
Genome-Wide Identification and Characterization of the Abiotic-Stress-Responsive LACS Gene Family in Soybean (Glycine max)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of the LACS Gene Family in Soybean
2.2. Analysis of Gene Sequence and Physicochemical Properties
2.3. Phylogenetic Tree Construction and Protein Conserved Motif Analysis
2.4. Gene Location on Chromosome and Gene Structure of GmLACSs
2.5. Collinearity Analysis of GmLACSs
2.6. Promoter Analysis of GmLACSs
2.7. Expression Profiles of GmLACSs in Diverse Tissues
2.8. Quantitative Real-Time RT-PCR Analysis
2.9. Co-Expression of GmLACSs and Transcription Factors under Drought Stresses
2.10. Statistical Analysis
3. Results
3.1. Identification Sequence Analysis and Physicochemical Properties Analysis of LACS Gene Family in Soybean
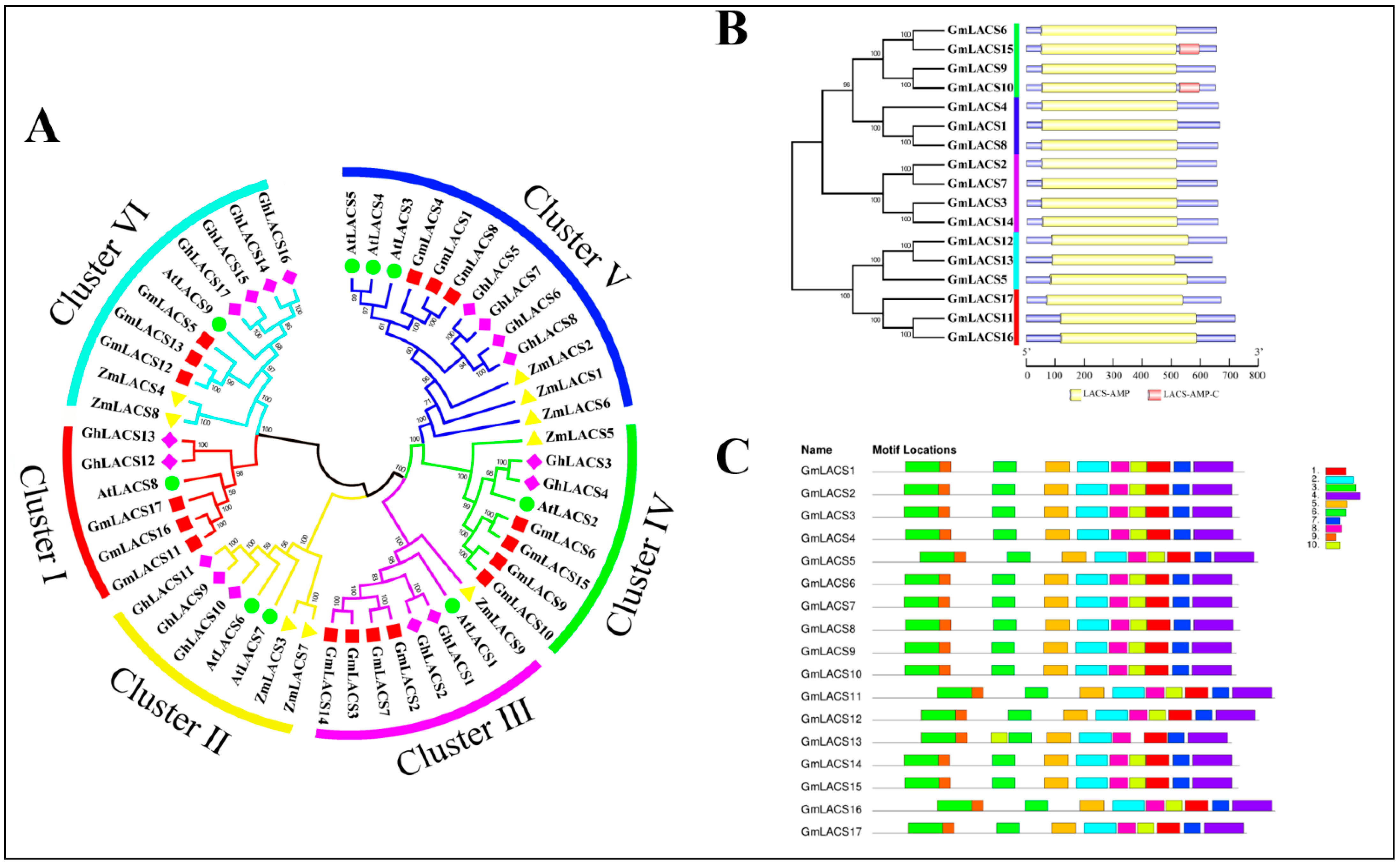
3.2. Phylogenetic, Protein Domain and Protein Conserved Motifs Analysis of GmLACSs
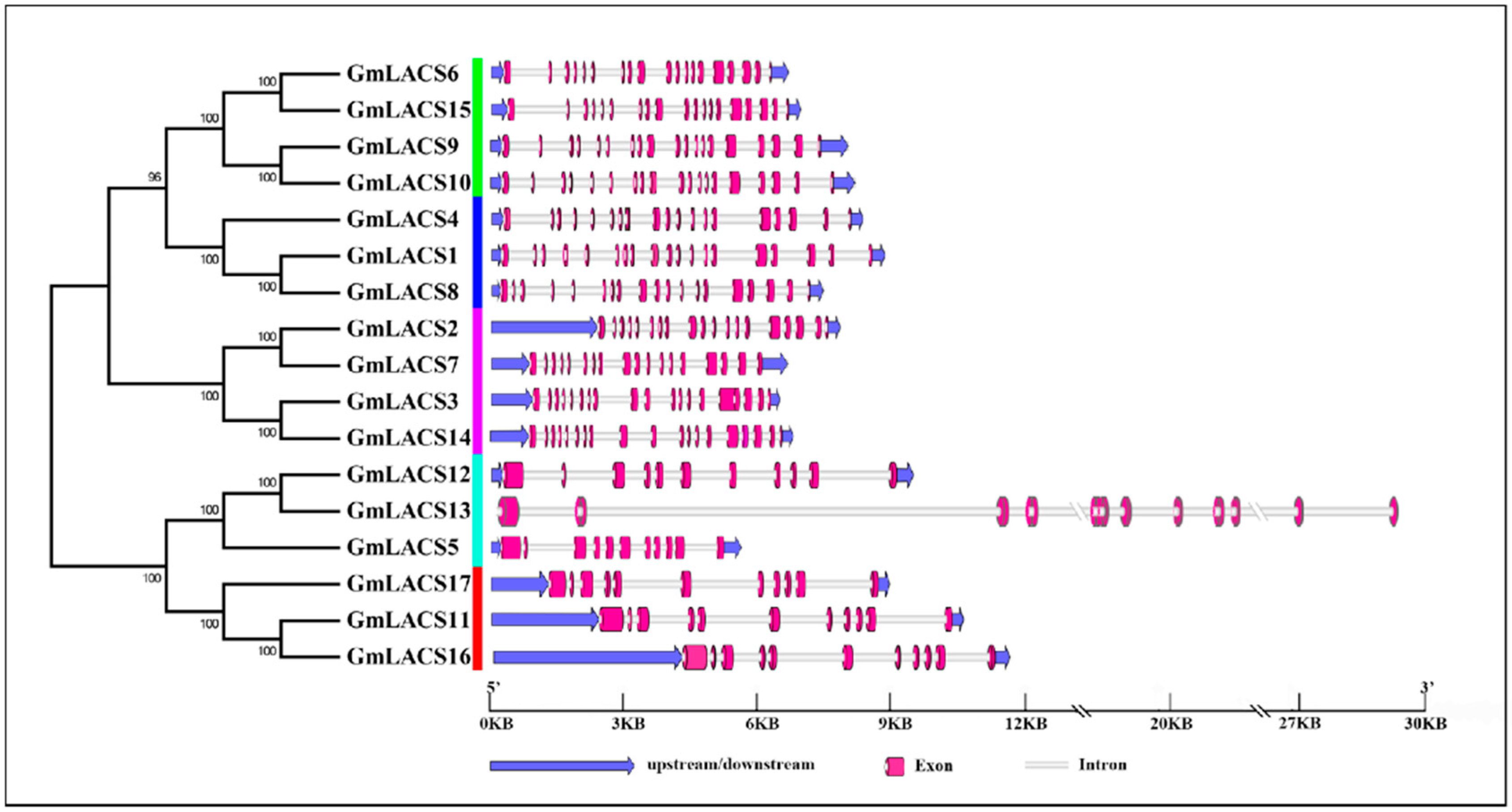
3.3. Gene Location Analysis and Gene Structure Analysis of GmLACSs
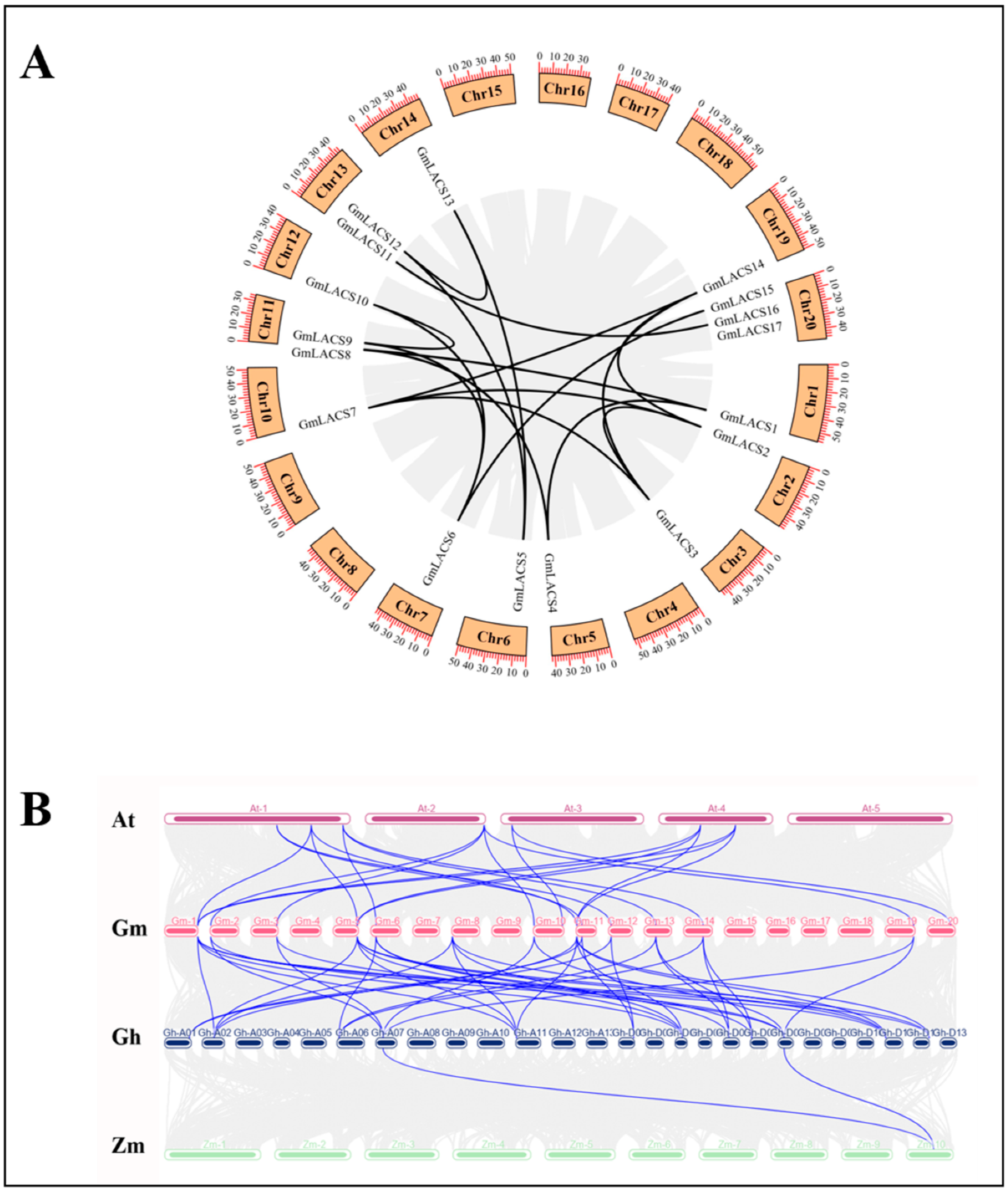
3.4. Collinearity Relation of GmLACSs
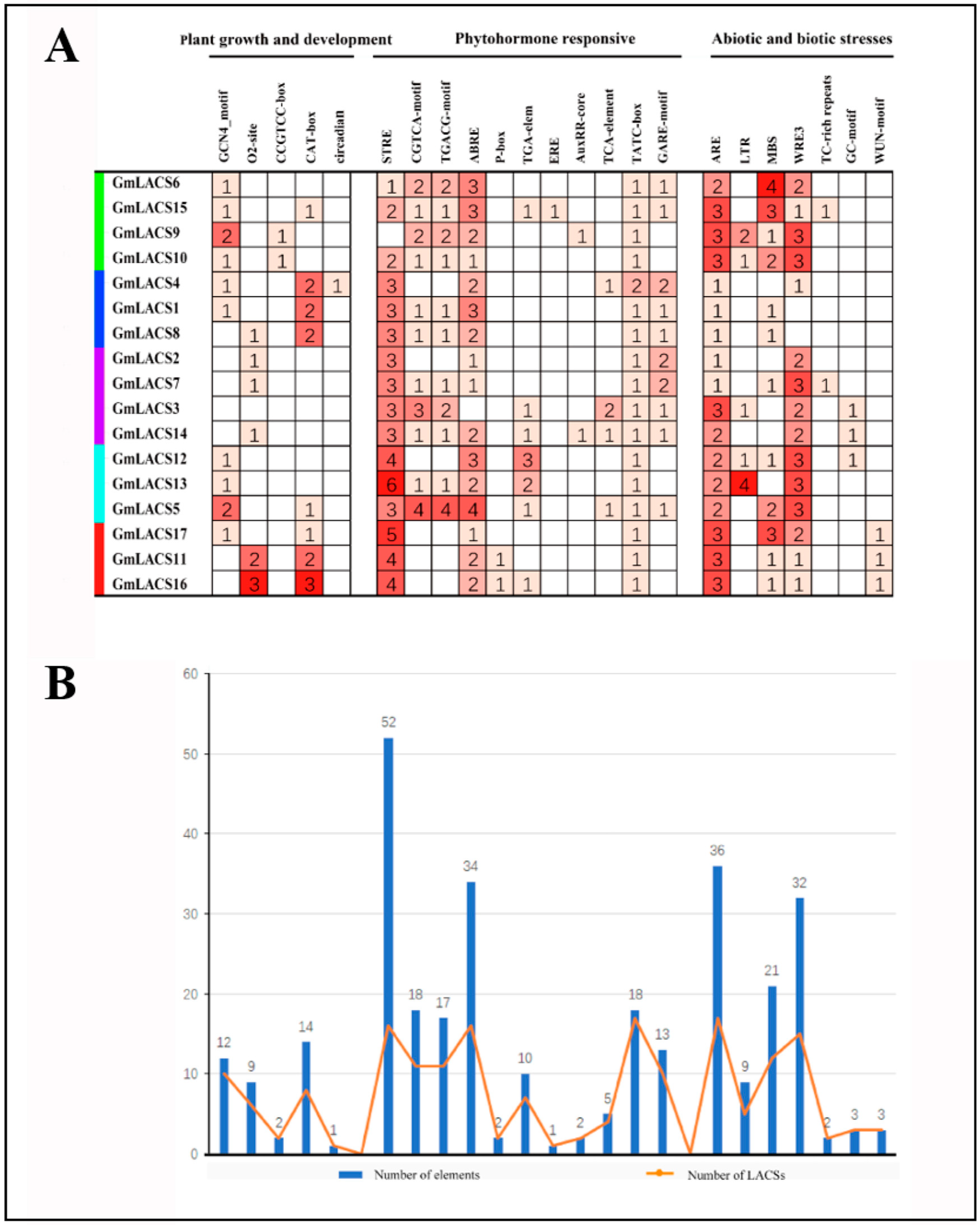
3.5. Identification of Cis-Acting Regulatory Elements in the Promoter of GmLACS Genes
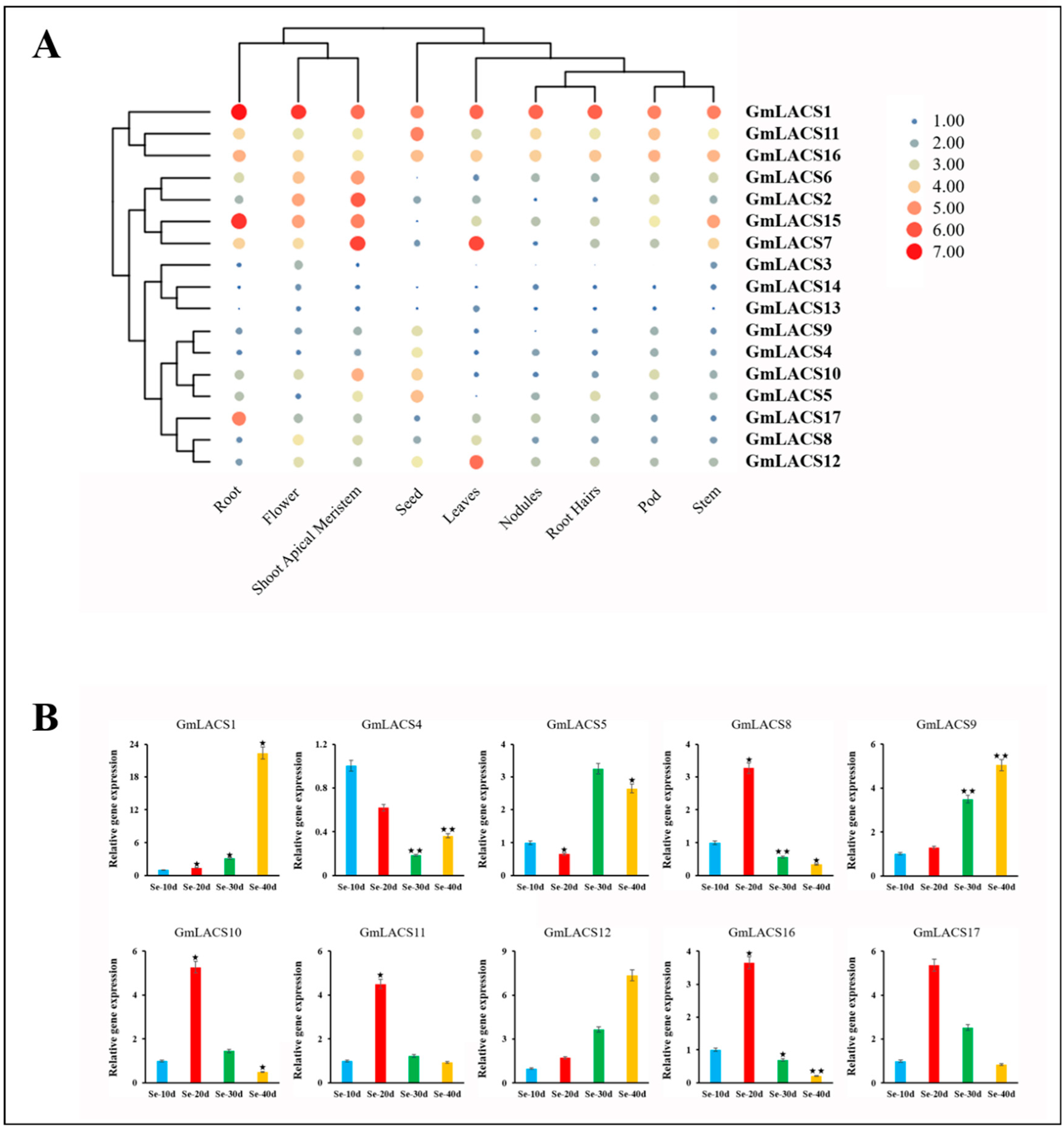
3.6. Expression Profiles of GmLACSs in Different Tissues and Developmental Phases
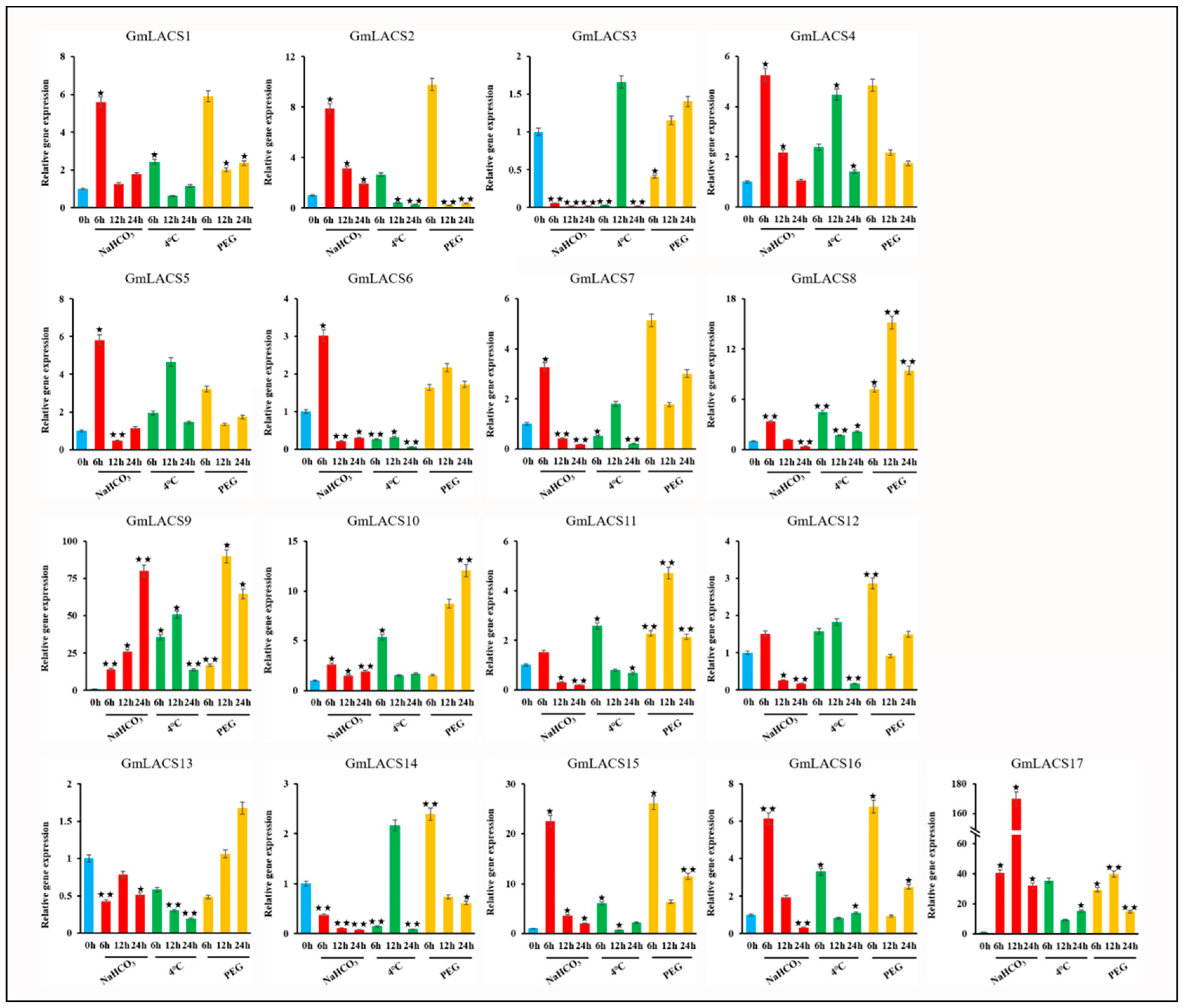
3.7. Transcript Level of GmLACSs under Abiotic Stress
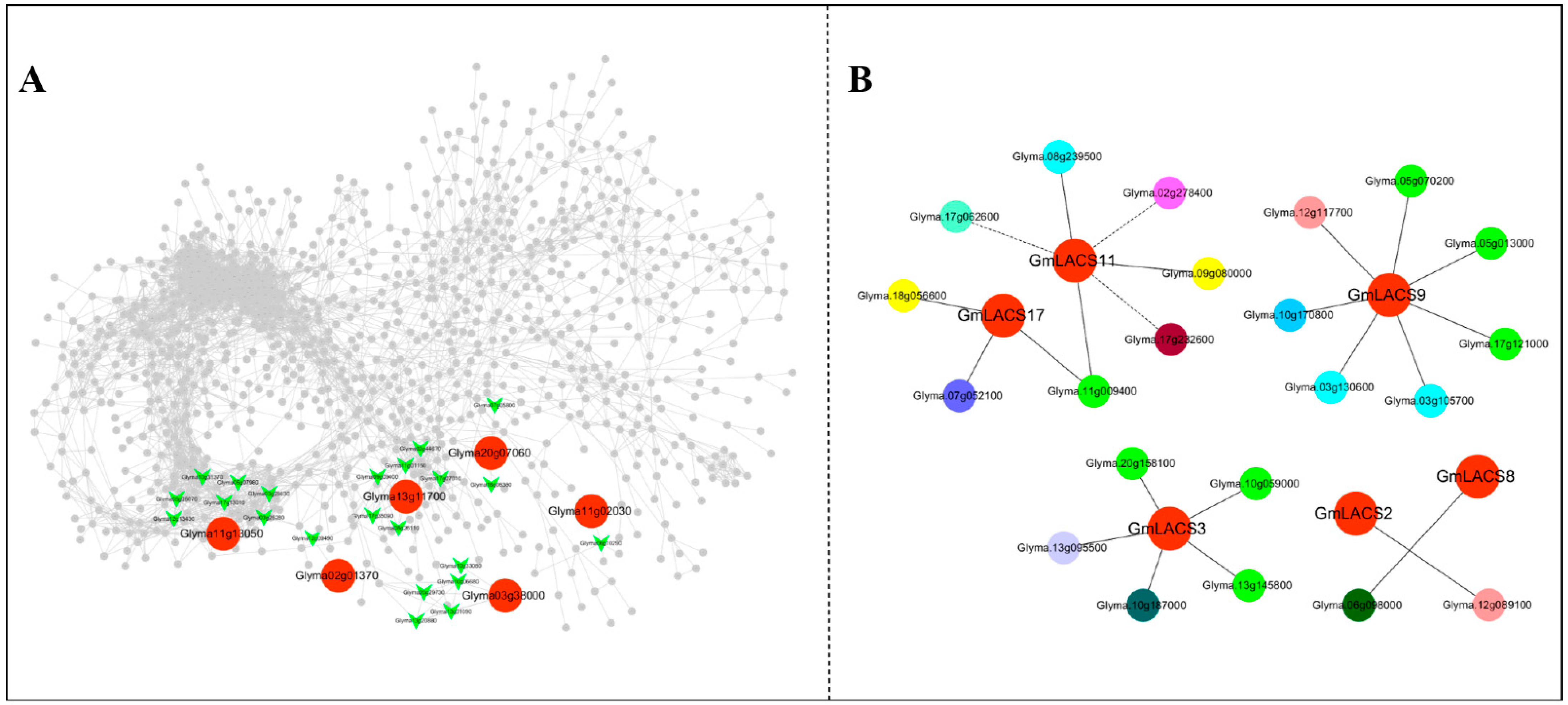
3.8. Co-Expression Analysis of Transcription Factors and GmLACSs in Soybean
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karantonis, H.C.; Nomikos, T.; Demopoulos, C.A. Triacylglycerol Metabolism. Curr. Drug Targets 2009, 10, 302–319. [Google Scholar] [CrossRef] [PubMed]
- Brash, A.R.; Ingram, C.D.; Harris, T.M. Analysis of a specific oxygenation reaction of soybean lipoxygenase-1 with fatty acids esterified in phospholipids. Biochemistry 1987, 26, 5465–5471. [Google Scholar] [CrossRef] [PubMed]
