
Regulatory Role of Trehalose Metabolism in Cold Stress of Harmonia axyridis Laboratory and Overwinter Populations

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of Experimental Insects
2.2. Distribution of Elytra Color of H. axyridis in a Natural Overwintering Population
2.3. Low-Temperature Storage Treatment and Survival Rate Analysis
2.4. Measurement of Trehalose and Glycogen Contents under Cold Stress
2.5. Soluble Trehalase and Membrane Bound Trehalase Activity Assay
2.6. RT-qPCR Analysis
2.7. Statistical Analysis
3. Results
3.1. Distribution of Yellow and Black H. axyridis in the Overwinter Population
3.2. Changes in Trehalose and Glycogen Contents under Cold Stress
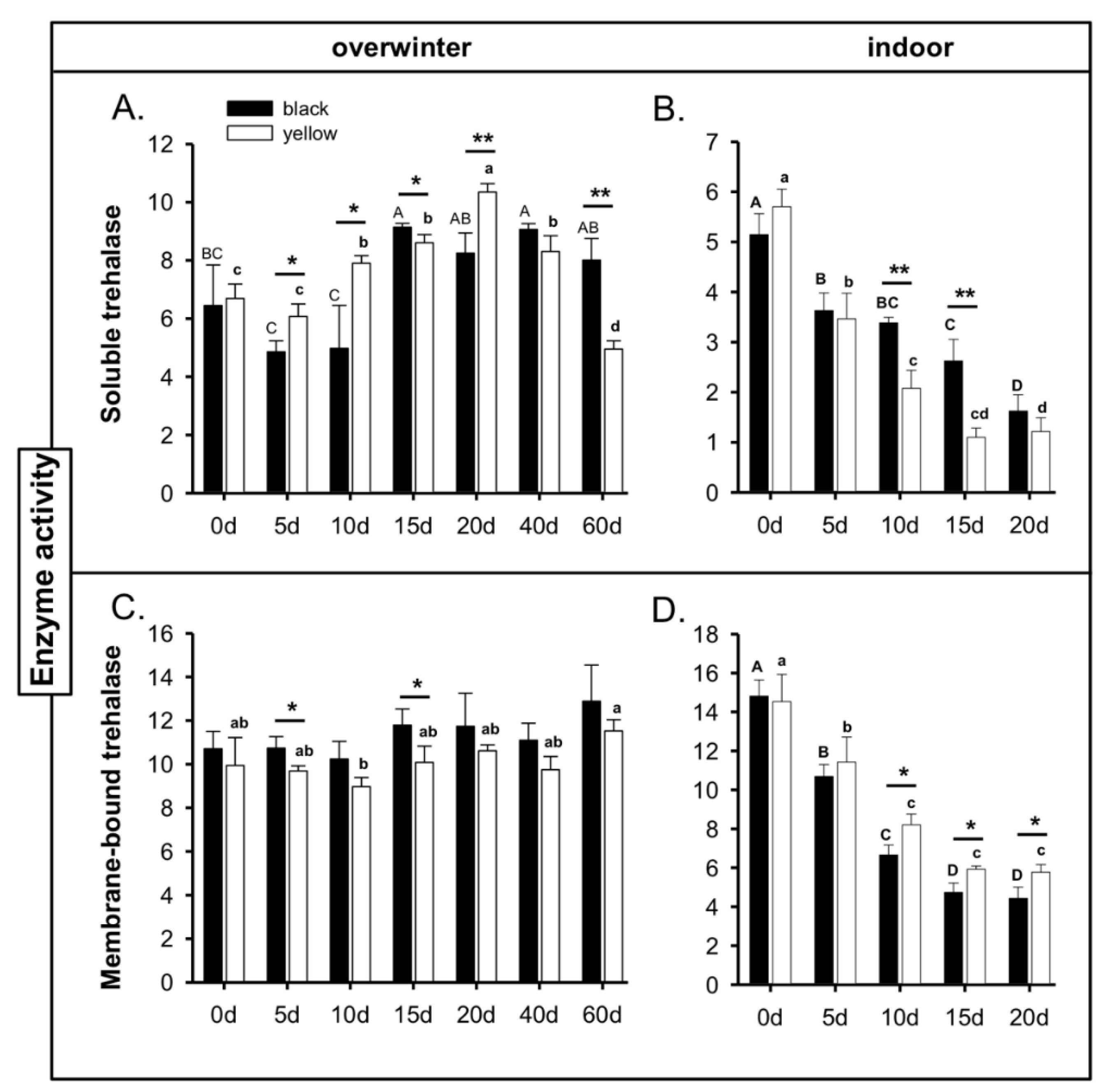
3.3. Changes in Trehalase Activity during Cold-Stress Storage
3.4. Expression of TRE and TPS Genes in the Overwinter Population during Cold Storage
3.5. Expression of TRE and TPS Genes in the Experimental Population during Cold Storage
3.6. GP and GS Gene Expression during Cold Storage
3.7. Survival Rates of H. axyridis during Cold Storage
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Riddick, E.W. Spotlight on the positive effects of the ladybird Harmonia axyridis on agriculture. BioControl 2017, 62, 319–330. [Google Scholar] [CrossRef]
- Di, N.; Zhang, K.; Xu, Q.; Zhang, F.; Harwood, J.D.; Wang, S.; Desneux, N. Predatory ability of Harmonia axyridis (Coleoptera: Coccinellidae) and Orius sauteri (Hemiptera: Anthocoridae) for suppression of fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2021, 12, 1063. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.P.; Ouyang, F.; Su, J.W.; Zhang, X.R.; Liu, C.L.; Xiao, Y.L.; Zhang, J.P.; Ge, F. Attraction of adult Harmonia axyridis to volatiles of the insectary plant Cnidium monnieri. BioControl 2020, 143, 104189. [Google Scholar]
- Thomine, E.; Rusch, A.; Supplisson, C.; Monticelli, L.S.; Amiens-Desneux, E.; Lavoir, A.V.; Desneux, N. Highly diversified crop systems can promote the dispersal and foraging activity of the generalist predator Harmonia axyridis. Entomol. Gen. 2020, 40, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Tan, X.L.; Michaud, J.P.; Shi, Z.K.; Zhang, F. Sexual selection drives the evolution of limb regeneration in Harmonia axyridis (Coleoptera: Coccinellidae). Bull. Entomol. Res. 2015, 105, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Roy, H.E.; Brown, P.M.J.; Adriaens, T.; Berkvens, N.; Borges, I.; Clusella-Trullas, S.; Comont, R.E.; Clercq, P.D.; Eschen, R.; Estoup, A.; et al. The harlequin ladybird, Harmonia axyridis: Global perspectives on invasion history and ecology. Biol. Invasions 2016, 18, 997–1044. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikov, A.N.; Belyakova, N.A.; Ovchinnikova, A.A.; Reznik, S.Y. Factors determining larval cannibalistic behavior in invasive and native populations of the multicolored Asian ladybird, Harmonia axyridis. Entomol. Gen. 2019, 38, 243–254. [Google Scholar] [CrossRef]
