
Comparative Transcriptome Analyses between Resistant and Susceptible Varieties in Response to Soybean Mosaic Virus Infection


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Virus Inoculation
2.2. RNA Extraction, Library Construction, and Sequencing
2.3. Read Alignment and Differential Expression Gene Screening
2.4. Functional Enrichment and Pathway Analysis of DEGs
2.5. RNA Extraction and Gene Expression Analysis by qRT-PCR
3. Results
3.1. Phenotypic Identification of NN1138-2 and Kefeng-1 after Inoculation with SC18
3.2. Evaluation of RNA-Seq Data
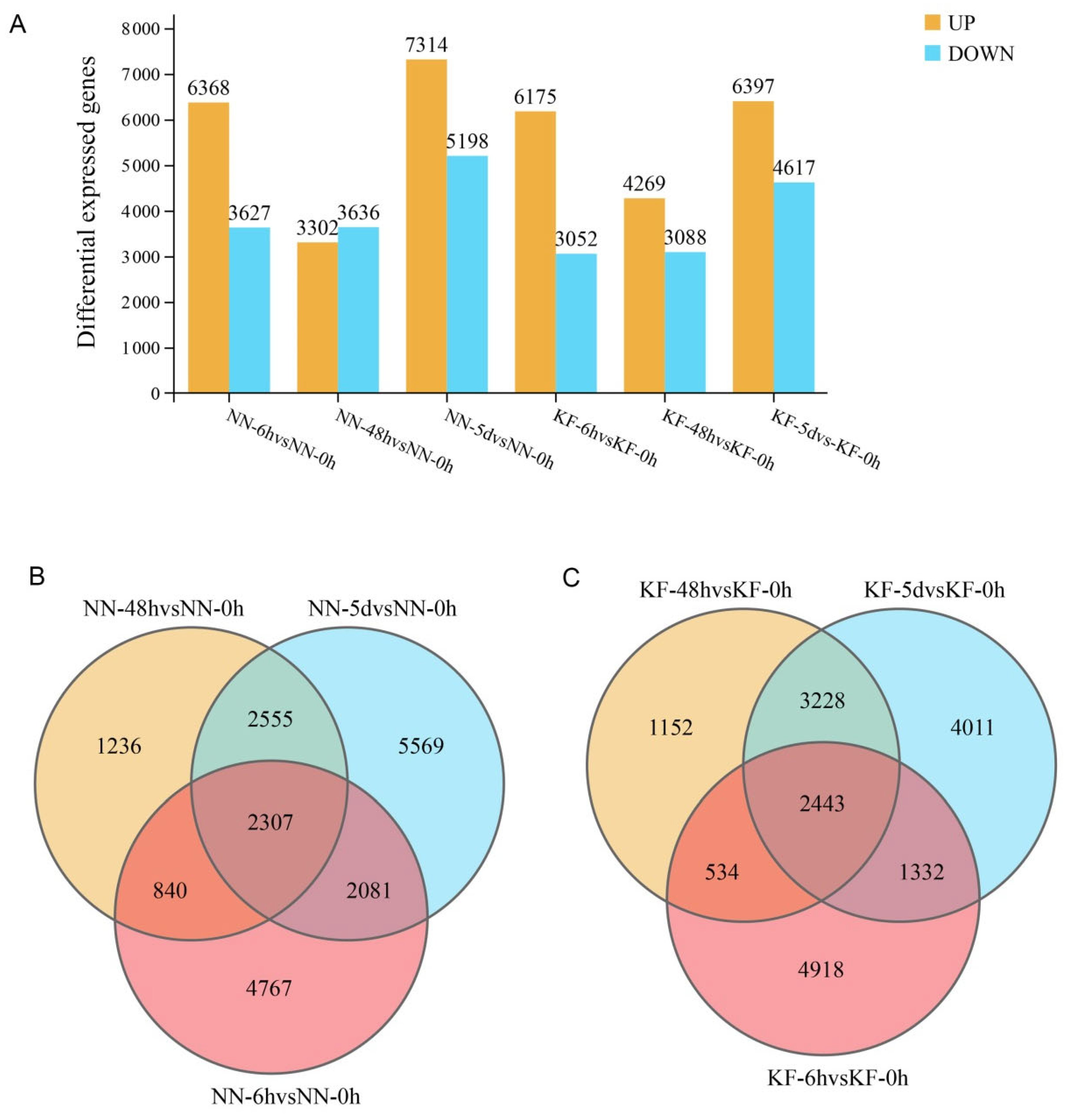
3.3. Statistics on the Number of DEGs
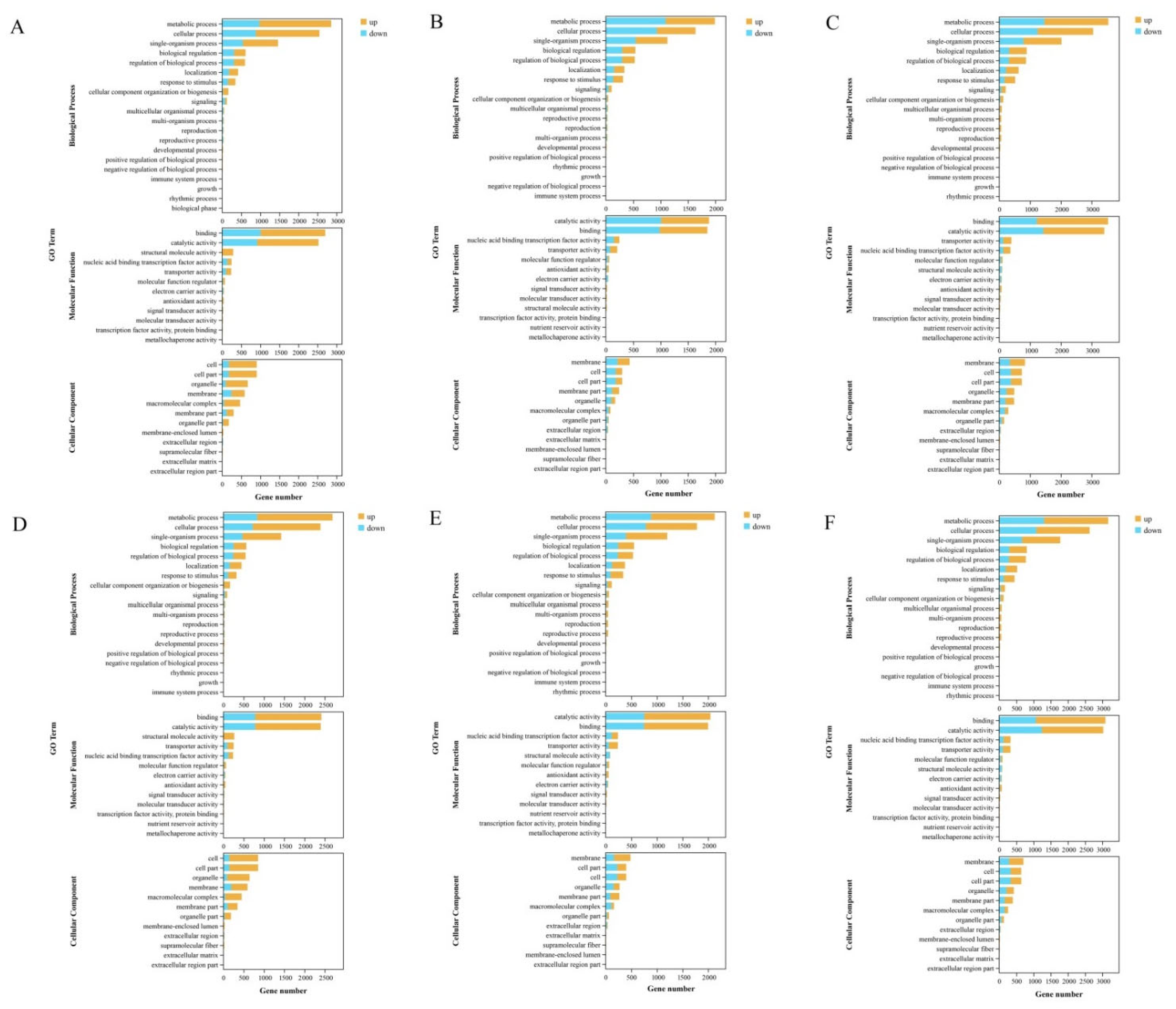
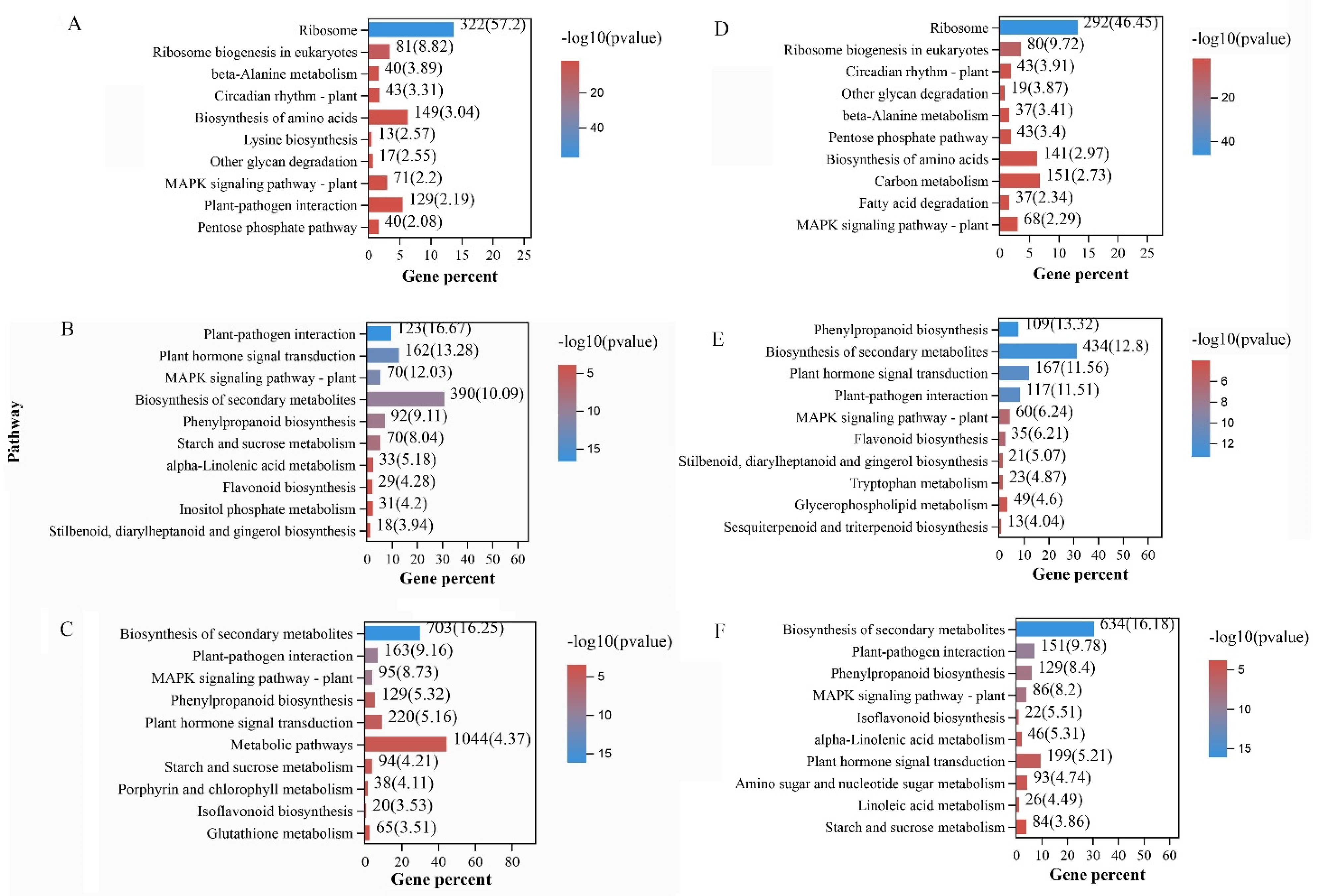
3.4. Functional Annotation and Pathway Enrichment Analysis of DEGs
3.5. Plant Hormone Effects on Soybean Defense against SMV
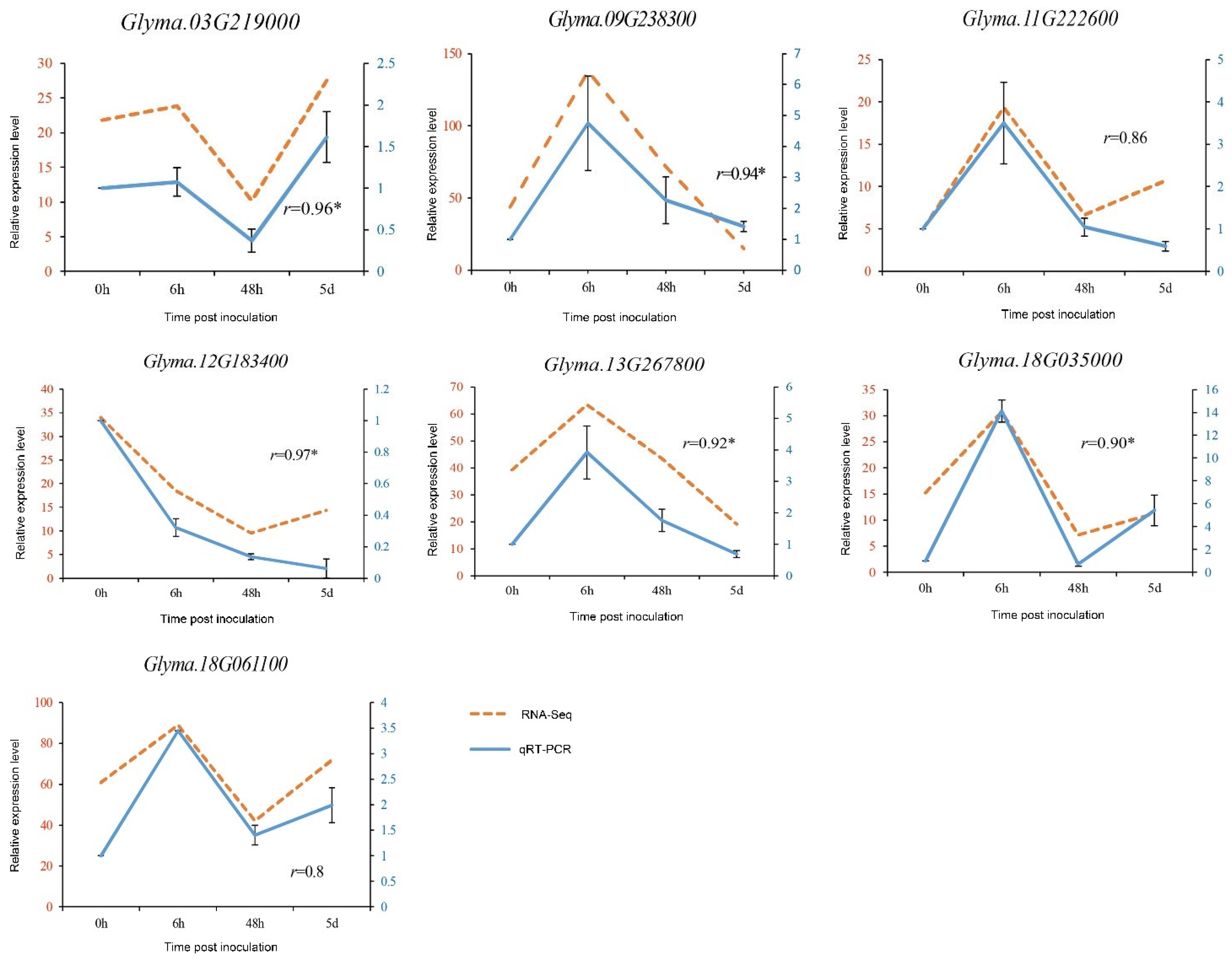
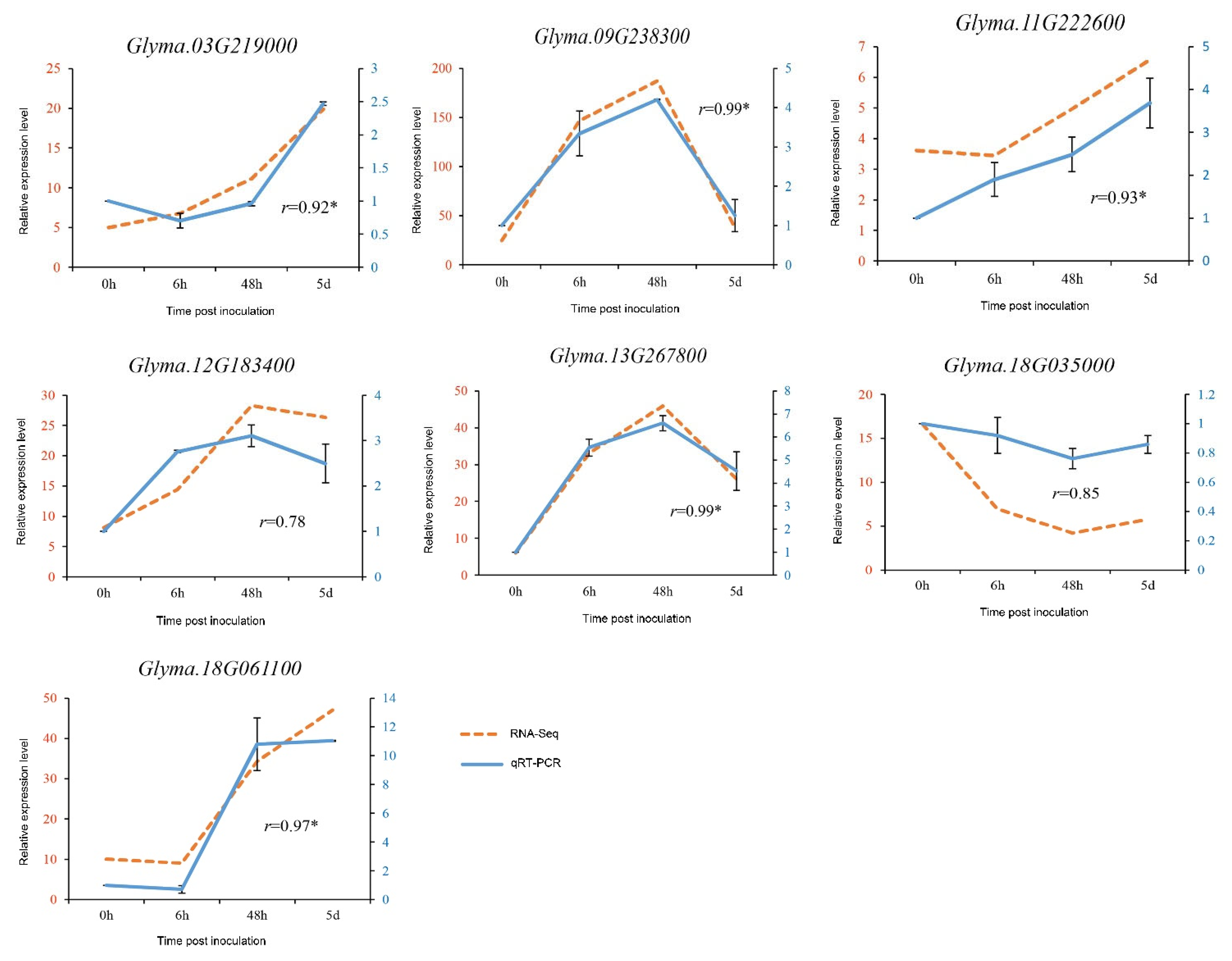
3.6. Validation of RNA-Seq Data by qRT-PCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, K.; Ren, R.; Wang, T.; Gao, L.; Luo, J.Y.; Liu, S.C.; Zhi, H.J.; Gai, J.Y. Genetic and genetic localization of resistance to soybean mosaic virus SC18 strains in soybean. Soybean Sci. 2017, 36, 187–192. [Google Scholar] [CrossRef]
- Revers, F.; García, J.A. Molecular biology of potyviruses. Adv. Virus Res. 2015, 92, 101–199. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.K.; Goodman, R.M. Strains of Soybean mosaic virus: Classification based on virulence in resistant soybean variety. Phytopathology 1979, 69, 467–490. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanaka, T.; Iida, W.; Tsuda, Y. Studies on virus diseases and causal viruses of soybean in Japan. Bull. Tohoku Natl. Agric. Exp. Stn. 1980, 62, 1–130. [Google Scholar]
- Seo, J.K.; Lee, H.G.; Choi, H.S.; Lee, S.H.; Kim, K.H. Infectious in vivo transcripts from a full-length clone of Soybean mosaic virus strain G5H. Plant Pathol. J. 2009, 25, 54–61. [Google Scholar] [CrossRef]