- Tonon, T.; Qing, R.; Harvey, D.; Li, Y.; Larson, T.R.; Graham, I.A. Identification of a long-chain polyunsaturated fatty acid acyl-coenzyme a synthetase from the diatom thalassiosira pseudonana. Plant Physiol. 2005, 138, 402–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fich, E.A.; Segerson, N.A.; Rose, J.K. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.F.; Haslam, T.M.; Sonntag, A.; Molina, I.; Kunst, L. Functional overlap of long-chain acyl-CoA synthetases in Arabidopsis. Plant Cell Physiol. 2019, 60, 1041–1054. [Google Scholar] [CrossRef]
- Hills, M.J.; Beevers, H. ATPase in lipid body membranes of castor bean endosperm. Plant Physiol. 1986, 82, 671–674. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Yu, C.; Huang, A.H. Substrate specificities of lipases from corn and other seeds. Arch. Biochem. Biophys. 1986, 244, 346–356. [Google Scholar] [CrossRef]
- Goepfert, S.; Poirier, Y. β-oxidation in fatty acid degradation and beyond. Curr. Opin. Plant Biol. 2007, 10, 245–251. [Google Scholar] [CrossRef]
- Eastmond, P.J.; Germain, V.; Lange, R.P.; Bryce, J.H.; Smith, S.M.; Graham, I.A. Postgerminative growth and lipid catabolism in oilseeds lacking the glyoxylate cycle. Proc. Natl. Acad. Sci. USA 2000, 97, 5669–5674. [Google Scholar] [CrossRef] [Green Version]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Debono, A.; Durrett, T.P.; Franke, R.B.; Graham, I.A.; et al. Acyl-lipid metabolism. Arab. Book 2010, 8, e0133. [Google Scholar] [CrossRef] [Green Version]
- Hills, M.J.; Murphy, D.J.; Beevers, H. Inhibition of neutral lipase from castor bean lipid bodies by coenzyme A (CoA) and oleoyl-CoA. Plant Physiol. 1989, 89, 1006–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beevers, H. Glyoxysomes of castor bean endosperm and their relation to gluconeogenesis. Ann. N. Y. Acad. Sci. 2010, 168, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Groot, P.H.; Scholte, H.R.; Hlsmann, W.C. Fatty acid activation: Specificity, localization, and function. Adv. Lipid Res. 1976, 14, 75. [Google Scholar]