- Mirande, L.; Desneux, N.; Haramboure, M.; Schneider, M.I. Intraguild predation between an exotic and a native coccinellid in Argentina: The role of prey density. J. Pest Sci. 2015, 88, 155–162. [Google Scholar] [CrossRef]
- Wang, S.S.; Chen, X.; Li, Y.; Pan, B.Y.; Wang, S.G.; Dai, H.J.; Wang, S.; Tang, B. Effects of changing temperature on the physiological and biochemical properties of Harmonia axyridis larvae. Entomol. Gen. 2020, 40, 229–241. [Google Scholar] [CrossRef]
- Wang, Y.S.; Yao, F.L.; Soares, M.A.; Basiri, S.E.; Amiens-Desneux, E.; Campos, M.R.; Lavoir, A.V.; Desneux, N. Effects of four non-crop plants on life history traits of the lady beetle Harmonia axyridis. Entomol. Gen. 2020, 40, 243–252. [Google Scholar] [CrossRef]
- Ando, T.; Niimi, T. Development and evolution of color patterns in ladybird beetles: A case study in Harmonia axyridis. Dev. Growth Diff. 2019, 61, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xiao, D.; Du, X.Y.; Zhang, F.; Zang, L.S.; Wang, S. Impact of polymorphism and abiotic conditions on prey consumption by Harmonia axyridis. Entomol. Gen. 2019, 39, 251–258. [Google Scholar] [CrossRef]
- Gautier, M.; Yamaguchi, J.; Foucaud, J.; Loiseau, A.; Ausset, A.; Facon, B.; Gschloessl, B.; Lagnel, J.; Loire, E.; Parrinello, H.; et al. The genomic basis of color pattern polymorphism in the Harlequin ladybird. Curr. Biol. 2018, 28, 3296–3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.M.; Yang, X.B.; Zang, L.S.; Wang, Z.; Ruan, C.C.; Liu, X.J. Effect of geographic variation on biology and cold tolerance of Harmonia axyridis in China. Entomol. Gen. 2017, 36, 239–250. [Google Scholar] [CrossRef]
- Ando, T.; Matsuda, T.; Goto, K.; Hara, K.; Ito, A.; Hirata, J.; Yatomi, J.; Kajitani, R.; Okuno, M.; Yamaguchi, K.; et al. Repeated inversions within a pannier intron drive diversification of intraspecific colour patterns of ladybird beetles. Nat. Commun. 2018, 9, 3843. [Google Scholar] [CrossRef] [Green Version]
- Heimpel, G.E.; Lundgren, J.G. Sexual ration of commercially reared biological control agent. BioControl. 2000, 19, 77–93. [Google Scholar]
- Seo, M.J.; Youn, Y.N. The asian lady-bird, Harmonia axyridis, as biological control agents. I. Predacious behavior and feeding ablity. Korean J. Appl. Entomol. 2000, 39, 59–71. [Google Scholar]
- Knapp, M.; Vernon, P.; Renault, D. Studies on chill coma recovery in the ladybird, Harmonia axyridis: Ontogenetic profile, effect of repeated cold exposures, and capacity to predict winter survival. J. Therm. Biol. 2018, 74, 275–280. [Google Scholar] [CrossRef]
- Shukla, E.; Thorat, L.J.; Nath, B.B.; Gaikwad, S.M. Insect trehalase: Physiological significance and potential applications. Glycobiology 2015, 25, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.N.; Ji, S.X.; Liu, W.X.; Guo, J.Y.; Lu, Z.C.; Wan, F.H. Molecular characteristics of three cold resistance genes and their roles in temperature stress response in two Bemisia tabaci cryptic species. Entomol. Gen. 2021, 41, 317–328. [Google Scholar]
