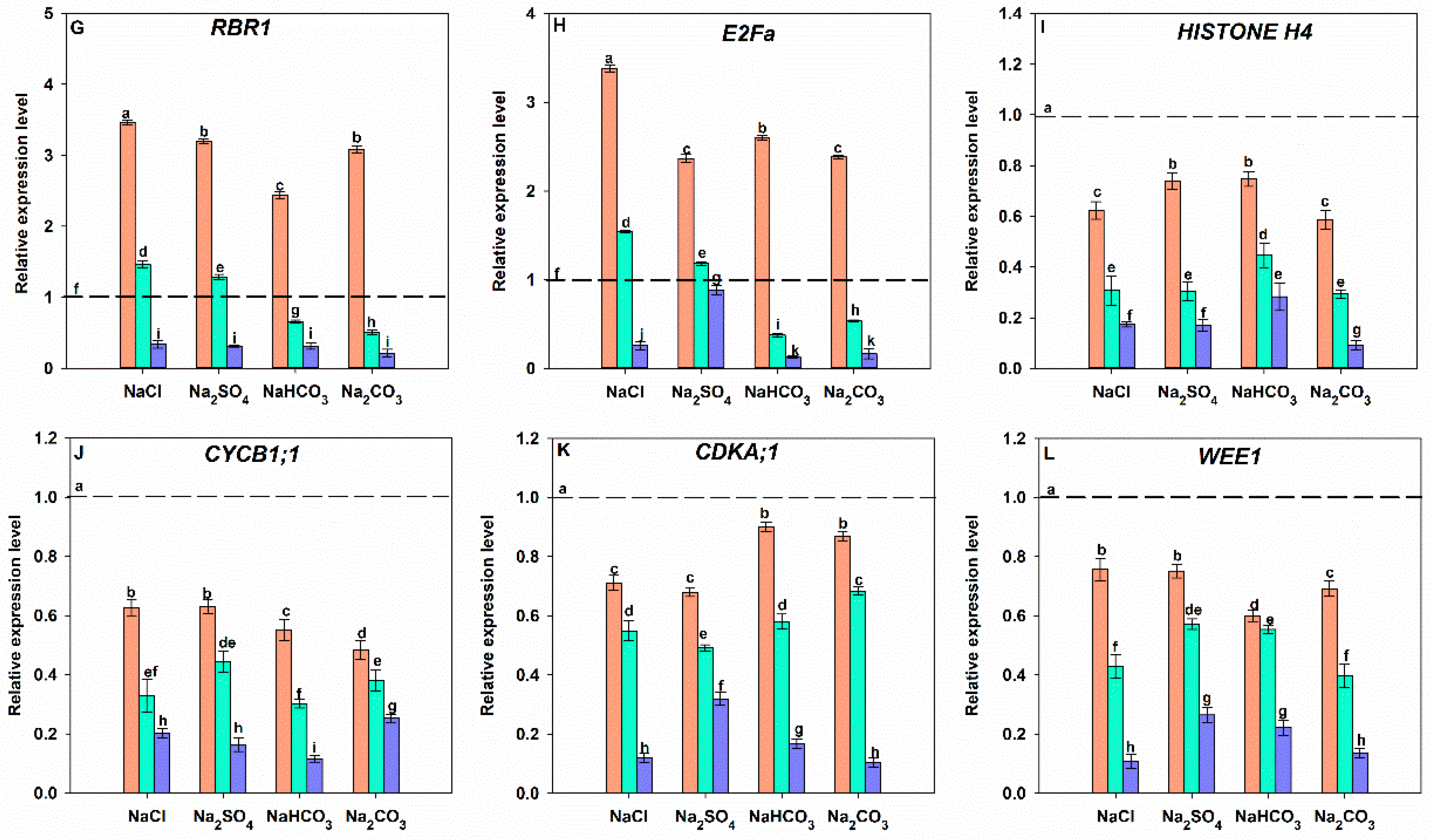
3.7. Effects of Different Salt Stress Treatments on Cell Cycle Progression of Soybean Seedling Roots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Jiang, X.; Zhao, C.; Fang, Z.; Jiao, P. Transcriptomic and metabolomic analysis reveals the role of CoA in the salt tolerance of Zygophyllum spp. BMC Plant Biol. 2020, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Leng, B.; Wang, B. Progress in Studying Salt Secretion from the Salt Glands in Recretohalophytes: How Do Plants Secrete Salt? Front. Plant Sci. 2016, 7, 977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robles, P.; Quesada, V. Transcriptional and Post-transcriptional Regulation of Organellar Gene Expression (OGE) and Its Roles in Plant Salt Tolerance. Int. J. Mol. Sci. 2019, 20, 1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Shen, F.; Xiao, Y.; Fang, H.; Qiu, C.; Li, W.; Wu, T.; Xu, X.; Wang, Y.; Zhang, X.; et al. Genomics-assisted prediction of salt and alkali tolerances and functional marker development in apple rootstocks. BMC Genom. 2020, 21, 550. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, X.; Lu, X.; Shu, N.; Wang, X.; Yang, X.; Wang, S.; Wang, J.; Guo, L.; Wang, D.; et al. Transcriptome Analysis of Gossypium hirsutum L. Reveals Different Mechanisms among NaCl, NaOH and Na2CO3 Stress Tolerance. Sci. Rep. 2018, 8, 13527. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Cui, J.; Cheng, D.; Dai, C.; Liu, T.; Wang, C.; Luo, C. iTRAQ protein profile analysis of sugar beet under salt stress: Different coping mechanisms in leaves and roots. BMC Plant Biol. 2020, 20, 347. [Google Scholar] [CrossRef]
- Gul Jan, F.; Hamayun, M.; Hussain, A.; Jan, G.; Iqbal, A.; Khan, A.; Lee, I.J. An endophytic isolate of the fungus Yarrowia lipolytica produces metabolites that ameliorate the negative impact of salt stress on the physiology of maize. BMC Microbiol. 2019, 19, 3. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Tang, X.; Yang, X.; Zhang, H. Comprehensive transcriptome and metabolome profiling reveal metabolic mechanisms of Nitraria sibirica Pall. to salt stress. Sci. Rep. 2021, 11, 12878. [Google Scholar] [CrossRef]
- Darko, E.; Gierczik, K.; Hudak, O.; Forgo, P.; Pal, M.; Turkosi, E.; Kovacs, V.; Dulai, S.; Majlath, I.; Molnar, I.; et al. Differing metabolic responses to salt stress in wheat-barley addition lines containing different 7H chromosomal fragments. PLoS ONE 2017, 12, e0174170. [Google Scholar] [CrossRef]