- Wang, X.Q.; Gai, J.Y.; Pu, Z.Q. Identification and distribution of soybean Mosaic virus strains in Huang-Huai region and the middle and lower reaches of Yangtze River. Soybean Sci. 2003, 22, 102–107. [Google Scholar]
- Zhan, Y.; Zhi, H.J.; Yu, D.Y.; Gai, J.Y. Identification and distribution of soybean Mosaic virus strains in Huang-Huai region. Agric. Sci. China 2006, 39, 2009–2015. [Google Scholar]
- Li, K.; Yang, Q.H.; Zhi, H.J.; Gai, J.Y. Identification and Distribution of Soybean mosaic virus Strains in Southern China. Plant Dis. 2010, 94, 351–357. [Google Scholar] [CrossRef]
- Wang, D.G.; Li, H.W.; Zhi, H.J.; Tian, Z.; Hu, C.; Hu, G.Y.; Huang, Z.P.; Zhang, L. Identification of strains and screening of resistance resources to Soybean mosaic virus in Anhui Province. Chin. J. Oil. Crop Sci. 2014, 36, 374–379. [Google Scholar] [CrossRef]
- Jeong, S.C.; Kristipati, S.; Hayes, A.J.; Maughan, P.J.; Noffsinger, S.L.; Gunduz, I.; Buss, G.R.; Maroof, M.A.S. Genetic and Sequence Analysis of Markers Tightly Linked to the Soybean mosaic virus Resistance Gene, Rsv3. Crop Sci. 2002, 42, 265–270. [Google Scholar] [CrossRef]
- Jeong, S.C.; Saghai, M.M.A. Detection and genotyping of SNPs tightly linked to two disease resistance loci, Rsv1 and Rsv3, of soybean. Plant Breed. 2004, 123, 305–310. [Google Scholar] [CrossRef]
- Hayes, A.J.; Ma, G.; Buss, G.R.; Maroof, M.A. Molecular Marker Mapping of RSV4, a Gene Conferring Resistance to all Known Strains of Soybean Mosaic Virus. Crop Sci. 2000, 40, 1434–1437. [Google Scholar] [CrossRef]
- Maroof, M.A.S.; Tucker, D.M.; Skoneczka, J.A.; Bowman, B.C.; Tripathy, S.; Tolin, S.A. Fine mapping and candidate gene discovery of the soybean mosaic virus resistance gene, Rsv4. Plant Genome 2010, 3, 14–32. [Google Scholar] [CrossRef]
- Gunduz, I.; Buss, G.R.; Chen, P.; Tolin, S.A. Characterization of SMV Resistance Genes in Tousan 140 and Hourei Soybean. Crop Sci. 2002, 42, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Buss, G.R.; Tolin, S.A. Resistance to soybean mosaic virus conferred by two independent dominant genes in PI486355. J. Hered. 1993, 84, 25–28. [Google Scholar] [CrossRef]