- Shi, Z.K.; Liu, X.J.; Xu, Q.Y.; Qin, Z.; Wang, S.; Zhang, F.; Wang, S.G.; Tang, B. Two novels soluble trehalase genes cloned from Harmonia axyridis and regulation of the enzyme in a rapid changing temperature. Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2016, 198, 10–18. [Google Scholar] [CrossRef]
- Di, N.; Wang, S.; Ridsdill-Smith, J.; Chen, Y.F.; Harwood, J.D.; Zhang, K.; Liu, T.X. Nitrogen and plant growth regulator affect plant detoxification metabolism and tritrophic interactions among Triticum aestivum, Sitobion avenae and Aphelinus asychis. Entomol. Gen. 2021, 41, 369–384. [Google Scholar]
- Matsuda, H.; Yamada, T.; Yoshida, M.; Nishimura, T. Flies without trehalose. J. Biol. Chem. 2015, 290, 1244–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanmohamadi, F.; Khajehali, J.; Izadi, H. Diapause and cold hardiness of the almond wasp, Eurytoma amygdali (Hymenoptera: Eurytomidae), two independent phenomena. J. Econ. Entomol. 2016, 109, 1646–1650. [Google Scholar] [CrossRef] [PubMed]
- Takiguchi, M.; Niimi, T.; Su, Z.H.; Yaginuma, T. Trehalase from male accessory gland of an insect, Tenebrio molitor. cDNA sequencing and developmental profile of the gene expression. Biochem. J. 1992, 288, 19–22. [Google Scholar]
- Zhao, L.N.; Yang, M.M.; Shen, Q.D.; Liu, X.J.; Shi, Z.K.; Wang, S.G.; Tang, B. Functional characterization of three trehalase genes regulating the chitin metabolism pathway in rice brown planthopper using RNA interference. Sci. Rep. 2016, 6, 27841. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Wang, S.; Wang, S.G.; Wang, H.J.; Zhang, J.Y.; Cui, S.Y. Invertebrate trehalose-6-phosphate synthase gene: Genetic architecture, biochemistry, physiological function, and potential applications. Front. Physiol. 2018, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.J.; Zhang, T.; Li, C.; Zhi, J.R. Cloning spatio-temporal expression profile and Response to temperature stress of trehalose synthase gene from Spodoptera Litura. J. Entomol. 2021, 64, 1417–1426. [Google Scholar]
- Zhang, Y.; Wang, F.; Feng, Q.; Wang, H.; Tang, T.; Huang, D.; Liu, F. Involvement of trehalose-6-phosphate synthase in innate immunity of Musca domestica. Dev. Comp. Immunol. 2019, 91, 85–92. [Google Scholar] [CrossRef]
- Knapp, M.; Řeřicha, M. Effects of the winter temperature regime on survival, body mass loss and post-winter starvation resistance in laboratory-reared and field-collected ladybirds. Sci. Rep. 2020, 10, 4970. [Google Scholar] [CrossRef] [Green Version]
- Colinet, H.; Rinehart, J.P.; Yocum, G.D.; Greenlee, K.J. Mechanisms underpinning the beneficial effects of fluctuating thermal regimes in insect cold tolerance. J. Exp. Biol. 2018, 23 Pt 14, 221. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.F. Studies on physiological response and offspring fitness of Harmonia axyridis to rapid cold acclimation. HZAU 2018, 1–72. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD201901&filename=1018192794.nh (accessed on 3 November 2022).