- Chauhan, A.; AbuAmarah, B.A.; Kumar, A.; Verma, J.S.; Ghramh, H.A.; Khan, K.A.; Ansari, M.J. Influence of gibberellic acid and different salt concentrations on germination percentage and physiological parameters of oat cultivars. Saudi J. Biol. Sci. 2019, 26, 1298–1304. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Li, J.; Yong, L.; Ai, F. Research progress of salt stress physiology and its adaptive regulation mechanism in plants. Ningxia Agric. For. Sci. Technol. 2007, 24, 34–36. [Google Scholar]
- Liu, C.; Liu, Y.; Guo, K.; Wang, S.; Yang, Y. Concentrations and resorption patterns of 13 nutrients in different plant functional types in the karst region of south-western China. Ann. Bot. 2014, 113, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Liu, L.; Du, M.; Tian, H.; Wang, B. Cation and Zn Accumulation in Brown Seeds of the Euhalophyte Suaeda salsa Improves Germination Under Saline Conditions. Front. Plant Sci. 2020, 11, 602427. [Google Scholar] [CrossRef]
- Li, X.; Song, Z.; Dong, Z. Salt stress physiology of plants. J. Northwest. Norm. Univ. (Nat. Sci. Ed.) 2004, 3, 106–111. [Google Scholar] [CrossRef]
- Lin, J.; Hua, X.; Peng, X.; Dong, B.; Yan, X. Germination Responses of Ryegrass (Annual vs. Perennial) Seed to the Interactive Effects of Temperature and Salt-Alkali Stress. Front. Plant Sci. 2018, 9, 1458. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Q.; Cai, H.; Xu, F. Ethylene Mediates Alkaline-Induced Rice Growth Inhibition by Negatively Regulating Plasma Membrane H(+)-ATPase Activity in Roots. Front. Plant Sci. 2017, 8, 1839. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Du, M.; Lu, C.; Wang, B. NaCl improves reproduction by enhancing starch accumulation in the ovules of the euhalophyte Suaeda salsa. BMC Plant Biol. 2020, 20, 262. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, X.; Hu, Z.; Leng, P. Nitric oxide modulating ion balance in Hylotelephium erythrostictum roots subjected to NaCl stress based on the analysis of transcriptome, fluorescence, and ion fluxes. Sci. Rep. 2019, 9, 18317. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.J.; Yang, H.Y.; Bai, J.P.; Liang, X.Y.; Lou, Y.; Zhang, J.L.; Wang, D.; Zhang, J.L.; Niu, S.Q.; Chen, Y.L. Ultrastructural and physiological responses of potato (Solanum tuberosum L.) plantlets to gradient saline stress. Front. Plant Sci. 2014, 5, 787. [Google Scholar] [CrossRef]
- Hashem, A.; Abd Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Shah, M.A. Induction of Osmoregulation and Modulation of Salt Stress in Acacia gerrardii Benth. by Arbuscular Mycorrhizal Fungi and Bacillus subtilis (BERA 71). Biomed. Res. Int. 2016, 2016, 6294098. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, L.; Sheng, X.; Yan, C.; Zhou, R.; Hang, J.; Yin, P.; Yan, N. Molecular basis for the selective and ABA-independent inhibition of PP2CA by PYL13. Cell Res. 2013, 23, 1369–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Zhang, T.; Zheng, A.; He, L.; Lai, R.; Liu, J.; Xing, P.; Tang, X. Exogenous proline induces regulation in 2-acetyl-1-pyrroline (2-AP) biosynthesis and quality characters in fragrant rice (Oryza sativa L.). Sci. Rep. 2020, 10, 13971. [Google Scholar] [CrossRef]
