
B Cell Response Induced by SARS-CoV-2 Infection Is Boosted by the BNT162b2 Vaccine in Primary Antibody Deficiencies


 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Study Design and Patients
2.2. ELISA for Specific IgG Detection
2.3. Detection of SARS-CoV-2-Specific B Cells
2.4. Detection of SARS-CoV-2-Specific T Cells
2.5. Statistical Analysis
3. Results
3.1. Patients
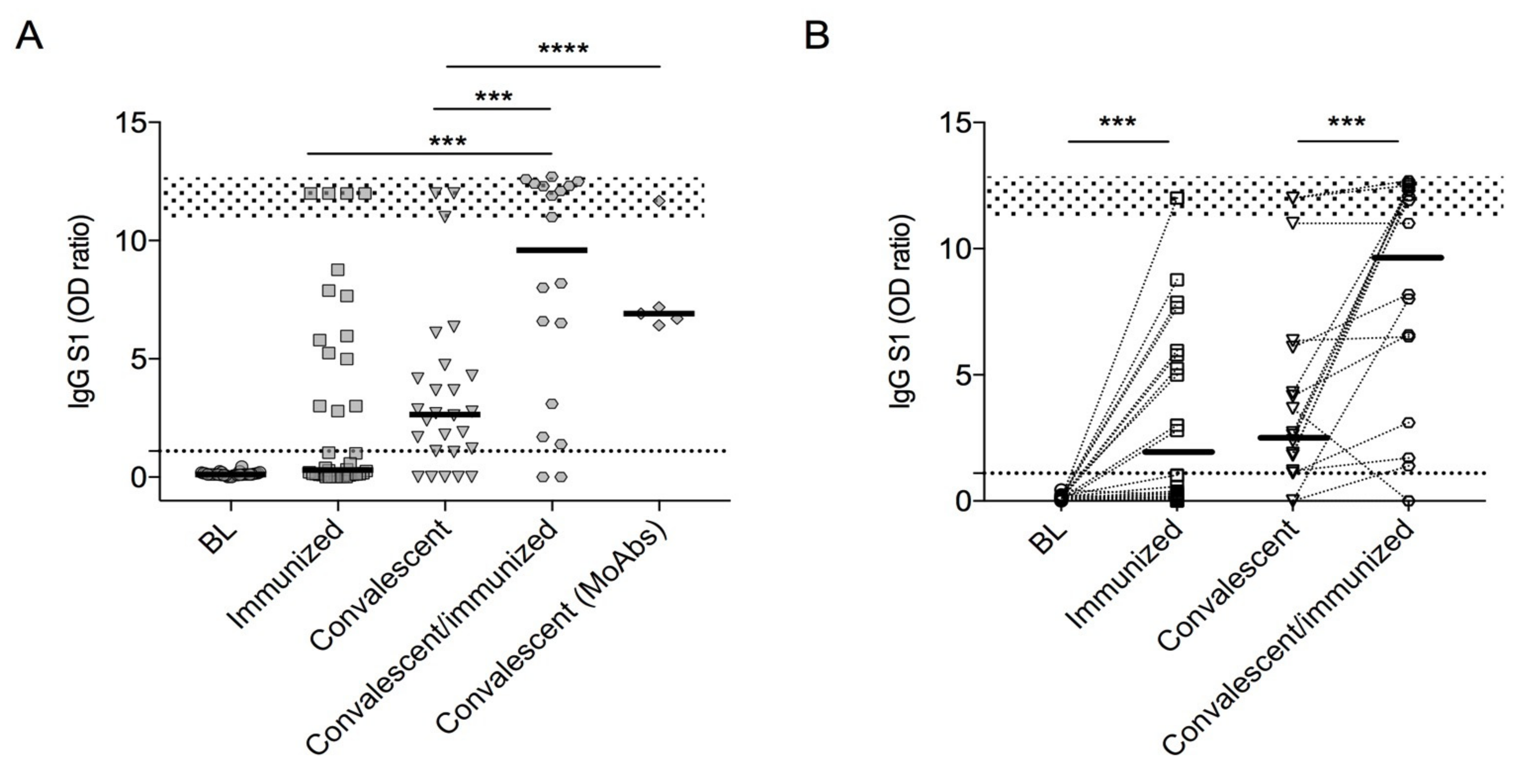
3.2. SARS-CoV-2 Antibodies
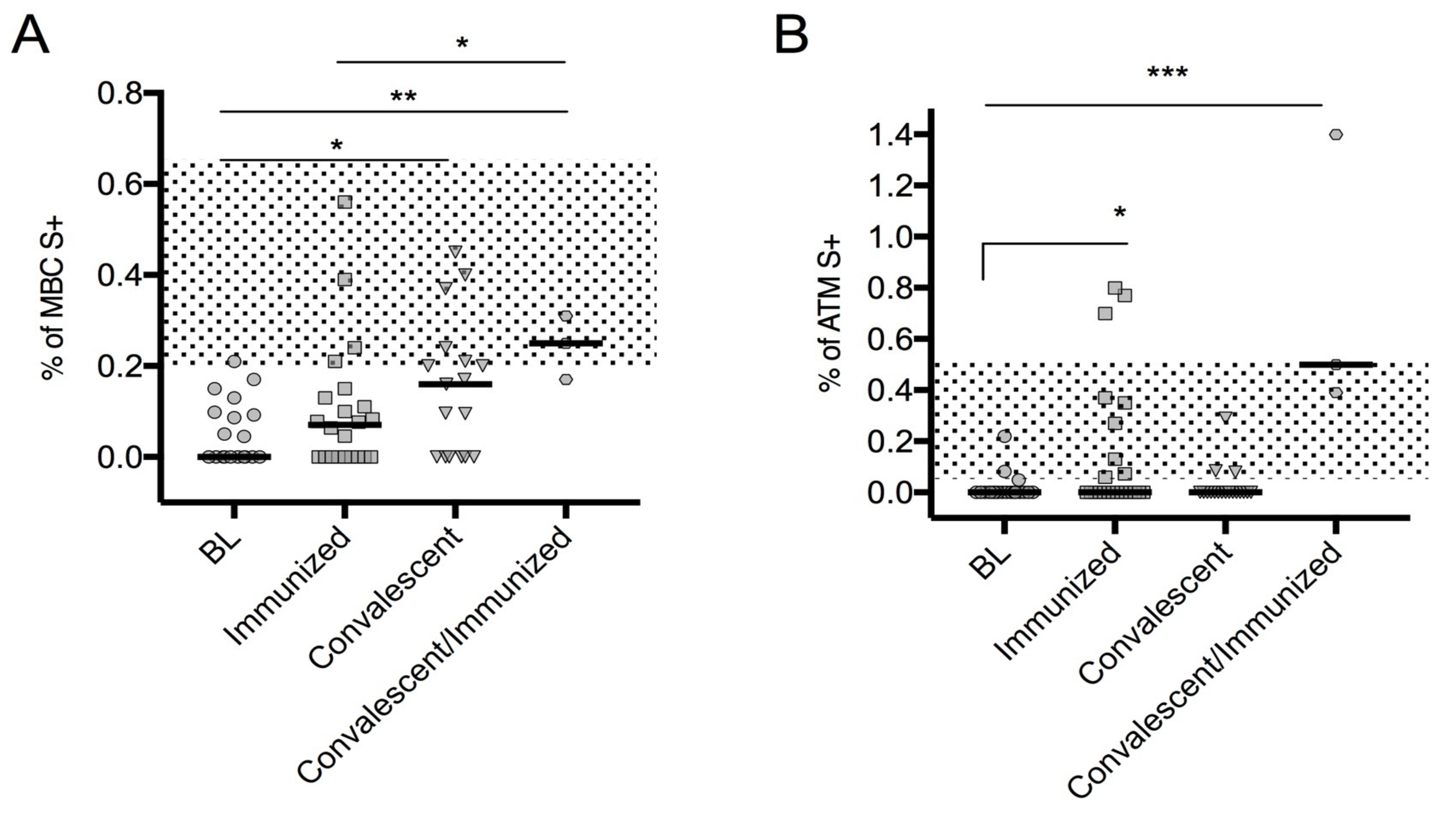
3.3. Spike-Specific SARS-CoV-2 MBCs
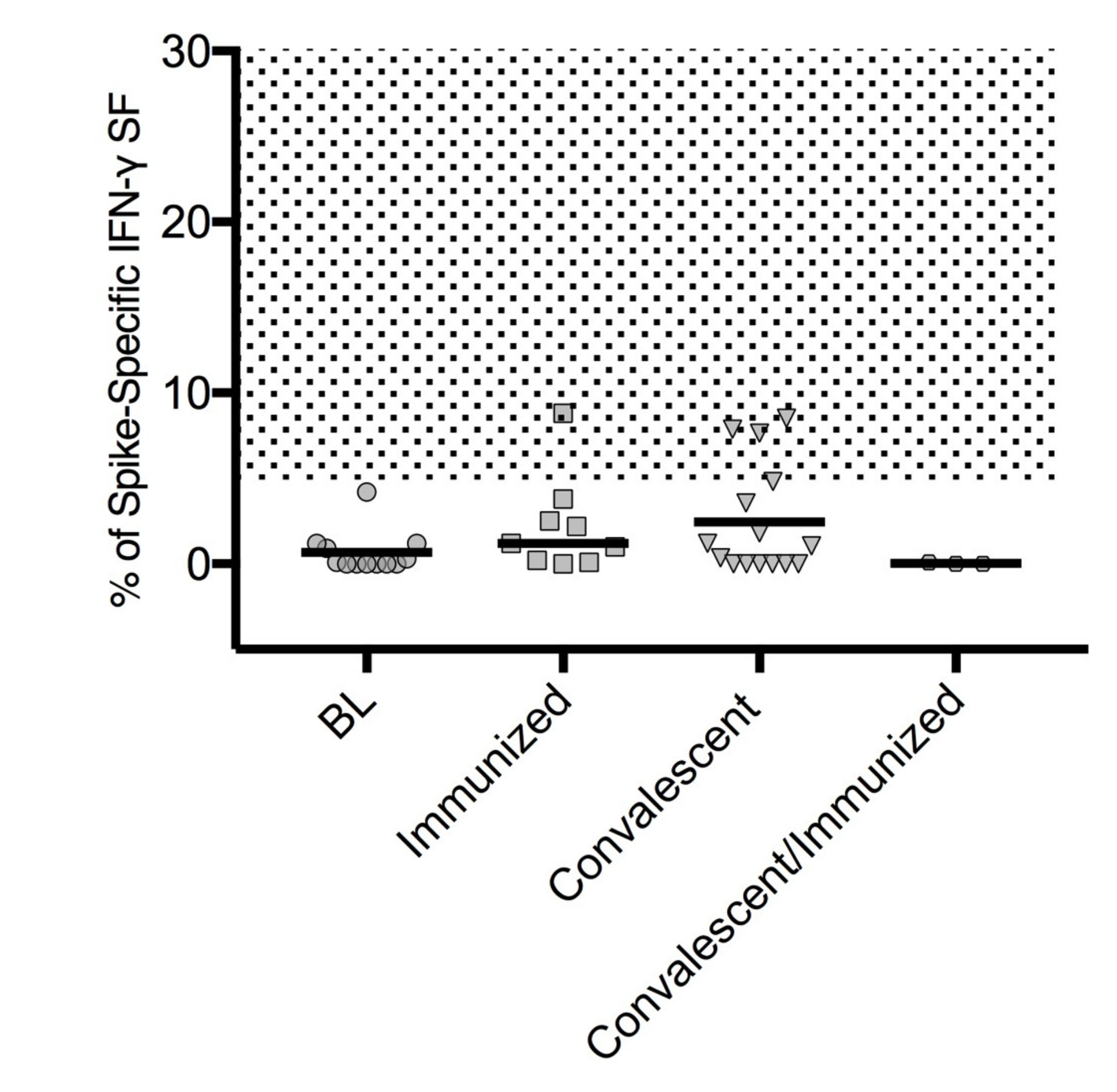
3.4. SARS-CoV-2 T Cell-Specific ELISpot Response
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Picard, C.; Gaspar, H.B.; Al-Herz, W.; Bousfiha, A.; Casanova, J.-L.; Chatila, T.; Crow, Y.J.; Cunningham-Rundles, C.; Etzioni, A.; Franco, J.L.; et al. International Union of Immunological Societies: 2017 Primary Immunodeficiency Diseases Committee Report on Inborn Errors of Immunity. J. Clin. Immunol. 2017, 38, 96–128. [Google Scholar] [CrossRef] [Green Version]
- Quinti, I.; Soresina, A.; Spadaro, G.; Martino, S.; Donnanno, S.; Agostini, C.; Claudio, P.; Franco, D.; Pesce, A.M.; Borghese, F.; et al. Long-Term Follow-Up and Outcome of a Large Cohort of Patients with Common Variable Immunodeficiency. J. Clin. Immunol. 2007, 27, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, F.A.; Barlan, I.; Chapel, H.; Costa-Carvalho, B.T.; Cunningham-Rundles, C.; De La Morena, M.T.; Espinosa-Rosales, F.J.; Hammarström, L.; Nonoyama, S.; Quinti, I.; et al. International Consensus Document (ICON): Common Variable Immunodeficiency Disorders. J. Allergy Clin. Immunol. Pract. 2015, 4, 38–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehr, C.; Kivioja, T.; Schmitt, C.; Ferry, B.; Witte, T.; Eren, E.; Vlkova, M.; Hernandez-Gonzalez, M.; Detkova, D.; Bos, P.R.; et al. The EUROclass trial: Defining subgroups in common variable immunodeficiency. Blood 2008, 111, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.