- Zeng, B.P.; Wang, S.S.; Li, Y.; Xiao, Z.J.; Zhou, M.; Wang, S.G.; Zhang, D.W. Effect of long-term cold storage on trehalose metabolism of pre-wintering Harmonia axyridis adults and changes in morphological diversity before and after wintering. PLoS ONE. 2020, 15, e0230435. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.; Alves-Bezerra, M.; Rosas-Oliveira, R.; Majerowicz, D.; Meyer-Fernandes, J.R.; Gondim, K.C. Gene identification and enzymatic properties of a membrane-bound trehalase from the ovary of Rhodnius prolixus. Arch. Insect Biochem. Physiol. 2012, 81, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Tatun, N.; Singtripop, T.; Tungjitwitayakul, J.; Sakurai, S. Regulation of soluble and membrane-bound trehalase activity and expression of the enzyme in the larval midgut of the bamboo borer Omphisa fuscidentalis. Insect Biochem. Mol. Biol. 2008, 38, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Xie, W.; Zhang, Z.; Wang, S.L.; Wu, Q.J.; Liu, Y.; Zhou, X.M.; Zhou, X.G.; Zhang, Y.J. Exploring valid reference genes for quantitative real-time PCR analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 2013, 9, 792–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osanai-Futahashi, M.; Ohde, T.; Hirata, J.; Uchino, K.; Futahashi, R.; Tamura, T.; Niimi, T.; Sezutsu, H. A visible dominant marker for insect transgenesis. Nat. Commun. 2012, 3, 1295. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Shi, Z.K.; Shen, Q.D.; Xu, C.D.; Wang, B.; Meng, Z.J.; Wang, S.G.; Tang, B.; Wang, S. Molecular cloning and induced expression of six small heat shock proteins mediating cold-hardiness in Harmonia axyridis (Coleoptera: Coccinellidae). Front. Physiol 2017, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Xiao, D.; Li, X.L.; Xu, Y.Y.; Wang, S. Cold tolerance of different adult elytral color morphs in Harmonia axyridis (Coleoptera: Coccinellidae). Chin. J. Entomol. 2015, 52, 428–433. [Google Scholar]
- Wang, S.; Michaud, J.P.; Zhang, R.Z.; Zhang, F.; Liu, S. Seasonal cycles of assortative mating and reproductive behavior in polymorphic populations of Harmonia axyridis in China. Ecol. Entomol. 2009, 34, 483–494. [Google Scholar]
- Cuesta, E.; Lobo, J.M. Visible and near-infrared radiation may be transmitted or absorbed differently by beetle elytra according to habitat preference. PeerJ 2019, 7, e8104. [Google Scholar] [CrossRef]
- Purse, B.V.; Comont, R.; Butler, A.; Brown, P.M.J.; Kessel, C.; Roy, H.E. Landscape and climate determine patterns of spread for all colour morphs of the alien ladybird Harmonia axyridis. J. Biogeogr. 2015, 42, 575–588. [Google Scholar] [CrossRef]
- Geng, Z.C.; Tan, Z. Several genetic problems of Harmonia axyridis. J. Nat. 1980, 3, 512–518. [Google Scholar]