- Butt, M.; Sattar, A.; Abbas, T.; Hussain, R.; Ijaz, M.; Sher, A.; Shahzad, U.; Ullah, S.; Brestic, M.; Zivcak, M.; et al. Morpho-physiological and biochemical attributes of Chili (Capsicum annum L.) genotypes grown under varying salinity levels. PLoS ONE 2021, 16, e0257893. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Siddique, K.H.M.; Colmer, T.D. Vegetative and reproductive growth of salt-stressed chickpea are carbon-limited: Sucrose infusion at the reproductive stage improves salt tolerance. J. Exp. Bot. 2017, 68, 2001–2011. [Google Scholar] [CrossRef]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Wang, P.; Li, D.; Yin, T.; Zhuge, Q. Characterization, Expression Profiling, and Functional Analysis of PtDef, a Defensin-Encoding Gene From Populus trichocarpa. Front. Microbiol. 2020, 11, 106. [Google Scholar] [CrossRef]
- Hu, J.; Ren, B.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Comparative proteomic analysis reveals that exogenous 6-benzyladenine (6-BA) improves the defense system activity of waterlogged summer maize. BMC Plant Biol. 2020, 20, 44. [Google Scholar] [CrossRef]
- Iwaniuk, P.; Borusiewicz, A.; Lozowicka, B. Fluazinam and its mixtures induce diversified changes of crucial biochemical and antioxidant profile in leafy vegetable. Sci. Hortic. 2022, 298, 110988. [Google Scholar] [CrossRef]
- Li, S.; Cong, Y.; Liu, Y.; Wang, T.; Shuai, Q.; Chen, N.; Gai, J.; Li, Y. Optimization of Agrobacterium-Mediated Transformation in Soybean. Front. Plant Sci. 2017, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, X.; Gao, Y.; Yang, S. Genome-Wide Identification and Characterization of TALE Superfamily Genes in Soybean (Glycine max L.). Int. J. Mol. Sci. 2021, 22, 4117. [Google Scholar] [CrossRef]
- Cao, D.; Li, Y.; Liu, B.; Kong, F.; Tran, L.-S.P. Adaptive Mechanisms of Soybean Grown on Salt-Affected Soils. Land Degrad. Dev. 2018, 29, 1054–1064. [Google Scholar] [CrossRef]
- Pan, W.J.; Tao, J.J.; Cheng, T.; Bian, X.H.; Wei, W.; Zhang, W.K.; Ma, B.; Chen, S.Y.; Zhang, J.S. Soybean miR172a Improves Salt Tolerance and Can Function as a Long-Distance Signal. Mol. Plant 2016, 9, 1337–1340. [Google Scholar] [CrossRef] [Green Version]