- Ma, G.; Chen, P.; Buss, G.R.; Tolin, S.A. Genetic characteristics of two genes for resistance to soybean mosaic virus in PI486355 soybean. Theor. Appl. Genet. 1995, 91, 907–914. [Google Scholar] [CrossRef]
- Ma, G.; Chen, P.; Buss, G.R.; Tolin, S.A. Complementary Action of Two Independent Dominant Genes in Columbia Soybean for Resistance to Soybean Mosaic Virus. J. Hered. 2002, 93, 179–184. [Google Scholar] [CrossRef]
- Gunduz, I.; Buss, G.R.; Chen, P.; Tolin, S.A. Genetic and Phenotypic Analysis of Soybean mosaic virus Resistance in PI 88788 Soybean. Phytopathology 2004, 94, 687–692. [Google Scholar] [CrossRef]
- Maroof, M.A.S.; Jeong, S.C.; Gunduz, I.; Tucker, M.; Buss, G.R.; Tolin, S.A. Pyramiding of Soybean Mosaic Virus Resistance Genes by Marker Assisted Selection. Crop Sci. 2008, 48, 517–526. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, D.G.; Li, H.C.; Zheng, G.J.; Yang, Y.Q.; Zhi, H.J. Fine mapping of the RSC14Q locus for resistance to soybean mosaic virus. Euphytica 2011, 181, 127–135. [Google Scholar] [CrossRef]
- Wang, D.G.; Ma, Y.; Liu, N.; Yang, Z.L.; Zheng, G.J.; Zhi, H.J. Fine mapping and identification of the soybean Rsc4 resistance candidate gene to soybean mosaic virus. Plant Breed. 2011, 130, 653–659. [Google Scholar] [CrossRef]
- Zheng, G.J.; Yang, Y.Q.; Ma, Y.; Yang, X.F.; Chen, S.Y.; Ren, R. Fine Mapping and Candidate Gene Analysis of Resistance Gene RSC3Q to Soybean mosaic virus in Qihuang-1. J. Integr. Agric. 2014, 130, 2608–2615. [Google Scholar] [CrossRef]
- Ren, R.; Liu, S.C.; Karthikeyan, A.; Wang, T.; Niu, H.P.; Yin, J.L.; Yang, Y.H.; Wang, L.Q.; Yang, Q.H.; Zhi, H.J.; et al. Fine-mapping and identification of a novel locus Rsc15 underlying soybean resistance to Soybean mosaic virus. Theor. Appl. Genet. 2017, 130, 2395–2410. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Gusberti, M.; Gessler, C.; Broggini, G.A. RNA-Seq analysis reveals candidate genes for ontogenic resistance in Malus-Venturia pathosystem. PLoS ONE 2017, 8, e78457. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.Q.; Zhang, K.; Yang, J.; Zhang, N.; Fang, A.F.; Zhang, Y.; Liu, Y.F.; Chen, Z.Y.; Tom, H.; Sun, W.X. Differential expression profiling of the early response to Ustilaginoidea virens between false smut resistant and susceptible rice varieties. BMC Genom. 2015, 16, 995. [Google Scholar] [CrossRef] [PubMed]
- Nanoth, V.N.; Wang, H.; Eyun, S.I.; Moriyama, E.N.; Coates, B.S.; Miller, N.J.; Siegfried, B.D. Transcriptional analysis of susceptible and resistant European corn borer strains and their response to Cry1F protoxin. BMC Genom. 2015, 16, 558. [Google Scholar] [CrossRef]
- Ocaña, S.; Seoane, P.; Bautista, R.; Palomino, C.; Claros, G.M.; Torres, A.M.; Madrid, E. Transcriptome Analysis in Faba Bean (Vicia faba L.) under Ascochyta fabae Infection. PLoS ONE 2015, 10, e0135143. [Google Scholar] [CrossRef]
- Zheng, W.J.; Ma, L.; Zhao, J.M.; Li, Z.Q.; Sun, F.Y.; Lu, X.C. Comparative transcriptome analysis of two rice varieties in response to rice stripe virus and small brown planthoppers during early interaction. PLoS ONE 2017, 8, e82126. [Google Scholar] [CrossRef]
- Yu, Y.; Shi, J.Y.; Li, X.Y.; Liu, J.; Geng, Q.; Shi, H.C.; Ke, Y.P.; Sun, Q. Transcriptome analysis reveals the molecular mechanisms of the defense response to gray leaf spot disease in maize. BioMed Cent. 2018, 19, 742. [Google Scholar] [CrossRef]
- Yuan, Y.; Yang, Y.Q.; Yin, J.L.; Shen, Y.C.; Li, B.W.; Wang, L.Q.; Zhi, H.J. Transcriptome-based discovery of genes and networks related to RSC3Q-mediated resistance to Soybean mosaic virus in soybean. Crop Pasture Sci. 2020, 71, 987–995. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Liao, L.; Sun, D.M. Screening for resistant of soybean germplasm to SMV. Jilin Agric. Sci. 1997, 1, 30–34. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Costa, V.; Angelini, C.; De, F.I.; Ciccodicola, A.; Xiong, M. Uncovering the Complexity of Transcriptomes with RNA-Seq. J. Biomed. Biotechnol. 2010, 2010, 853916. [Google Scholar] [CrossRef]