- Knapp, M.; Nedvěd, O. Gender and timing during ontogeny matter: Effects of a temporary high temperature on survival, body size and colouration in Harmonia axyridis. PloS One. 2013, 8, e74984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.C. Mosaic dominance in the inheritance of color patterns in the lady-bird beetle, Harmonia axyridis. Genetics. 1946, 31, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Su, Z.H.; Pang, H. Genetic diversity of populations of Harmonia axyridis (Coleoptera: Coccinellidae) from USA andChina revealed by ISSR-PCR. J. Environ. Entomol. 2009, 31, 311–319. [Google Scholar]
- Zhao, J.; Li, X.L.; Xu, Y.Y.; Wang, S. Effects of overwintering aggregation behavior on energy metabolism in Harmonia axyridis (Coleoptera: Coccinellidae). J. Econ. Entomol. 2014, 36, 879–883. [Google Scholar]
- Wang, S.; Michaud, J.P.; Tan, X.L.; Zhang, F.; Guo, X.J. The aggregation behavior of Harmonia axyridis in its native range in Northeast China. BioControl 2011, 56, 193–206. [Google Scholar] [CrossRef]
- Ruan, C.C.; Du, W.M.; Wang, X.M.; Zhang, J.J.; Zang, L.S. Effect of long-term cold storage on the fitness of pre-wintering Harmonia axyridis, (Pallas). BioControl 2012, 57, 95–102. [Google Scholar] [CrossRef]
- Labrie, G.; Coderre, D.; Lucas, É. Overwintering strategy of multicolored Asian lady beetle (Coleoptera: Coccinellidae): Cold-free space as a factor of invasive success. Ann. Entomol. Soc. Am. 2008, 101, 860–866. [Google Scholar] [CrossRef]
- Řeřicha, M.; Dobeš, P.; Knapp, M. Changes in haemolymph parameters and insect ability to respond to immune challenge during overwintering. Ecol. Evol. 2021, 11, 4267–4275. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Chang, N.T. Effects of temperature on the development and population growth of the melon thrips, Thrips palmi, on eggplant, Solanum melongena. J. Insect Sci. 2014, 14, 78. [Google Scholar] [CrossRef]
- Broufas, G.D.; Koveos, D.S. Rapid cold hardening in the predatory mite Euseius (Amblyseius) finlaandicus (Acari: Phytoseiidae). J. Insect Physiol. 2001, 47, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Heydari, M.; Izadi, H. Effects of seasonal acclimation on cold tolerance and biochemical status of the carob moth, Ectomyelois ceratoniae Zeller, last instar larvae. Bull. Entomol. Res. 2014, 104, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V.; Miklas, B.; Doležal, P.; Rozsypal, J.; Zahradníčková, H. Physiology of cold tolerance in the bark beetle, Pityogenes chalcogra phus and its overwintering in spruce stands. J. Insect Physiol. 2014, 63, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Du, W.M.; Zhang, J.J.; Sun, G.Z.; Zang, L.S.; Ruan, C.C. Effects of cold storage on physiological and biochemical indexes of overwintering Harmonia axyridis (Pall). J. Jilin Univ. Med. Ed. 2014, 36, 536–539. [Google Scholar]
- Chen, J.; Tang, B.; Chen, H.X.; Yao, Q.; Huang, X.F.; Chen, J.; Zhang, D.W.; Zhang, W.Q. Different functions of the insect soluble and membrane-bound trehalase genes in chitin biosynthesis revealed by RNA Interference. PLoS ONE. 2010, 5, e10133. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Chen, J.; Yao, Q.; Pan, Z.Q.; Xu, W.H.; Wang, S.G.; Zhang, W.Q. Characterization of a trehalose-6-phosphate synthase gene from Spodoptera exigua and its function identification through RNA interference. J. Insect Physiol. 2010, 56, 813–821. [Google Scholar] [CrossRef]