- Zeng, A.; Chen, P.; Korth, K.L.; Ping, J.; Thomas, J.; Wu, C.; Srivastava, S.; Pereira, A.; Hancock, F.; Brye, K.; et al. RNA sequencing analysis of salt tolerance in soybean (Glycine max). Genomics 2019, 111, 629–635. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, D.; Li, M.; Shi, L. Metabolic Profiles Reveal Changes in Wild and Cultivated Soybean Seedling Leaves under Salt Stress. PLoS ONE 2016, 11, e0159622. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Xie, F. Effect of drought stress at reproductive stages on growth and nitrogen metabolism in soybean. Agronomy 2020, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. Ascorbate peroxidase-a hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Heupel, E.; A. Formation of hydrogen peroxide by isolated cell walls from horseradish (armoracia lapathifolia gilib.). Planta 1976, 130, 175–180. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants—Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Ji, W.; Cong, R.; Li, S.; Li, R.; Qin, Z.; Li, Y.; Zhou, X.; Chen, S.; Li, J. Comparative Proteomic Analysis of Soybean Leaves and Roots by iTRAQ Provides Insights into Response Mechanisms to Short-Term Salt Stress. Front. Plant Sci. 2016, 7, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Jin, H.; Suo, J.; Yu, W.; Zhou, M.; Dai, W.; Song, L.; Hu, Y.; Wu, J. Effect of Temperature and Humidity on Oil Quality of Harvested Torreya grandis cv. Merrillii Nuts During the after-Ripening Stage. Front. Plant Sci. 2020, 11, 573681. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, H.; Cai, J.; Bi, Y.; Li, D.; Song, F. Rice NAC transcription factor ONAC066 functions as a positive regulator of drought and oxidative stress response. BMC Plant Biol. 2019, 19, 278. [Google Scholar] [CrossRef]
- Wang, H.; He, L.; Song, J.; Cui, W.; Zhang, Y.; Jia, C.; Francis, D.; Rogers, H.J.; Sun, L.; Tai, P.; et al. Cadmium-induced genomic instability in Arabidopsis: Molecular toxicological biomarkers for early diagnosis of cadmium stress. Chemosphere 2016, 150, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, H.; Du, Y.; Rogers, H.J.; Wu, Z.; Jia, S.; Yao, X.; Xie, F.; Liu, W. MSH2 and MSH6 in Mismatch Repair System Account for Soybean (Glycine max (L.) Merr.) Tolerance to Cadmium Toxicity by Determining DNA Damage Response. J. Agric. Food Chem. 2020, 68, 1974–1985. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Alavilli, H.; Awasthi, J.P.; Rout, G.R.; Sahoo, L.; Lee, B.H.; Panda, S.K. Overexpression of a Barley Aquaporin Gene, HvPIP2;5 Confers Salt and Osmotic Stress Tolerance in Yeast and Plants. Front. Plant Sci. 2016, 7, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Huang, Z.; Li, M.; Hou, Z. Growth, ionic homeostasis, and physiological responses of cotton under different salt and alkali stresses. Sci. Rep. 2020, 10, 21844. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Shumayla; Sharma, S.; Kumar, R.; Mendu, V.; Singh, K.; Upadhyay, S.K. Genomic Dissection and Expression Profiling Revealed Functional Divergence in Triticum aestivum Leucine Rich Repeat Receptor Like Kinases (TaLRRKs). Front. Plant Sci. 2016, 7, 1374. [Google Scholar] [CrossRef] [Green Version]