- Tan, X.L.; Zheng, X.F.; Zhang, Z.Y.; Wang, Z.; Xia, H.C.; Chang-Ming, L.U.; Shou-Lai, G.U. Acyl-coenzyme a synthetase 4 (BnLACS4) gene, which enhances the yeast lipid contents. J. Integr. Agric. 2014, 13, 54–62. [Google Scholar] [CrossRef]
- Chen, H.; Yu, C.E.; Sun, R.H.; Li, D.D.; Zheng, Y.S. Cloning and yeast expression of long-chain fatty Acyl-Coenzyme a synthetase 1 (LACS1) gene from Arabidopsis. Mol. Plant Breed. 2017, 15, 1623–1629. [Google Scholar]
- Fulda, M.; Shockey, J.; Werber, M.; Wolter, F.P.; Heinz, E. Two long-chain acyl-CoA synthetases from Arabidopsis thaliana involved in peroxisomal fatty acid β-oxidation. Plant 2002, 32, 93–103. [Google Scholar]
- Lu, S.Y.; Song, T.; Kosma, D.K.; Parsons, E.P.; Jenks, M.A. Arabidopsis CER8 encodes long-chain acyl CoA synthetase 1 (LACS1) and has overlapping functions with LACS2 in plant wax and cutin synthesis. Plant J. 2009, 59, 553–564. [Google Scholar] [CrossRef]
- Shockey, J.; Browse, J. Genome-level and biochemical diversity of the acyl-activating enzyme superfamily in plants. Plant J. Cell Mol. Biol. 2011, 66, 143. [Google Scholar] [CrossRef]
- Weng, H.; Molina, I.; Shockey, J.; Browse, J. Organ fusion and defective cuticle function in a lacs1 lacs2 double mutant of Arabidopsis. Planta 2010, 231, 1089–1100. [Google Scholar] [CrossRef]
- Shockey, J.M.; Fulda, M.S.; Browse, J.A. Arabidopsis contains nine long-chain acyl-coenzyme a synthetase genes that participate in fatty acid and glycerolipid metabolism. Plant Physiol. 2002, 129, 1710–1722. [Google Scholar] [CrossRef] [Green Version]
- Jessen, D.; Roth, C.; Wiermer, M.; Fulda, M. Two activities of long-chain acyl-coenzyme a synthetase are involved in lipid trafficking between the endoplasmic reticulum and the plastid in Arabidopsis. Plant Physiol. 