- Tang, B.; Xiao, Z.J.; Zeng, B.P.; Qiu, L.Y.; Pan, B.Y.; Li, K.; Zhang, D.W. Characteristics analysis of TRE gene and RNAi suppression expression effect of Tribolium castaneum. J. Econ. Entomol. 2019, 41, 1311–1320. [Google Scholar]
- Łopieńska-Biernat, E.; Żółtowska, K.; Zaobidna, E.A.; Dmitryjuk, N.; Bak, B. Developmental changes in gene expression and enzyme activities of anabolic and catabolic enzymes for storage carbohydrates in the honeybee, Apis mellifera. Insectes Soc. 2018, 65, 571–580. [Google Scholar] [CrossRef] [Green Version]
- Mori, H.; Lee, J.H.; Okuyama, M.; Nishimoto, M.; Ohguchi, M.; Kim, D.; Kimura, A.; Chiba, S. Catalytic reaction mechanism based on alpha-secondary deuterium isotope effects in hydrolysis of trehalose by European honeybee trehalase. Biosci. Biotechnol. Biochem. 2009, 73, 2466–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamei, Y.; Hasegawa, Y.; Niimi, T.; Yamashita, O.; Yaginuma, T. Trehalase-2 protein contributes to trehalase activity enhanced by diapause hormone in developing ovaries of the silkworm, Bombyx mori. J. Insect Physiol. 2011, 57, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lu, D.H.; Pu, J.; Wu, M.; Han, Z.J. Cloning and RNA interference effects of trehalase genes in Laodelphax striatellus (Homoptera: Delphacidae). Acta. Entomol. Sin. 2012, 55, 911–920. [Google Scholar]
- Tatun, N.; Singtripop, T.; Sakurai, S. Dual control of midgut trehalase activity by 20-hydroxyecdysone and an inhibitory factor in the bamboo borer Omhisa fuscidentalis Hampson. J. Insect Physiol. 2008, 54, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Mian, M.A.; Mittapalli, O.; Michel, A.P. Molecular characterization and expression analysis of soluble trehalase gene in Aphis glycines, a migratory pest of soybean. Bull. Entomol. Res. 2013, 103, 286–295. [Google Scholar] [CrossRef]
- Tang, B.; Yang, M.M.; Shen, Q.D.; Xu, Y.X.; Wang, H.J.; Wang, S.G. Suppressing the activity of trehalase with validamycin disrupts the trehalose and chitin biosynthesis pathways in the rice brown planthopper, Nilaparvata lugens. Pestic. Biochem. Physiol. 2017, 137, 81–90. [Google Scholar] [CrossRef]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colinet, H.; Renaul, D.; Hance, T.; Veron, P. The impact offuctuatina therma reimes on the survival of a cold-exposed parasitic wasp. Aphidius colemani. Physiol. Entomol. 2006, 31, 234–240. [Google Scholar] [CrossRef]
- Teets, N.M.; Kawarasaki, Y.; Lee, R.E., Jr.; Denlinger, D.L. Survival and energetic costs of repeated cold exposure in the Antarctic midge, Belgica antarctica: A comparison between frozen and supercooled larvae. J. Exp. Biol. 2011, 214 Pt 5, 806–814. [Google Scholar] [CrossRef] [Green Version]
- Kawarasaki, Y.; Teets, N.M.; Denlinger, D.L.; Lee, R.E., Jr. The protective effect of rapid cold-hardening develops more quickly in frozen versus supercooled larvae of the Antarctic midge, Belgica antarctica. J. Exp. Biol. 2013, 216 Pt 20, 3937–3945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, J.B.; Lopez-Martinez, G.; Elnitsky, M.A.; Lee, R.E., Jr.; Denlinger, D.L. Dehydration-induced cross tolerance of Belgica antarctica larvae to cold and heat is facilitated bytrehalose accumulation. Comp. Biochem. Physiol. A-Mol. Integr. Physiol. 2009, 152, 518–523. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Name | Nucleotide Sequences (5′-3′) | GenBank Number |
|---|---|---|---|
| Treh1-1 | HaTreh1-1QF | CTTCGCCAGTCAAATCGTCA | HM056038 |
| HaTreh1-1QR | CCGTTTGGGACATTCCAGATA | ||
| Treh1-2 | HaTreh1-2QF | TGACAACTTCCAACCTGGTAATG | FJ501961 |
| HaTreh1-2QR | TTCCTTCGAGACATCTGGCTTA | ||
| Treh1-3 | HaTreh1-3QF | ACAGTCCCTCAGAATCTATCGTCA | JX514372 |
| HaTreh1-3QR. | GGAGCCAAGTCTCAAGCTCATC | ||
| Treh1-4 | HaTreh1-4QF | TTACTGCCAGTTTGATGACCATT | KP318742 |
| HaTreh1-4QR | CATTTCGCTAATCAGAAGACCCT | ||
| Treh1-5 | HaTreh1-5QF | TGATGATGAGGTACGACGAGAA | KX349223 |
| HaTreh1-5QR | GTAGCAAGGACCTAACAAACTGC | ||
| Treh2-1 | HaTreh2-1QF | TTCCAGGTGGGAGATTCAGG | KX349224 |
| HaTreh2-1QR | GGGATCAATGTAGGAGGCTGTG | ||
| Treh2-2 | HaTreh2-2QF | CAATCAGGGTGCTGTAATGTCG | KX349225 |
| HaTreh2-2QR | CGTAGTTGGCTCATTCGTTTCC | ||
| TPS | HaTPS-QF | GACCCTGACGAAGCCATACC | FJ501960 |
| HaTPS-QR | AAAGTTCCATTACACGCACCA | ||
| GP | HaGP-QF | GCTGAAGCCCTCTACCAACT | NC059507 |
| HaGP-QR | CGCCGTACTCGTATCTTATGC | ||
| GS | HaGS-QF | CCCTTAGGATCGGATGTTCTC | NC059503 |
| HaGS-QR | CACCAGCCATCTCCCAGTT | ||
| Harp49 | Harp49-qF | GCG ATC GCTATGGAAAAC TC | AB552923 |
| Harp49-qR | TACGATTTTGCATCAACAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, S.; He, J.; Chao, L.; Shi, Z.; Wang, S.; Yu, W.; Huang, Z.; Wang, S.; Wang, S.; Zhang, Z. Regulatory Role of Trehalose Metabolism in Cold Stress of Harmonia axyridis Laboratory and Overwinter Populations. Agronomy 2023, 13, 148. https://doi.org/10.3390/agronomy13010148
Wan S, He J, Chao L, Shi Z, Wang S, Yu W, Huang Z, Wang S, Wang S, Zhang Z. Regulatory Role of Trehalose Metabolism in Cold Stress of Harmonia axyridis Laboratory and Overwinter Populations. Agronomy. 2023; 13(1):148. https://doi.org/10.3390/agronomy13010148
Chicago/Turabian StyleWan, Sijing, Jianyun He, Lei Chao, Zuokun Shi, Shasha Wang, Weidong Yu, Zhen Huang, Su Wang, Shigui Wang, and Zhijun Zhang. 2023. "Regulatory Role of Trehalose Metabolism in Cold Stress of Harmonia axyridis Laboratory and Overwinter Populations" Agronomy 13, no. 1: 148. https://doi.org/10.3390/agronomy13010148
APA StyleWan, S., He, J., Chao, L., Shi, Z., Wang, S., Yu, W., Huang, Z., Wang, S., Wang, S., & Zhang, Z. (2023). Regulatory Role of Trehalose Metabolism in Cold Stress of Harmonia axyridis Laboratory and Overwinter Populations. Agronomy, 13(1), 148. https://doi.org/10.3390/agronomy13010148