- Wawrzynska, A.; Sirko, A. The Role of Selective Protein Degradation in the Regulation of Iron and Sulfur Homeostasis in Plants. Int. J. Mol. Sci. 2020, 21, 2771. [Google Scholar] [CrossRef] [Green Version]
- Geng, G.; Li, R.; Stevanato, P.; Lv, C.; Lu, Z.; Yu, L.; Wang, Y. Physiological and Transcriptome Analysis of Sugar Beet Reveals Different Mechanisms of Response to Neutral Salt and Alkaline Salt Stresses. Front. Plant Sci. 2020, 11, 571864. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Uddin, M.I.; Tanaka, K.; Yin, L.; Shi, Z.; Qi, Y.; Mano, J.; Matsui, K.; Shimomura, N.; Sakaki, T.; et al. Maintenance of Chloroplast Structure and Function by Overexpression of the Rice Monogalactosyldiacylglycerol Synthase Gene Leads to Enhanced Salt Tolerance in Tobacco. Plant Physiol. 2014, 165, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, D.; Zhou, R.; Wang, X.; Dossa, K.; Wang, L.; Zhang, Y.; Yu, J.; Gong, H.; Zhang, X.; et al. Transcriptome and metabolome analyses of two contrasting sesame genotypes reveal the crucial biological pathways involved in rapid adaptive response to salt stress. BMC Plant Biol. 2019, 19, 66. [Google Scholar] [CrossRef] [Green Version]
- Domingo, C.; Lalanne, E.; Catala, M.M.; Pla, E.; Reig-Valiente, J.L.; Talon, M. Physiological Basis and Transcriptional Profiling of Three Salt-Tolerant Mutant Lines of Rice. Front. Plant Sci. 2016, 7, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, D.; Sanad, M.; Duscha, K.; Lyons, M.; Liu, F.; Bozhkov, P.; Kunz, H.H.; Hu, J.; Neuhaus, H.E.; Steel, P.G.; et al. Impact of salt stress, cell death, and autophagy on peroxisomes: Quantitative and morphological analyses using small fluorescent probe N-BODIPY. Sci. Rep. 2017, 7, 39069. [Google Scholar] [CrossRef] [Green Version]
- Soni, S.; Kumar, A.; Sehrawat, N.; Kumar, A.; Kumar, N.; Lata, C.; Mann, A. Effect of saline irrigation on plant water traits, photosynthesis and ionic balance in durum wheat genotypes. Saudi J. Biol. Sci. 2021, 28, 2510–2517. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Yan, C.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Jia, Q.; Ji, S.; Gong, B.; Li, J.; Lu, G.; Gao, H. Gamma-aminobutyric acid (GABA) alleviates salt damage in tomato by modulating Na(+) uptake, the GAD gene, amino acid synthesis and reactive oxygen species metabolism. BMC Plant Biol. 2020, 20, 465. [Google Scholar] [CrossRef]
- Zhao, X.; Fang, J.; Li, S.; Gaur, U.; Xing, X.; Wang, H.; Zheng, W. Artemisinin Attenuated Hydrogen Peroxide (H2O2)-Induced Oxidative Injury in SH-SY5Y and Hippocampal Neurons via the Activation of AMPK Pathway. Int. J. Mol. Sci. 2019, 20, 2680. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Kashyap, S.P.; Kumari, N.; Mishra, P.; Prasad Moharana, D.; Aamir, M.; Singh, B.; Prasanna, H.C. Transcriptional regulation-mediating ROS homeostasis and physio-biochemical changes in wild tomato (Solanum chilense) and cultivated tomato (Solanum lycopersicum) under high salinity. Saudi J. Biol. Sci. 2020, 27, 1999–2009. [Google Scholar] [CrossRef]
- Nan, X.; Huihui, Z.; Haixiu, Z.; Yining, W.; Jinbo, L.; Li, X.; Zepeng, Y.; Wenxu, Z.; Yi, Q.; Guangyu, S. The Response of Photosynthetic Functions of F1 Cutting Seedlings From Physocarpus amurensis Maxim (female symbol) × Physocarpus opulifolius “Diabolo” (male symbol) and the Parental Seedlings to Salt Stress. Front. Plant Sci. 2018, 9, 714. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, W.; Chan, Z.; Wu, Y. Endogenous Cytokinin Overproduction Modulates ROS Homeostasis and Decreases Salt Stress Resistance in Arabidopsis Thaliana. Front. Plant Sci. 2015, 6, 1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, J.; Jiang, W.; Liu, J.; Yang, S.; Gai, J.; Li, Y. Identification and Analysis of NaHCO3 Stress Responsive Genes in Wild Soybean (Glycine soja) Roots by RNA-seq. Front. Plant Sci. 2016, 7, 1842. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Serrano, L.; Calatayud, A.; Lopez-Galarza, S.; Serrano, R.; Bueso, E. Uncovering salt tolerance mechanisms in pepper plants: A physiological and transcriptomic approach. BMC Plant Biol. 2021, 21, 169. [Google Scholar] [CrossRef]
- Jiang, Z.; Song, G.; Shan, X.; Wei, Z.; Liu, Y.; Jiang, C.; Jiang, Y.; Jin, F.; Li, Y. Association Analysis and Identification of ZmHKT1;5 Variation With Salt-Stress Tolerance. Front. Plant Sci. 2018, 9, 1485. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, M.; Liu, L.; Meng, F. Physiological and proteomic responses of diploid and tetraploid black locust (Robinia pseudoacacia L.) subjected to salt stress. Int. J. Mol. Sci. 2013, 14, 20299–20325. [Google Scholar] [CrossRef] [Green Version]
- Gurunathan, S.; Jeyaraj, M.; Kang, M.H.; Kim, J.H. The Effects of Apigenin-Biosynthesized Ultra-Small Platinum Nanoparticles on the Human Monocytic THP-1 Cell Line. Cells 2019, 8, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagano, A.; Araujo, S.S.; Macovei, A.; Leonetti, P.; Balestrazzi, A. The Seed Repair Response during Germination: Disclosing Correlations between DNA Repair, Antioxidant Response, and Chromatin Remodeling in Medicago truncatula. Front. Plant Sci. 2017, 8, 1972. [Google Scholar] [CrossRef] [PubMed]
- Reszka, E.; Lesicka, M.; Wieczorek, E.; Jablonska, E.; Janasik, B.; Stepnik, M.; Konecki, T.; Jablonowski, Z. Dysregulation of Redox Status in Urinary Bladder Cancer Patients. Cancers 2020, 12, 1296. [Google Scholar] [CrossRef]
- Buisson, R.; Dion-Cote, A.M.; Coulombe, Y.; Launay, H.; Cai, H.; Stasiak, A.Z.; Stasiak, A.; Xia, B.; Masson, J.Y. Cooperation of breast cancer proteins PALB2 and piccolo BRCA2 in stimulating homologous recombination. Nat. Struct. Mol. Biol. 2010, 17, 1247–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, V.M.; Mukherjee, B.; Wang, F.; Huang, S.C.; Stoffel, E.M.; Kastrinos, F.; Syngal, S.; Cooney, K.A.; Gruber, S.B. Elevated risk of prostate cancer among men with Lynch syndrome. J. Clin. Oncol. 2013, 31, 1713–1718. [Google Scholar] [CrossRef] [PubMed]
- Renaud-Young, M.; Lloyd, D.C.; Chatfield-Reed, K.; George, I.; Chua, G.; Cobb, J. The NuA4 complex promotes translesion synthesis (TLS)-mediated DNA damage tolerance. Genetics 2015, 199, 1065–1076. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Treatments | Na+ Concentration (mM) | Fresh Weight (g) | Dry Weight (g) | Length (cm) | Superficial Area (cm2) | Volume (cm3) | Length Inhibition Rate (%) |
|---|---|---|---|---|---|---|---|---|
| HH49 | CK | 0 | 1.34 ± 0.01 a | 92.41 ± 0.42 a | 37.97 ± 1.97 a | 73.71 ± 8.66 a | 16.25 ± 0.52 a | 0 h |
| NaCl | 25 | 1.29 ± 0.01 ab | 89.2 ± 0.69 ab | 30.95 ± 1.61 b | 55.54 ± 6.21 ab | 14.04 ± 1.32 abcd | 18.48 ± 0.01 g | |
| 50 | 1.20 ± 0.01 cd | 88.25 ± 0.67 b | 24.17 ± 1.14 b | 59.11 ± 5.2 ab | 11.19 ± 1.3 de | 36.34 ± 0.17 de | ||
| 75 | 1.14 ± 0.01 de | 86.62 ± 0.43 b | 20.99 ± 1.06 b | 38.15 ± 7.51 b | 5.22 ± 0.34 f | 44.72 ± 0.04 ab | ||
| Na2SO4 | 25 | 1.24 ± 0.02 bc | 85.84 ± 0.64 b | 26.86 ± 0.72 b | 56.41 ± 5.31 ab | 13.18 ± 1.53 abcd | 29.26 ± 1.03 f | |
| 50 | 1.19 ± 0.02 cde | 85.84 ± 0.23 b | 24.29 ± 0.89 b | 39.52 ± 3.84 b | 5.25 ± 0.87 f | 36.03 ± 0.56 e | ||
| 75 | 1.13 ± 0.01 e | 79.66 ± 0.61 c | 22.87 ± 1.84 b | 34.61 ± 7.79 b | 3.62 ± 0.94 f | 39.77 ± 1.00 cd | ||
| NaHCO3 | 25 | 1.21 ± 0.01 c | 88.63 ± 0.23 b | 26.49 ± 0.56 b | 60.98 ± 3.17 ab | 10.55 ± 0.76 de | 30.23 ± 1.24 f | |
| 50 | 1.01 ± 0.02 f | 85.61 ± 0.67 b | 22.60 ± 0.77 b | 51.30 ± 3.47 ab | 7.75 ± 0.50 ef | 40.48 ± 0.61 c | ||
| 75 | 0.84 ± 0.01 g | 78.64 ± 1.21 c | 20.62 ± 0.71 b | 33.53 ± 1.56 b | 4.45 ± 0.48 f | 45.69 ± 0.55 a | ||
| Na2CO3 | 25 | 1.25 ± 0.01 bc | 79.43 ± 0.62 c | 26.20 ± 0.62 b | 54.46 ± 6.7 ab | 11.89 ± 0.71 bcde | 31.00 ± 1.13 f | |
| 50 | 0.97 ± 0.01 f | 74.84 ± 0.87 d | 22.19 ± 1.34 b | 48.06 ± 4.89 ab | 6.35 ± 0.56 f | 41.56 ± 0.29 bc | ||
| 75 | 0.88 ± 0.02 g | 66.85 ± 1.23 e | 20.55 ± 0.59 b | 37.66 ± 3.42 b | 3.19 ± 0.26 f | 45.88 ± 0.73 a | ||
| HN95 | CK | 0 | 1.45 ± 0.01 a | 89.21 ± 0.88 a | 38.38 ± 2.64 a | 87.95 ± 7.98 a | 13.35 ± 2.64 a | 0 i |
| NaCl | 25 | 1.18 ± 0.02 c | 77.20 ± 1.21 bc | 28.65 ± 1.29 b | 72.90 ± 8.01 a | 12.01 ± 3.07 abc | 25.35 ± 1.03 h | |
| 50 | 1.11 ± 0.02 cd | 75.40 ± 0.83 cd | 21.21 ± 0.63 bc | 35.55 ± 5.69 bc | 5.16 ± 1.59 abcd | 44.74 ± 1.25 fg | ||
| 75 | 0.97 ± 0.01 e | 63.43 ± 1.23 fg | 16.25 ± 1.50 cd | 27.08 ± 5.15 bc | 3.77 ± 1.18 bcd | 57.66 ± 0.58 bc | ||
| Na2SO4 | 25 | 1.07 ± 0.01 d | 75.20 ± 1.22 cd | 22.73 ± 1.07 bc | 46.53 ± 1.48 b | 5.46 ± 1.33 abcd | 40.78 ± 0.75 g | |
| 50 | 0.96 ± 0.02 e | 70.40 ± 1.26 de | 20.43 ± 1.22 bcd | 24.90 ± 2.00 bc | 3.49 ± 0.94 cd | 46.77 ± 0.28 f | ||
| 75 | 0.72 ± 0.02 g | 55.10 ± 1.01 hi | 18.53 ± 0.77 cd | 22.39 ± 2.31 bc | 3.21 ± 1.00 d | 51.72 ± 0.76 de | ||
| NaHCO3 | 25 | 1.27 ± 0.01 b | 82.86 ± 0.83 b | 19.73 ± 0.94 bcd | 37.40 ± 3.87 bc | 5.87 ± 1.18 abcd | 48.59 ± 0.63 ef | |
| 50 | 0.89 ± 0.01 f | 65.61 ± 1.81 ef | 16.85 ± 1.65 cd | 25.47 ± 2.45 bc | 3.93 ± 0.72 bcd | 56.10 ± 0.74 bc | ||
| 75 | 0.76 ± 0.01 g | 58.70 ± 1.66 gh | 12.54 ± 1.09 d | 17.86 ± 2.07 c | 2.18 ± 0.53 d | 67.33 ± 0.34 a | ||
| Na2CO3 | 25 | 0.86 ± 0.01 f | 65.46 ± 0.81 ef | 21.25 ± 1.04 bc | 38.23 ± 4.78 bc | 7.40 ± 1.87 abcd | 44.63 ± 0.64 fg | |
| 50 | 0.72 ± 0.01 g | 55.00 ± 1.41 hi | 17.76 ± 2.29 cd | 27.35 ± 3.23 bc | 3.56 ± 0.52 cd | 53.73 ± 1.62 cd | ||
| 75 | 0.6 ± 0.02 h | 50.4 ± 1.28 i | 15.89 ± 1.23 cd | 16.03 ± 4.4 c | 2.34 ± 0.66 d | 58.60 ± 0.21 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Shen, W.; Zhang, Z.; Guo, S.; Hu, J.; Feng, R.; Zhao, Q.; Du, J.; Du, Y. The Effect of Neutral Salt and Alkaline Stress with the Same Na+ Concentration on Root Growth of Soybean (Glycine max (L.) Merr.) Seedlings. Agronomy 2022, 12, 2708. https://doi.org/10.3390/agronomy12112708
Wang G, Shen W, Zhang Z, Guo S, Hu J, Feng R, Zhao Q, Du J, Du Y. The Effect of Neutral Salt and Alkaline Stress with the Same Na+ Concentration on Root Growth of Soybean (Glycine max (L.) Merr.) Seedlings. Agronomy. 2022; 12(11):2708. https://doi.org/10.3390/agronomy12112708
Chicago/Turabian StyleWang, Guangda, Wanzheng Shen, Zhaoning Zhang, Shuang Guo, Jiachen Hu, Ruiqi Feng, Qiang Zhao, Jidao Du, and Yanli Du. 2022. "The Effect of Neutral Salt and Alkaline Stress with the Same Na+ Concentration on Root Growth of Soybean (Glycine max (L.) Merr.) Seedlings" Agronomy 12, no. 11: 2708. https://doi.org/10.3390/agronomy12112708
APA StyleWang, G., Shen, W., Zhang, Z., Guo, S., Hu, J., Feng, R., Zhao, Q., Du, J., & Du, Y. (2022). The Effect of Neutral Salt and Alkaline Stress with the Same Na+ Concentration on Root Growth of Soybean (Glycine max (L.) Merr.) Seedlings. Agronomy, 12(11), 2708. https://doi.org/10.3390/agronomy12112708