- Brian, T.W.; Samuel, M.; Stephen, W.; Falk, S.; Valerie, W.; Ian, G.; Christopher, J.P.; Jane, R.; Jürg, B. Dynamic repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution. Nat. Int. Wkly. J. Sci. 2008, 453, 1239–1243. [Google Scholar] [CrossRef]
- Zhong, W.; Mark, G.; Michael, S. RNA-Seq: A revolutionary tool for transcriptomic. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Narnia 2008, 36, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.P.; Li, D.Q. MAPK and reactive oxygen species participate in signal transduction of plant disease resistance defense response. Plant Physiol. Commun. 2009, 45, 73–79. [Google Scholar]
- Yanagisawa, S.; Yoo, S.D.; Sheen, J. Differential regulation of EIN3 stability by glucose and ethylene signalling in plants. Nature 2003, 425, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Jyoti, S.D.; Azim, J.B.; Robin, A.H.K. Genome-wide characterization and expression profiling of EIN3/EIL family genes in Zea mays. Plant Gene 2021, 25, 100270. [Google Scholar] [CrossRef]
- Alazem, M.; Lin, N.S. Roles of plant hormones in the regulation of host–virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef]
- Lee, M.W.; Qi, M.; Yang, Y.A. Novel jasmonic acid-inducible rice myb gene associates with fungal infection and host cell death. Mol. Plant-Microbe Interact. 2001, 14, 527–535. [Google Scholar] [CrossRef]
- Penninckx, I.A.; Eggermont, K.; Terras, F.R.; Thomma, B.P.; De, S.G.W.; Buchala, A.M.J.P.; Manners, J.M.; Broekaert, W.F. Pathogen-induced systemic activation of a plant defensin gene in Arabidopsis follows a salicylic acid-independent pathway. Plant Cell 1996, 8, 2309–2323. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J.; Eggermont, K.; Penninckx, I.A.M.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.A.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef]
- Vijayan, P.; Shockey, J.; Andre, L.C. A role for jasmonate in pathogen defense of Arabi-dopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 7209–7214. [Google Scholar] [CrossRef]
- Xie, D.X.; Feys, B.F.; James, S.; Nieto, R.M.; Turner, J.G. Col1: An Arabidopsis gene required for jasmonate-regulated defense and fertility. Science 1998, 280, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Lu, X.; Zhang, L.; Gao, E.D.; Tang, K.X. Advances in the function of MYC2 transcription factor in plants. J. Shanghai Jiaotong Univ. Agric. Sci. 2012, 30, 51–57. [Google Scholar]
- Oka, K.; Kobayashi, M.; Mitsuhara, I.; Seo, S. Jasmonic acid negativelyregulates resistance to Tobacco mosaic virus in tobacco. Plant Cell Physiol. 2013, 54, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Alazem, M.; Tseng, K.C.; Chang, W.C.; Seo, J.K.; Kim, K.H. Elements involved in the Rsv3-mediated extreme resistance against an avirulent strain of soybean mosaic virus. Viruses 2018, 10, 581. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.M.; Luo, D.P.; Cao, Q.L.; Feng, J.X.; Duan, C.J. Resistance and agronomic traits of 253 T3 rice lines with NPR1 gene to bacterial blight of rice. J. South. Agric. 2012, 43, 417–420. [Google Scholar]
- Pieterse, C.M.J.; Leon Reyes, A.; Van, D.E.S.; Wees, S.C.M.V. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef]
- Wang, L.M.; Lin, D.; Zhai, X.Q. Advances in MAPK signal transduction pathways in plants. Henan For. Sci. Technol. 2019, 39, 17–20. [Google Scholar]
- Lam, H.M.; Wong, P.; Chan, H.K.; Chen, L.; Chow, C.M.; Coruzzi, G.M. Overexpression of the ASN1 Gene Enhances Nitrogen Status in Seeds of Arabidopsis. Plant Physiol. 2003, 132, 926–935. [Google Scholar] [CrossRef]
- Castillo, L.; Díaz, M.; González, C.A.; González, A.; Alonso, P.E.; Bassagoda, M.J.; Rossini, C. Clytostoma callistegioides (Bignoniaceae) wax extract with activity on aphid settling. Phytochemistry 2010, 71, 2052–2057. [Google Scholar] [CrossRef]
- Hansjakob, A.; Bischof, S.; Bringmann, G.; Riederer, M.; Hildebrandt, U. Very-long-chain aldehydes promote in vitro prepenetration processes of Blumeria graminis in a dose- and chain length-dependent manner. New Phytol. 2010, 188, 1039–1054. [Google Scholar] [CrossRef]
- Wen, Z.X.; Tan, R.J.; Zhang, S.C.; Collins, P.J.; Yuan, J.Z.; Du, W.Y.; Gu, C.H.; Ou, S.J.; Song, Q.J.; Charles, Y.Q.; et al. Integrating GWAS and gene expression data for functional characterization of resistance to white mould in soyabean. Plant Biotechnol. J. 2018, 16, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Whaley, A.; Sheridan, J.; Safari, S.; Burton, A.; Burkey, K.; Schlueter, J. RNA-seq analysis reveals genetic response and tolerance mechanisms to ozone exposure in soybean. BMC Genom. 2015, 16, 426. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Description | Symbol | log2FC (NN-48 h/0 h) | log2FC (NN-5d/0 h) | log2FC (KF-48 h/0 h) | log2FC (KF-5d/0 h) | Pathway |
|---|---|---|---|---|---|---|---|
| Glyma.11G239000 | putative ETHYLENE INSENSITIVE 3-like 4 protein (Glycine max(soybean)) | EIN/EIL | −7.4 | −7.4 | - | - | ETH |
| Glyma.14G041500 | ETHYLENE INSENSITIVE 3-like 1 protein (Glycine max(soybean)) | - | - | - | 1.8 | ||
| Glyma.18G018400 | putative ETHYLENE INSENSITIVE 3-like 4 protein (Glycine max(soybean)) | - | −4.5 | - | - | ||
| MSTRG.38091 | the new gene | ETR | 2.5 | 2.5 | - | - | |
| MSTRG.38100 | the new gene | −2.4 | - | - | - | ||
| Glyma.09G002600 | ethylene receptor (Glycine max(soybean)) | 1.1 | 2 | - | 1.1 | ||
| Glyma.12G241700 | ethylene receptor (Glycine max(soybean)) | 1.2 | - | - | - | ||
| Glyma.19G213300 | ethylene response sensor 1 (Glycine max(soybean)) | 1.1 | - | - | - | ||
| Glyma.09G077500 | uncharacterized LOC100306045 (Glycine max(soybean)) | TIFY/JAZ | - | −1.1 | - | - | JA |
| Glyma.13G116100 | CCT motif and tify domain containing protein (Glycine max(soybean)) | - | - | - | 1.4 | ||
| Glyma.17G043700 | CCT and tify domains containing protein (Glycine max(soybean)) | −1.1 | −1.7 | - | - | ||
| Glyma.01G096600 | transcription factor MYC2 (Glycine max(soybean)) | MYC2 | −1.3 | 11.1 | 11.4 | - | |
| Glyma.07G051500 | transcription factor MYC2 (Glycine max(soybean)) | −2 | - | - | 1.4 | ||
| Glyma.14G117300 | uncharacterized LOC100775685 (Glycine max(soybean)) | - | 2.4 | 1.2 | 2.9 | ||
| Glyma.03G128600 | BTB/POZ domain and ankyrin repeat containing protein COCH | NPR1 | - | 1.8 | 1.3 | - | SA |
| Glyma.09G020800 | NPR1-1 protein (Glycine max(soybean)) | −1 | - | - | - | ||
| Glyma.14G031300 | BTB/POZ domain and ankyrin repeat containing protein NPR1 (Glycine max(soybean)) | - | - | - | 1.1 | ||
| Glyma.19G131000 | regulatory protein NPR5-like (Glycine max(soybean)) | - | 2.5 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Shen, Y.; Chen, B.; Xie, L.; Xiao, Y.; Chong, Z.; Cai, H.; Xing, G.; Zhi, H.; Li, K. Comparative Transcriptome Analyses between Resistant and Susceptible Varieties in Response to Soybean Mosaic Virus Infection. Agronomy 2022, 12, 1785. https://doi.org/10.3390/agronomy12081785
Chen Y, Shen Y, Chen B, Xie L, Xiao Y, Chong Z, Cai H, Xing G, Zhi H, Li K. Comparative Transcriptome Analyses between Resistant and Susceptible Varieties in Response to Soybean Mosaic Virus Infection. Agronomy. 2022; 12(8):1785. https://doi.org/10.3390/agronomy12081785
Chicago/Turabian StyleChen, Yuanyuan, Ying Shen, Boyu Chen, Lijun Xie, Yanmin Xiao, Zheng Chong, Han Cai, Guangnan Xing, Haijian Zhi, and Kai Li. 2022. "Comparative Transcriptome Analyses between Resistant and Susceptible Varieties in Response to Soybean Mosaic Virus Infection" Agronomy 12, no. 8: 1785. https://doi.org/10.3390/agronomy12081785
APA StyleChen, Y., Shen, Y., Chen, B., Xie, L., Xiao, Y., Chong, Z., Cai, H., Xing, G., Zhi, H., & Li, K. (2022). Comparative Transcriptome Analyses between Resistant and Susceptible Varieties in Response to Soybean Mosaic Virus Infection. Agronomy, 12(8), 1785. https://doi.org/10.3390/agronomy12081785