2015, 167, 351–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, S.Y.; Mashek, D.G. Hepatic long-chain acyl-CoA synthetase 5 mediates fatty acid channeling between anabolic and catabolic pathways. J. Lipid Res. 2010, 51, 3270–3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digel, M.; Ehehalt, R.; Stremmel, W.; Fullekrug, J. Acyl-CoA synthetases: Fatty acid uptake and metabolic channeling. Mol. Cell. Biochem. 2009, 326, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Schnurr, J.A.; Shockey, J.M.; Boer, G.J.; Browse, J.A. Fatty acid export from the chloroplast. Molecular characterization of a major plastidial acyl-coenzyme a synthetase from Arabidopsis. Plant Physiol. 2002, 129, 1700–1709. [Google Scholar] [CrossRef] [Green Version]
- Fulda, M.; Schnurr, J.A.; Abbadi, A.; Heinz, E.; Browse, J. Peroxisomal acyl-CoA synthetase activity is essential for seedling development in Arabidopsis thaliana. Plant Cell 2004, 16, 394–405. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.L.; Li, X.B. The GhLACS1 gene encodes an acyl-CoA synthetase which is essential for normal microsporogenesis in early anther development of cotton. Plant J. 2010, 57, 473–486. [Google Scholar] [CrossRef]
- He, X.; Chen, G.Q.; Kang, S.T.; Mckeon, T.A. Ricinus communis contains an acyl-coa synthetase that preferentially activates ricinoleate to its CoA thioester. Lipids 2007, 42, 931. [Google Scholar] [CrossRef]
- Liu, X.; He, Z.; Mao, L.; Wang, J. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature 2013, 496, 91–95. [Google Scholar]
- Ichihara, K.; Kobayashi, N.; Saito, K. Lipid synthesis and acyl-CoA synthetase in developing rice seeds. Lipids 2003, 38, 881–884. [Google Scholar] [CrossRef]
- Zhang, C.L.; Mao, K.; Zhou, L.J.; Wang, G.L.; Zhang, Y.L.; Li, Y.Y.; Hao, Y.J. Genome-wide identification and characterization of apple long-chain acyl-CoA synthetases and expression analysis under different stresses. Plant Physiol. Biochem. 2018, 132, 320–332. [Google Scholar] [CrossRef]