-J.; Liang, W.-H.; Zhao, Y.; Liang, H.-R.; Chen, Z.-S.; Li, Y.-M.; Liu, X.-Q.; Chen, R.-C.; Tang, C.-L.; Wang, T.; et al. Comorbidity and its impact on 1590 patients with COVID-19 in China: A nationwide analysis. Eur. Respir. J. 2020, 55, 2000547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minotti, C.; Tirelli, F.; Barbieri, E.; Giaquinto, C.; Dona, D. How is immunosuppressive status affecting children and adults in SARS-CoV-2 infection? A systematic review. J. Infect. 2020, 8, e61–e66. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, K.F.A.; Tavernier, S.; Van Roy, N.; De Leeuw, E.; Declercq, J.; Bosteels, C.; Maes, B.; De Bruyne, M.; Bogaert, D.; Bosteels, V.; et al. Case Report: Convalescent Plasma, a Targeted Therapy for Patients with CVID and Severe COVID-19. Front. Immunol. 2020, 11, 596761. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.C.; Benites, B.D.; Ulaf, R.G.; Nunes, T.A.; Costa-Lima, C.; Addas-Carvalho, M.; Proenca-Modena, J.L.; Granja, F.; da Costa, V.A.; Duarte, A.D.S.S.; et al. Rapid clinical recovery of a SARS-CoV-2 infected common variable immunodeficiency patient following the infusion of COVID-19 convalescent plasma. Allergy Asthma Clin. Immunol. 2021, 17, 14. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Su, H.; Narsai, T.; Agrawal, S. SARS-CoV-2-Associated T-Cell Responses in the Presence of Humoral Immunodeficiency. Int. Arch. Allergy Immunol. 2021, 182, 195–209. [Google Scholar]
- Meyts, I.; Bucciol, G.; Quinti, I.; Neven, B.; Fischer, A.; Seoane, E.; Lopez-Granados, E.; Gianelli, C.; Robles-Marhuenda, A.; Jeandel, P.-Y.; et al. Coronavirus disease 2019 in patients with inborn errors of immunity: An international study. J. Allergy Clin. Immunol. 2020, 147, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Quinti, I.; Lougaris, V.; Milito, C.; Cinetto, F.; Pecoraro, A.; Mezzaroma, I.; Mastroianni, C.M.; Turriziani, O.; Bondioni, M.P.; Filippini, M.; et al. A possible role for B cells in COVID-19? Lesson from patients with agammaglobulinemia. J. Allergy Clin. Immunol. 2020, 146, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Milito, C.; Lougaris, V.; Giardino, G.; Punziano, A.; Vultaggio, A.; Carrabba, M.; Cinetto, F.; Scarpa, R.; Delle Piane, R.M.; Baselli, L.; et al. Clinical outcome, incidence, and SARS-CoV-2 infection-fatality rates in Italian patients with inborn errors of immunity. J. Allergy Clin. Immunol. Pract. 2021, 9, 2904–2906. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.-E.; Mathew, S.; Peluso, M.J.; Cunningham-Rundles, C. Clinical outcomes and features of COVID-19 in patients with primary immunodeficiencies in New York City. J. Allergy Clin. Immunol. Pract. 2020, 9, 490–493. [Google Scholar] [CrossRef]
- Aljaberi, R.; Wishah, K. Positive outcome in a patient with coronavirus disease 2019 and common variable immunodeficiency after intravenous immunoglobulin. Ann. Allergy Asthma Immunol. 2020, 125, 349–350. [Google Scholar] [CrossRef] [PubMed]
- Marcus, N.; Frizinsky, S.; Hagin, D.; Ovadia, A.; Hanna, S.; Farkash, M.; Maoz-Segal, R.; Agmon-Levin, N.; Broides, A.; Nahum, A.; et al. Minor Clinical Impact of COVID-19 Pandemic on Patients With Primary Immunodeficiency in Israel. Front. Immunol. 2021, 11, 614086. [Google Scholar] [CrossRef] [PubMed]
- Milito, C.; Soccodato, V.; Auria, S.; Pulvirenti, F.; Quinti, I. COVID-19 in complex common variable immunodeficiency patients affected by lung diseases. Curr. Opin. Allergy Clin. Immunol. 2021. epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Saad-Roy, C.M.; Morris, S.E.; Metcalf, C.J.; Mina, M.J.; Baker, R.E.; Farrar, J.; Holmes, E.C.; Pybus, O.G.; Graham, A.L.; Levin, S.A.; et al. Epidemiological and evolutionary considerations of SARS-CoV-2 vaccine dosing regimens. Science 2021, 372, 363–370. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Vogler, I.; Derhovanessian, E.; Kranz, L.M.; Vormehr, M.; Quandt, J.; Bidmon, N.; Ulges, A.; Baum, A.; et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature 2021, 595, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Stoesser, N.; Matthews, P.C.; Ayoubkhani, D.; Studley, R.; Bell, I.; Bell, J.I.; Newton, J.N.; Farrar, J.; Diamond, I.; et al. Antibody responses to SARS-CoV-2 vaccines in 45,965 adults from the general population of the United Kingdom. Nat. Microbiol. 2021, 6, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Raccomandazioni ad Interim sui Gruppi Target della Vaccinazione Anti-SARS-CoV-2/COVID-19. Available online: https://www.trovanorme.salute.gov.it/norme/renderPdf.spring?seriegu=SG&datagu=24/03/2021&redaz=21A01802&artp=1&art=1&subart=1&subart1=10&vers=1&prog=002 (accessed on 16 September 2021).
- ESID Positions on SARS-CoV-2 Infections in Inborn Errors of Immunity. Available online: https://esid.org/COVID-19/ESID-COVID-19-Statement (accessed on 16 September 2021).
- Kostinova, A.M.; Akhmatova, N.K.; Latysheva, E.A.; Dagil, Y.A.; Klimova, S.V.; Vlasenko, A.E.; Khromova, E.A.; Latysheva, T.V.; Kostinov, M.P. Assessment of Immunogenicity of Adjuvanted Quadrivalent Inactivated Influenza Vaccine in Healthy People and Patients With Common Variable Immune Deficiency. Front. Immunol. 2020, 11, 1876. [Google Scholar] [CrossRef] [PubMed]
- Pulvirenti, F.; Milito, C.; Cavaliere, F.M.; Mezzaroma, I.; Cinetto, F.; Quinti, I. IGA Antibody Induced by Immunization With Pneumococcal Polysaccharides Is a Prognostic Tool in Common Variable Immune Deficiencies. Front. Immunol. 2020, 11, 1283. [Google Scholar] [CrossRef]
- Seidel, M.G.; Kindle, G.; Gathmann, B.; Quinti, I.; Buckland, M.; Van Montfrans, J.; Scheible, R.; Rusch, S.; Gasteiger, L.M.; Grimbacher, B.; et al. The European Society for Immunodeficiencies (ESID) Registry Working Definitions for the Clinical Diagnosis of Inborn Errors of Immunity. J. Allergy Clin. Immunol. Pract. 2019, 7, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Salinas, A.F.; Mortari, E.P.; Terreri, S.; Quintarelli, C.; Pulvirenti, F.; Di Cecca, S.; Guercio, M.; Milito, C.; Bonanni, L.; Auria, S.; et al. SARS-CoV-2 Vaccine Induced Atypical Immune Responses in Antibody Defects: Everybody Does their Best. J. Clin. Immunol. 2021, 20, 1–14. [Google Scholar]
- Ehrhardt, G.R.; Hsu, J.; Gartland, L.; Leu, C.-M.; Zhang, S.; Davis, R.S.; Cooper, M.D. Expression of the immunoregulatory molecule FcRH4 defines a distinctive tissue-based population of memory B cells. J. Exp. Med. 2005, 202, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Fecteau, J.F.; Côté, G.; Néron, S. A new memory CD27-IgG+ B cell population in peripheral blood expressing VH genes with low frequency of somatic mutation. J. Immunol. 2006, 177, 3728–3736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintarelli, C.; Dotti, G.; Hasan, S.T.; De Angelis, B.; Hoyos, V.; Errichiello, S.; Mims, M.; Luciano, L.; Shafer, J.; Leen, A.M.; et al. High-avidity cytotoxic T lymphocytes specific for a new PRAME-derived peptide can target leukemic and leukemic-precursor cells. Blood 2011, 117, 3353–3362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization Country & Technical Guidance—Coronavirus Disease 2019. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/technical-guidance (accessed on 16 September 2021).
- Tangye, S.G.; Tarlinton, D. Memory B cells: Effectors of long-lived immune responses. Eur. J. Immunol. 2009, 39, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Mesin, L.; Ersching, J.; Victora, G.D. Germinal Center B Cell Dynamics. Immunity 2016, 45, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Braddom, A.E.; Batugedara, G.; Bol, S.; Bunnik, E.M. Potential functions of atypical memory B cells in Plasmodium-exposed individuals. Int. J. Parasitol. 2020, 50, 1033–1042. [Google Scholar] [CrossRef]
- Piano Mortari, E.; Russo, C.; Vinci, M.R.; Terreri, S.; Fernandez Salinas, A.; Piccioni, L.; Alteri, C.; Colagrossi, L.; Coltella, L.; Ranno, S.; et al. Highly Specific Memory B Cells Generation after the 2nd Dose of BNT162b2 Vaccine Compensate for the Decline of Serum Antibodies and Absence of Mucosal IgA. Cells 2021, 10, 2541. [Google Scholar] [CrossRef] [PubMed]
- Goudouris, E.S.; Pinto-Mariz, F.; Mendonça, L.O.; Aranda, C.S.; Guimarães, R.R.; Kokron, C.; Barros, M.T.; Anísio, F.; Alonso, M.L.O.; Marcelino, F.; et al. Outcome of SARS-CoV-2 Infection in 121 Patients with Inborn Errors of Immunity: A Cross-Sectional Study. J. Clin. Immunol. 2021, 23, 1–11. [Google Scholar]
- Squire, J.; Joshi, A. Seroconversion after coronavirus disease 2019 vaccination in patients with immune deficiency. Ann. Allergy Asthma Immunol. 2021, 127, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Romano, C.; Esposito, S.; Donnarumma, G.; Marrone, A. Detection of neutralizing anti-severe acute respiratory syndrome coronavirus 2 antibodies in patients with common variable immunodeficiency after immunization with messenger RNA vaccines. Ann. Allergy Asthma Immunol. 2021, 4, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Hagin, D.; Freund, T.; Navon, M.; Halperin, T.; Adir, D.; Marom, R.; Levi, I.; Benor, S.; Alcalay, Y.; Freund, N.T. Immunogenicity of Pfizer-BioNTech COVID-19 vaccine in patients with inborn errors of immunity. J. Allergy Clin. Immunol. 2021, 148, 739–749. [Google Scholar] [CrossRef]
- Steensels, D.; Pierlet, N.; Penders, J.; Mesotten, D.; Heylen, L. Comparison of SARS-CoV-2 Antibody Response Following Vaccination With BNT162b2 and mRNA-1273. JAMA 2021, 15, 1533–1535. [Google Scholar] [CrossRef]
- Driessen, G.J.; Van Zelm, M.C.; