

Sustainable Agriculture: Rare-Actinomycetes to the Rescue


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. History of Sustainable Agricultural Practices
3. Sustainable Agriculture in the Resolution of Problems in Africa
3.1. Rare Actinomycetes as a Phytoenhancer
3.2. Rare Actinomycetes as Phytoprotectant
4. Rare Actinomycetes to the Rescue
5. Agro-Antibiotics Encoded by Biosynthetic Gene Clusters (BGCs) in Actinomycetes
5.1. Herbicidal Agents
5.2. Insecticidal/Acaricidal Agents
5.3. Anti-Phytopathogenic Agents
6. Targets of Fungicide
6.1. Cell Membrane Target
6.2. The Target of Cytoplasmic Organelles
7. Novel Fungicide Mining through Genome Sequencing
8. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheteni, P.; Umejesi, I. Sustainability of agriculture development in South Africa: Towards Vision 2030. Econ. Manag. Sustain. 2022, 7, 70–85. [Google Scholar] [CrossRef]
- Cilliers, J. The Future of Africa: Challenges and Opportunities; Springer Nature: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Mahon, N.; Crute, I.; Di Bonito, M.; Simmons, E.A.; Islam, M.M. Towards a broad-based and holistic framework of Sustainable Intensification indicators. Land Use Policy 2018, 77, 576–597. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, F.L.; Rosenstock, T.S.; Gitz, V.; Wollenberg, L. Agroforestry to Diversify Farms and Enhance Resilience. In 10 Best Bet Innovations for Adaptation in Agriculture: A Supplement to the UNFCCC NAP; CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS): Wageningen, The Netherlands, 2017; pp. 14–19. [Google Scholar]
- Talaviya, T.; Shah, D.; Patel, N.; Yagnik, H.; Shah, M. Implementation of artificial intelligence in agriculture for optimisation of irrigation and application of pesticides and herbicides. Artif. Intell. Agric. 2020, 4, 58–73. [Google Scholar] [CrossRef]
- Kayembe, C.; Nel, D. Challenges and opportunities for education in the Fourth Industrial Revolution. Afr. J. Public Aff. 2019, 11, 79–94. [Google Scholar]
- Santos, L.F.; Olivares, F.L. Plant microbiome structure and benefits for sustainable agriculture. Curr. Plant Biol. 2021, 26, 100198. [Google Scholar] [CrossRef]
- Glick, B.R.; Gamalero, E. Recent developments in the study of plant microbiomes. Microorganisms 2021, 9, 1533. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O.; Glick, B.R. Plant growth-promoting root-colonizing bacterial endophytes. Rhizosphere 2021, 20, 100433–100445. [Google Scholar] [CrossRef]
- Santoyo, G.; Guzmán-Guzmán, P.; Parra-Cota, F.I.; Santos-Villalobos, S.D.L.; Orozco-Mosqueda, M.D.C.; Glick, B.R. Plant growth stimulation by microbial consortia. Agronomy 2021, 11, 219. [Google Scholar] [CrossRef]
- Santoyo, G.; Gamalero, E.; Glick, B.R. Mycorrhizal-bacterial amelioration of plant abiotic and biotic stress. Front. Sustain. Food Syst. 2021, 5, 672881. [Google Scholar] [CrossRef]
- Akinola, S.A.; Ayangbenro, A.S.; Babalola, O.O. The diverse functional genes of maize rhizosphere microbiota assessed using shotgun metagenomics. J. Sci. Food Agric. 2021, 101, 3193–3201. [Google Scholar] [CrossRef] [PubMed]
- Babalola, O.O.; Kirby, B.M.; Le Roes-Hill, M.; Cook, A.E.; Cary, S.C.; Burton, S.G.; Cowan, D.A. Phylogenetic analysis of actinobacterial populations associated with Antarctic Dry Valley mineral soils. Environ. Microbiol. 2009, 11, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Seenivasagan, R.; Babalola, O.O. Utilization of microbial consortia as biofertilizers and biopesticides for the production of feasible agricultural product. Biology 2021, 10, 1111. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Wu, Z.; Zhang, Y.; Zhang, Z.; Fang, W.; Wang, Y.; Wan, Z.; Wang, K.; Ke, S. Herbicidal secondary metabolites from Actinomycetes: Structure diversity, modes of action, and their roles in the development of herbicides. J. Agric. Food Chem. 2019, 68, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Hwang, S.; Kim, J.; Cho, S.; Palsson, B.; Cho, B.K. Mini review: Genome mining approaches for the identification of secondary metabolite biosynthetic gene clusters in Streptomyces. Comput. Struct. Biotechnol. J. 2020, 18, 1548–1556. [Google Scholar] [CrossRef]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Rothman, J.M.; Pontzer, H.; Simpson, S.J. Macronutrient contributions of insects to the diets of hunter–gatherers: A geometric analysis. J. Hum. Evol. 2014, 71, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Vasey, D.E. An Ecological History of Agriculture 10000 Bc-Ad 10000; Purdue University Press: West Lafayette, IN, USA, 2002. [Google Scholar] [CrossRef]
- Sinclair, F.; Wezel, A.; Mbow, C.; Chomba, S.; Robiglio, V.; Harrison, R. The Contribution of Agroecological Approaches to Realizing Climate-Resilient Agriculture; GCA: Rotterdam, The Netherlands, 2019. [Google Scholar]
- Tiwari, T.P.; Brook, R.M.; Wagstaff, P.; Sinclair, F.L. Effects of light environment on maize in hillside agroforestry systems of Nepal. Food Secur. 2012, 4, 103–114. [Google Scholar] [CrossRef]
- Kong, Z.; Hart, M.; Liu, H. Paving the way from the lab to the field: Using synthetic microbial consortia to produce high-quality crops. Front. Plant Sci. 2018, 9, 1467–1472. [Google Scholar] [CrossRef] [Green Version]
- Chukwuneme, C.F.; Babalola, O.O.; Kutu, F.R.; Ojuederie, O.B. Characterization of actinomycetes isolates for plant growth promoting traits and their effects on drought tolerance in maize. J. Plant Interact. 2020, 15, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef]
- Shanthi, V. Actinomycetes: Implications and Prospects in Sustainable Agriculture. In Biofertilizers: Study and Impact; Wiley: Hoboken, NJ, USA, 2021; pp. 335–370. [Google Scholar] [CrossRef]
- Zhang, H.; Han, L.; Jiang, B.; Long, C. Identification of a phosphorus-solubilizing Tsukamurella tyrosinosolvens strain and its effect on the bacterial diversity of the rhizosphere soil of peanuts growth-promoting. World J. Microbiol. Biotechnol. 2021, 37, 109. [Google Scholar] [CrossRef]
- Alipour Kafi, S.; Karimi, E.; Akhlaghi Motlagh, M.; Amini, Z.; Mohammadi, A.; Sadeghi, A. Isolation and identification of Amycolatopsis sp. strain 1119 with potential to improve cucumber fruit yield and induce plant defense responses in commercial greenhouse. Plant Soil 2021, 468, 125–145. [Google Scholar] [CrossRef]
- Mawang, C.I.; Azman, A.S.; Fuad, A.S.M.; Ahamad, M. Actinobacteria: An eco-friendly and promising technology for the bioaugmentation of contaminants. Biotechnol. Rep. 2021, 32, e00679. [Google Scholar] [CrossRef]
- Jiang, B.; Long, C.; Xu, Y.; Han, L. Molecular mechanism of Tsukamurella tyrosinosolvens strain P9 in response to root exudates of peanut. Arch. Microbiol. 2023, 205, 48. [Google Scholar] [CrossRef]
- Shen, Q.; Dai, G.; Li, A.; Liu, Y.; Zhong, G.; Li, X.; Ren, X.; Sui, H.; Fu, J.; Jiao, N.; et al. Genome-Guided Discovery of Highly Oxygenated Aromatic Polyketides, Saccharothrixins D–M, from the Rare Marine Actinomycete Saccharothrix sp. D09. J. Nat. Prod. 2021, 84, 2875–2884. [Google Scholar] [CrossRef]
- Liu, K.; Hu, X.R.; Zhao, L.X.; Wang, Y.; Deng, Z.; Tao, M. Enhancing ristomycin a production by overexpression of ParB-like StrR family regulators controlling the biosynthesis genes. Appl. Environ. Microbiol. 2021, 87, e01066-21. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, N.; He, J.; Li, Y.; Gao, X.; Huang, L.; Yan, X. Expression and characterization of a novel chitinase with antifungal activity from a rare actinomycete, Saccharothrix yanglingensis Hhs. 015. Protein Expr. Purif. 2018, 143, 45–51. [Google Scholar] [CrossRef]
- Zhao, T.; Gao, Y.; Yu, T.; Zhang, Y.; Zhang, Z.; Zhang, L.; Zhang, L. Biodegradation of phenol by a highly tolerant strain Rhodococcus ruber C1: Biochemical characterization and comparative genome analysis. Ecotoxicol. Environ. Saf. 2021, 208, 111709. [Google Scholar] [CrossRef]
- Styczynski, M.; Rogowska, A.; Gieczewska, K.; Garstka, M.; Szakiel, A.; Dziewit, L. Genome-based insights into the production of carotenoids by Antarctic bacteria, Planococcus sp. ANT_H30 and Rhodococcus sp. ANT_H53B. Molecules 2020, 25, 4357. [Google Scholar] [CrossRef]
- Su, Y.S.; Cheng, M.J.; Wu, M.D.; Chai, C.Y.; Kwan, A.L.; Su, S.H.; Kuo, Y.H. Chemical Constituents from a Mangrove-Derived Actinobacteria Isoptericola chiayiensis BCRC 16888 and Evaluation of Their Anti-NO Activity. Chem. Biodivers. 2021, 18, e2100211. [Google Scholar] [CrossRef]
- Nouioui, I.; Ha, S.M.; Baek, I.; Chun, J.; Goodfellow, M. Genome insights into the pharmaceutical and plant growth promoting features of the novel species Nocardia alni sp. nov. BMC Genom. 2022, 23, 70. [Google Scholar] [CrossRef]
- Saygin, H.; Ay, H.; Guven, K.; Inan-Bektas, K.; Cetin, D.; Sahin, N. Saccharopolyspora karakumensis sp. nov., Saccharopolyspora elongata sp. nov., Saccharopolyspora aridisoli sp. nov., Saccharopolyspora terrae sp. nov. and their biotechnological potential revealed by genome analysis. Syst. Appl. Microbiol. 2021, 44, 126270. [Google Scholar] [CrossRef]
- Abraham, M.; Pingali, P. Transforming Smallholder Agriculture to Achieve the SDGs. In The Role of Smallholder Farms in Food and Nutrition Security; Springer: Berlin/Heidelberg, Germany, 2020; pp. 173–209. [Google Scholar] [CrossRef]
- Höhne, N.; Fekete, H.; den Elzen, M.G.; Hof, A.F.; Kuramochi, T. Assessing the ambition of post-2020 climate targets: A comprehensive framework. Clim. Policy 2018, 18, 425–441. [Google Scholar] [CrossRef] [Green Version]
- Kom, Z.; Nethengwe, N.S.; Mpandeli, N.S.; Chikoore, H. Determinants of small-scale farmers’ choice and adaptive strategies in response to climatic shocks in Vhembe District, South Africa. GeoJournal 2020, 87, 677–700. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. Reclamation of arid and semi-arid soils: The role of plant growth-promoting archaea and bacteria. Curr. Plant Biol. 2021, 25, 100173–100185. [Google Scholar] [CrossRef]
- Alori, E.T.; Babalola, O.O. Microbial inoculants for improving crop quality and human health in Africa. Front. Microbiol. 2018, 9, 2213–2231. [Google Scholar] [CrossRef] [Green Version]
- Fadiji, A.E.; Santoyo, G.; Yadav, A.N.; Babalola, O.O. Efforts towards overcoming drought stress in crops: Revisiting the mechanisms employed by plant growth-promoting bacteria. Front. Microbiol. 2022, 13, 962427. [Google Scholar] [CrossRef]
- del Carmen Orozco-Mosqueda, M.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Alawiye, T.T.; Babalola, O.O. Bacterial diversity and community structure in typical plant rhizosphere. Diversity 2019, 11, 179. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, Z.; Glick, B.R. Secondary metabolites produced by plant growth-promoting bacterial endophytes. Microorganisms 2022, 10, 2008. [Google Scholar] [CrossRef]
- Subbanna, A.R.N.S.; Stanley, J.; Rajasekhara, H.; Mishra, K.K.; Pattanayak, A.; Bhowmick, R. Perspectives of Microbial Metabolites as Pesticides in Agricultural Pest Management. In Co-Evolution of Secondary Metabolites; Springer: Cham, Switzerland; Midtown Manhattan, NY, USA, 2020; pp. 925–952. [Google Scholar] [CrossRef]
- Kucho, K.I.; Hashimoto, M.; Normand, P.; Fournier, P.; Pujic, P. Structure of surface polysaccharides affects symbiotic nitrogen fixation in Frankia alni ACN14a. J. For. Res. 2022, 1–7. [Google Scholar] [CrossRef]
- Bouizgarne, B. Phosphate-Solubilizing Actinomycetes as Biofertilizers and Biopesticides: Bioformulations for Sustainable Agriculture. In Microbial BioTechnology for Sustainable Agriculture Volume 1; Springer: Gateway East, Singapore, 2022; pp. 407–428. [Google Scholar] [CrossRef]
- Adenan, N.H.; Ting, A.S.Y. Actinobacteria from Soils and their Applications in Environmental Bioremediation. In Microbial Biotechnology: Role in Ecological Sustainability and Research; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2022; pp. 313–333. [Google Scholar] [CrossRef]
- Azadi, D.; Shojaei, H. Biodegradation of polycyclic aromatic hydrocarbons, phenol and sodium sulfate by Nocardia species isolated and characterized from Iranian ecosystems. Sci. Rep. 2020, 10, 21860. [Google Scholar] [CrossRef]
- Das, P.; Singh, S.K.; Singh, P.; Zeyad, M.T.; Aamir, M.; Upadhyay, R.S. Actinomycetes as biostimulants and their application in agricultural practices. In Microbiome Stimulants for Crops; Elsevier: Amsterdam, The Netherlands, 2021; pp. 267–282. [Google Scholar] [CrossRef]
- Momin, M.D.; Singh, N.S.; Kumar, A.; Tripathi, S.K. Structural and functional characterization of rhizosphere actinomycetes of major crop plants under shifting cultivation practice in Northeast India. Vegetos 2021, 34, 638–646. [Google Scholar] [CrossRef]
- Wang, K.; Ke, S.; Fang, W.; Wu, Z.; Zhang, Y. Novel Agroactive Secondary Metabolites from Actinomycetes in the Past Two Decades with Focus on Screening Strategies and Discovery. In Natural Products from Actinomycetes; Springer: Gateway East, Singapore, 2022; pp. 199–221. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The chemistry of plant–microbe interactions in the rhizosphere and the potential for metabolomics to reveal signaling related to defense priming and induced systemic resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Widada, J.; Damayanti, E.; Alhakim, M.R.; Yuwono, T.; Mustofa, M. Two strains of airborne Nocardiopsis alba producing different volatile organic compounds (VOCs) as biofungicide for Ganoderma boninense. FEMS Microbiol. Lett. 2021, 368, 138. [Google Scholar] [CrossRef]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Andreotti, C. Use of biostimulants for organic apple production: Effects on tree growth, yield, and fruit quality at harvest and during storage. Front. Plant Sci. 2018, 9, 1342–1359. [Google Scholar] [CrossRef]
- Li, S.; Yang, B.; Tan, G.Y.; Ouyang, L.M.; Qiu, S.; Wang, W.; Xiang, W.; Zhang, L. Polyketide pesticides from actinomycetes. Curr. Opin. Biotechnol. 2021, 69, 299–307. [Google Scholar] [CrossRef]
- Myronovskyi, M.; Rosenkränzer, B.; Stierhof, M.; Petzke, L.; Seiser, T.; Luzhetskyy, A. Identification and heterologous expression of the albucidin gene cluster from the marine strain Streptomyces albus subsp. chlorinus NRRL B-24108. Microorganisms 2020, 8, 237. [Google Scholar] [CrossRef] [Green Version]
- Lukoseviciute, L.; Lebedeva, J.; Kuisiene, N. Diversity of polyketide synthases and nonribosomal peptide synthetases revealed through metagenomic analysis of a deep oligotrophic cave. Microb. Ecol. 2021, 81, 110–121. [Google Scholar] [CrossRef]
- Bozhüyük, K.A.; Micklefield, J.; Wilkinson, B. Engineering enzymatic assembly lines to produce new antibiotics. Curr. Opin. Microbiol. 2019, 51, 88–96. [Google Scholar] [CrossRef]
- Dong, X.; Lv, L.; Wang, W.; Liu, Y.; Yin, C.; Xu, Q.; Yan, H.; Fu, J.; Liu, X. Differences in distribution of potassium-solubilizing bacteria in forest and plantation soils in Myanmar. Int. J. Environ. Res. Public Health 2019, 16, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeniji, A.A.; Babalola, O.O. Evaluation of Pseudomonas fulva PS9. 1 and Bacillus velezensis NWUMFkBS10. 5 as Candidate Plant Growth Promoters during Maize-Fusarium Interaction. Plants 2022, 11, 324. [Google Scholar] [CrossRef] [PubMed]
- El-Gawahergy, H.; Amin, D.H.; Elsayed, A.F. Mining for NRPS and PKS Genes in Actinobacteria Using Whole-Genome Sequencing and Bioinformatic Tools. In Natural Products from Actinomycetes: Diversity, Ecology and Drug Discovery; Springer: Gateway East, Singapore, 2022; pp. 393–410. [Google Scholar] [CrossRef]
- Harir, M.; Bendif, H.; Bellahcene, M.; Fortas, Z.; Pogni, R. Streptomyces Secondary Metabolites. In Basic Biology and Applications of Actinobacteria; IntechOpen: London, UK, 2018; pp. 99–122. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Cao, A.; Yan, D.; Ouyang, C.; Wang, Q.; Li, Y. Overview of mechanisms and uses of biopesticides. Int. J. Pest Manag. 2021, 67, 65–72. [Google Scholar] [CrossRef]
- Hu, J.; Xia, Z.; Shuai, L.; Chen, J.; Zhu, Z.; Cao, L.; Xie, J.; Dai, Z.; Hu, Y.; Huang, W.; et al. Effect of pII key nitrogen regulatory gene on strain growth and butenyl-spinosyn biosynthesis in Saccharopolyspora pogona. Appl. Microbiol. Biotechnol. 2022, 106, 3081–3091. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Xu, X.; Han, L.; Zhao, L.; Chen, X.; Miao, C.; Hu, L.; Huang, X.; Chen, Y.; Li, Y. Echinosporin antibiotics isolated from Amycolatopsis strain and their antifungal activity against root-rot pathogens of the Panax notoginseng. Folia Microbiol. 2019, 64, 171–175. [Google Scholar] [CrossRef]
- Fei, C.; She, R.; Li, G.; Zhang, L.; Fan, W.; Xia, S.; Xue, F. Safety and clinical efficacy of tenvermectin, a novel antiparasitic 16-membered macrocyclic lactone antibiotics. Eur. J. Pharm. Sci. 2018, 117, 154–160. [Google Scholar] [CrossRef]
- Chen, X.; Hu, L.F.; Huang, X.S.; Zhao, L.X.; Miao, C.P.; Chen, Y.W.; Xu, L.H.; Han, L.; Li, Y.Q. Isolation and characterization of new phenazine metabolites with antifungal activity against root-rot pathogens of Panax notoginseng from Streptomyces. J. Agric. Food Chem. 2019, 67, 11403–11407. [Google Scholar] [CrossRef]
- Lin, S.H.; Luo, P.; Yuan, E.; Zhu, X.; Zhang, B.; Wu, X. Physiological and proteomic analysis of Penicillium digitatum in response to X33 antifungal extract treatment. Front. Microbiol. 2020, 11, 584331. [Google Scholar] [CrossRef]
- Zhang, D.; Lu, Y.; Chen, H.; Wu, C.; Zhang, H.; Chen, L.; Chen, X. Antifungal peptides produced by actinomycetes and their biological activities against plant diseases. J. Antibiot. 2020, 73, 265–282. [Google Scholar] [CrossRef]
- Zhang, S.W.; Wu, Z.M.; Yang, Y.; Li, K.T. Antifungal action of Antifungalmycin N2 against Rhizoctonia solani by disrupting cell membrane and inhibiting succinate dehydrogenase. Curr. Microbiol. 2020, 77, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Moreno, Z.R.; Vinchira-Villarraga, D.M.; Vergara-Morales, D.I.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Degrassi, G.; Venturi, V.; Moreno-Sarmiento, N. Plant-growth promotion and biocontrol properties of three Streptomyces spp. isolates to control bacterial rice pathogens. Front. Microbiol. 2019, 10, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covington, B.C.; Xu, F.; Seyedsayamdost, M.R. A natural product chemist’s guide to unlocking silent biosynthetic gene clusters. Annu. Rev. Biochem. 2021, 90, 763. [Google Scholar] [CrossRef] [PubMed]
- Majer, H.M.; Ehrlich, R.L.; Ahmed, A.; Earl, J.P.; Ehrlich, G.D.; Beld, J. Whole genome sequencing of Streptomyces actuosus ISP-5337, Streptomyces sioyaensis B-5408, and Actinospica acidiphila B-2296 reveals secondary metabolomes with antibiotic potential. Biotechnol. Rep. 2021, 29, e00596. [Google Scholar] [CrossRef]



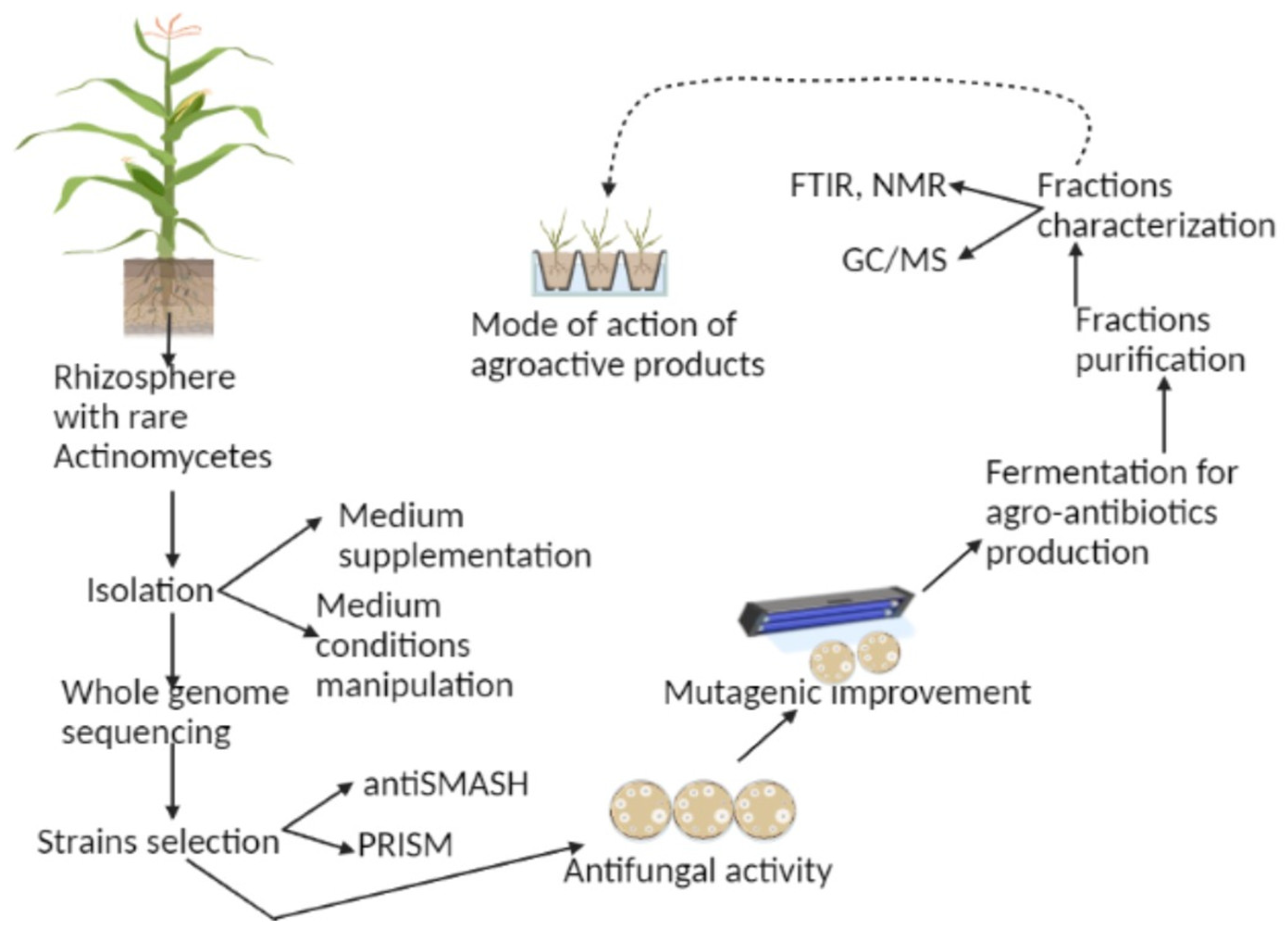{kind=link}
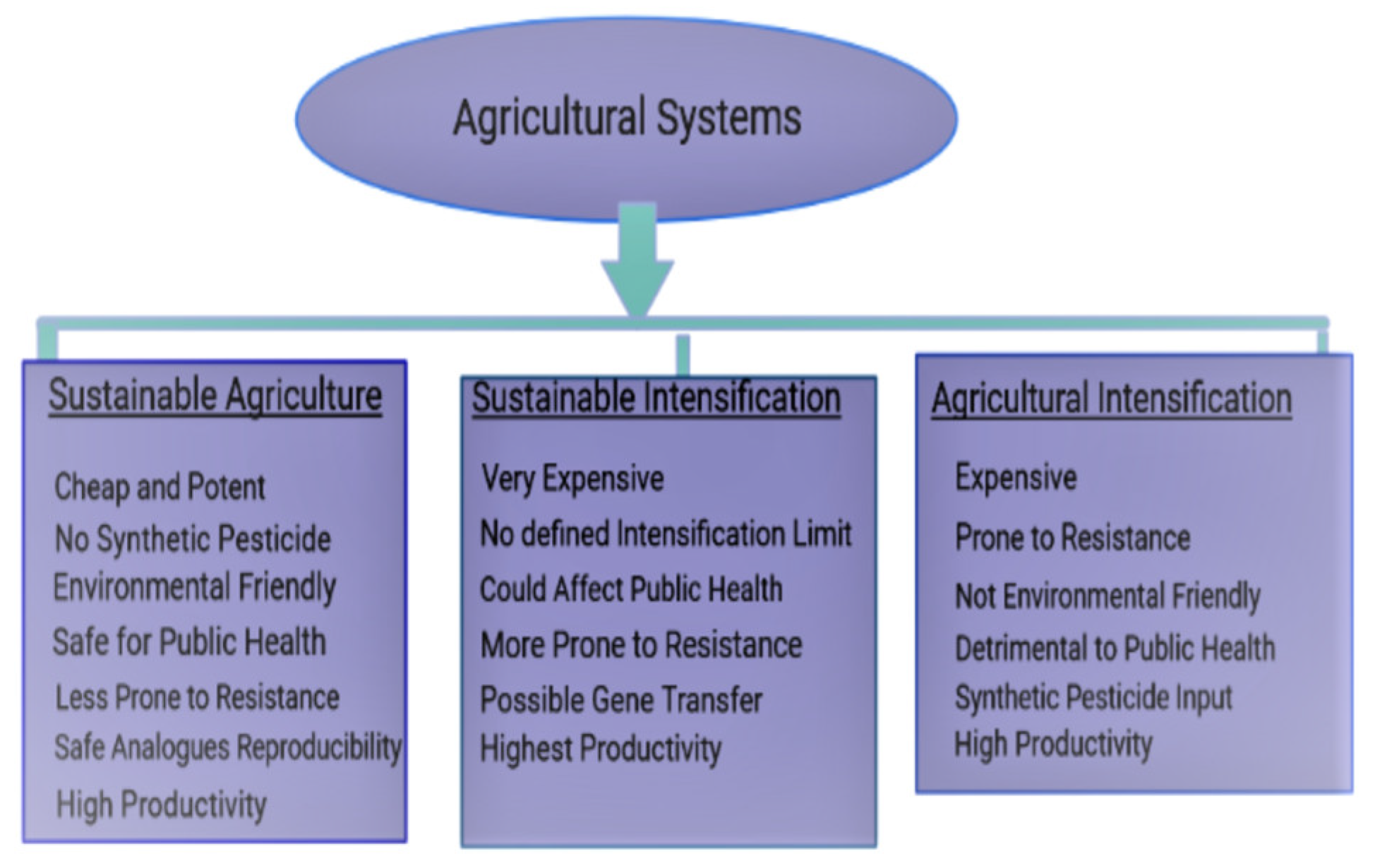{kind=link}
| Rare Actinomycetes | Core Genes | Metabolites Encoded | Functionalities | Roles in Sustainable Agriculture | References |
|---|---|---|---|---|---|
| Frankia sp. | nif | Nitrogenase enzyme | Nitrogen fixation | Soil fertilization | [25] |
| Tsukamurella tyrosinosolvens | pho | Phosphatase enzyme | Phosphate solubilization | Phosphate fertilization | [26] |
| Amycolatopsis sp. | PR1–1a & GLU | proteins | Systemic acquired resistance | Plant defence | [27] |
| Arthrobacter sp. SD3-25 | atzB, atzC & trzN | hydrolases | Atrazine & simazine pesticide biodegradation | Soil fertility and Bioaugmentation | [28] |
| mbtH, fagD | protein | Siderophores biosynthesis | Plant defence and Iron fertilization | [29] | |
| Tsukamurella tyrosinosolvens | febB, febD, yqjH, hpaC | Transport proteins | Biosynthesis of iron transporter | Plant defence and growth | [26] |
| atzF argG argH | Hydrolase Arginine succinate synthase Arginine succinate lyase | Urea degradation | Ammonia and amino acids biosynthesis for growth | [29] | |
| Saccharothrix sp. | SacA,B,C,E | Polyketide synthase | Saccharochelins A–E biosynthesis | Phytoprotection | [30] |
| Amycolatopsis sp. | asrR | Type III glycopeptide | Ristomycin | Phytoprotection | [31] |
| Saccharothrix yanglingensis Hhs.015 | Chi6769 | Protein | Chitinase biosynthesis | Phytoprotection | [32] |
| Rhodococcus ruber C1 | dmpP | Phenol hydroxylase | Phenol degradation | Biodegradation | [33] |
| Rhodococcus sp. ANT_H53B | crtP,M,N,Nc | Diapolycopene Oxygenase, dehydrosqualene synthase & other enzymes | C3 apocarotenoid biosynthesis | Phytoprotection and enhancement | [34] |
| Nonomuraea sp. NJM5123 | NRPS gene ecuE | Enzymes that form tridecapeptide full length | Ecumicins biosynthesis | Antuberculosis activity | [35] |
| Nocardia vaccinii NBRC15922 | AuaJ | Epoxidase LacC | Lasalocid | Antibacterial activity | [36] |
| Saccharopolyspora sp. | arsG | Arsenate reductase | Arsenic removal | Arsenic biocleansing | [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyedoh, O.P.; Yang, W.; Dhanasekaran, D.; Santoyo, G.; Glick, B.R.; Babalola, O.O. Sustainable Agriculture: Rare-Actinomycetes to the Rescue. Agronomy 2023, 13, 666. https://doi.org/10.3390/agronomy13030666
Oyedoh OP, Yang W, Dhanasekaran D, Santoyo G, Glick BR, Babalola OO. Sustainable Agriculture: Rare-Actinomycetes to the Rescue. Agronomy. 2023; 13(3):666. https://doi.org/10.3390/agronomy13030666
Chicago/Turabian StyleOyedoh, Oghoye P., Wei Yang, Dharumadurai Dhanasekaran, Gustavo Santoyo, Bernard R. Glick, and Olubukola O. Babalola. 2023. "Sustainable Agriculture: Rare-Actinomycetes to the Rescue" Agronomy 13, no. 3: 666. https://doi.org/10.3390/agronomy13030666
APA StyleOyedoh, O. P., Yang, W., Dhanasekaran, D., Santoyo, G., Glick, B. R., & Babalola, O. O. (2023). Sustainable Agriculture: Rare-Actinomycetes to the Rescue. Agronomy, 13(3), 666. https://doi.org/10.3390/agronomy13030666