- Xiao, Z.; Li, N.; Wang, S.; Sun, J.; Zhang, L.; Zhang, C.; Yang, H.; Zhao, H.; Yang, B.; Wei, L.; et al. Genome-wide identification and comparative expression profile analysis of the long-chain acyl-CoA synthetase (LACS) gene family in two different oil content cultivars of brassica napus. Biochem. Genet. 2019, 57, 781–800. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.S.; Zhao, L.M.; Liu, B.; Wang, Z.W.; Jin, Z.Q.; Sun, H. The genetic diversity of cultivated soybean grown in China. Theor. Appl. Genet. 2004, 108, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Ehirim, B.O. Improving Soybean Productivity Using Biotechnology Approach in Nigeria. World J. Agric. Sci. 2014, 2, 13–18. [Google Scholar]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2015, D1, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Robert, F.; Penelope, C.; Ruth, E.; Sean, E.; Jaina, M.; Alex, M.; Simon, P.; Marco, P.; Matloob, Q.; Amaia, S.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, D1, D279–D285. [Google Scholar]
- Peng, N.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; Xue, Y.; Ren, J. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 20, 3359. [Google Scholar]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Zhang, T.Z.; Hu, Y.; Chen, Z.J. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Rui, X.; Hao, C.; He, Y. TBtools, a Toolkit for Biologists Integrating Various HTS-Data Handling Tools with a User-Friendly Interface. Mol. Plant. 2018, 13, 8. [Google Scholar]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belamkar, V.; Weeks, N.T.; Bharti, A.K.; Farmer, A.D.; Graham, M.A.; Cannon, S.B. Comprehensive characterization and RNA-seq profiling of the hd-zip transcription factor family in soybean (Glycine max) during dehydration and salt stress. BMC Genom. 2014, 15, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H. Molecular Cloning and Functional Analysis of GmLACS2-3 Reveals Its Involvement in Cutin and Suberin Biosynthesis along with Abiotic Stress Tolerance. Int. J. Mol. Sci. 2001, 22, 9175. [Google Scholar] [CrossRef] [PubMed]
- Washida, H.; Wu, C.Y.; Suzuki, A.; Yamanouchi, U.; Akihama, T.; Harada, K.; Takaiwa, F. Identification of cis-regulatory elements required for endosperm expression of the rice storage protein glutelin gene glub-1. Plant Mol. Biol. 1999, 40, 1–12. [Google Scholar] [CrossRef]
- Koichiro, T.; Glen, S.; Daniel, P.; Alan, F.; Sudhir, K. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar]
- Rouster, J.; Leah, R.; Mundy, J.; Cameron-Mills, V. Identification of a methyl jasmonate-responsive region in the promoter of a lipoxygenase 1 gene expressed in barley grain. Plant J. 2010, 11, 513–523. [Google Scholar] [CrossRef]
- Shen, Q.; Ho, T.H. Functional dissection of an abscisic acid (ABA)-inducible gene reveals two independent ABA-responsive complexes each containing a G-box and a novel cis-acting element. Plant Cell 1995, 7, 295–307. [Google Scholar]
- Kim, J.K.; Cao, J.; Wu, R. Regulation and interaction of multiple protein factors with the proximal promoter regions of a rice high pI alpha-amylase gene. Mol. Gen. Genet. 1992, 232, 383–393. [Google Scholar] [CrossRef]
- Ulmasov, T.; Murfett, J.; Hagen, G.; Guilfoyle, T. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 1997, 9, 1963–1971. [Google Scholar]
- Goldsbrough, A.P.; Albrecht, H.; Stratford, R. Salicylic acid-inducible binding of a tobacco nuclear protein to a 10 bp sequence which is highly conserved amongst stress-inducible genes. Plant J. Cell Mol. Biol. 2010, 3, 563–571. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, M.; Feng, F.; Tian, Z. Toward a “Green Revolution” for Soybean. Mol. Plant 2020, 13, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Islam, T. Genome-wide analysis and expression profiling of glyoxalase gene families in soybean (Glycine max) indicate their development and abiotic stress specific response. BMC Plant Biol. 2016, 16, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babbitt, P.C.; Kenyon, G.L.; Martin, B.M.; Charest, H.; Slyvestre, M.; Scholten, J.D.; Chang, K.H.; Liang, P.H.; Dunaway-Mariano, D. Ancestry of the 4-chlorobenzoate dehalogenase: Analysis of amino acid sequence identities among families of acyl: Adenyl ligases, enoyl-CoA hydratases/isomerases, and acyl-CoA thioesterases. Biochemistry 1992, 31, 5594–5604. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Zhang, Y.L.; Hu, X. An apple long-chain acyl-CoA synthetase, MdLACS4, induces early flowering and enhances abiotic stress resistance in Arabidopsis. Plant Sci. 2020, 297, 110529. [Google Scholar] [CrossRef]
- Zhao, L.; Katavic, V.; Li, F.; Haughn, G.W.; Kunst, L. Insertional mutant analysis reveals that long-chain acyl-CoA synthetase 1 (LACS1), but not LACS8, functionally overlaps with LACS9 in Arabidopsis seed oil biosynthesis. Plant J. 2010, 64, 1048–1058. [Google Scholar] [CrossRef]
- Bessire, M.; Chassot, C.; Jacquat, A.C.; Humphry, M.; Nawrath, C. A permeable cuticle in arabidopsis leads to a strong resistance to botrytis cinerea. EMBO J. 2007, 26, 2158–2168. [Google Scholar] [CrossRef]
- Schmidt, R.R.; Fulda, M.; Paul, M.V.; Anders, M.; Plum, F.; Weits, D.A.; Kosmacz, M.; Larson, T.R.; Graham, I.A.; Beemster, G.T.S.; et al. PNSA plus: Low-oxygen response is triggered by an ATP-dependent shift in oleoyl-CoA in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E12101–E12110. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Kosma, D.K.; Lu, S. Functional role of long-chain acyl-coa synthetases in plant development and stress responses. Front. Plant Sci. 2021, 12, 640996. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | CDS Length (bp) | Protein Length (aa) | Isoelectric Point (pI) | Molecular Weight (Da) |
|---|---|---|---|---|---|
| GmLACS1 | Glyma.01G225200 | 2016 | 672 | 6.88 | 75,365.95 |
| GmLACS2 | Glyma.02G010300 | 1983 | 661 | 6.54 | 74,382.04 |
| GmLACS3 | Glyma.03G221400 | 1989 | 663 | 6.78 | 74,409.94 |
| GmLACS4 | Glyma.05G216600 | 1998 | 666 | 6.10 | 74,670.93 |
| GmLACS5 | Glyma.06G112900 | 2085 | 695 | 8.24 | 76,017.50 |
| GmLACS6 | Glyma.07G161900 | 1983 | 661 | 6.20 | 73,852.74 |
| GmLACS7 | Glyma.10G010800 | 1983 | 661 | 6.73 | 74,516.20 |
| GmLACS8 | Glyma.11G017900 | 1992 | 664 | 7.76 | 74,031.44 |
| GmLACS9 | Glyma.11G122500 | 1971 | 657 | 6.23 | 73,788.63 |
| GmLACS10 | Glyma.12G047400 | 1971 | 657 | 5.58 | 73,843.48 |
| GmLACS11 | Glyma.13G010100 | 2178 | 726 | 7.83 | 79,550.53 |
| GmLACS12 | Glyma.13G079900 | 2091 | 697 | 5.91 | 76,131.58 |
| GmLACS13 | Glyma.14G149700 | 1944 | 648 | 5.90 | 70,974.48 |
| GmLACS14 | Glyma.19G218300 | 1989 | 663 | 6.50 | 74,643.20 |
| GmLACS15 | Glyma.20G007900 | 1983 | 661 | 6.39 | 73,831.71 |
| GmLACS16 | Glyma.20G060100 | 2178 | 726 | 6.87 | 79,700.80 |
| GmLACS17 | Glyma.20G060300 | 2025 | 675 | 6.53 | 74,317.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, X.; Zhao, X.; Na, C.; Liu, H.; Miao, H.; Zhou, J.; Xiao, J.; Zhao, X.; Han, Y. Genome-Wide Identification and Characterization of the Abiotic-Stress-Responsive LACS Gene Family in Soybean (Glycine max). Agronomy 2022, 12, 1496. https://doi.org/10.3390/agronomy12071496
Wang J, Li X, Zhao X, Na C, Liu H, Miao H, Zhou J, Xiao J, Zhao X, Han Y. Genome-Wide Identification and Characterization of the Abiotic-Stress-Responsive LACS Gene Family in Soybean (Glycine max). Agronomy. 2022; 12(7):1496. https://doi.org/10.3390/agronomy12071496
Chicago/Turabian StyleWang, Jie, Xiaoxue Li, Xunchao Zhao, Chen Na, Hongliang Liu, Huanran Miao, Jinghang Zhou, Jialei Xiao, Xue Zhao, and Yingpeng Han. 2022. "Genome-Wide Identification and Characterization of the Abiotic-Stress-Responsive LACS Gene Family in Soybean (Glycine max)" Agronomy 12, no. 7: 1496. https://doi.org/10.3390/agronomy12071496
APA StyleWang, J., Li, X., Zhao, X., Na, C., Liu, H., Miao, H., Zhou, J., Xiao, J., Zhao, X., & Han, Y. (2022). Genome-Wide Identification and Characterization of the Abiotic-Stress-Responsive LACS Gene Family in Soybean (Glycine max). Agronomy, 12(7), 1496. https://doi.org/10.3390/agronomy12071496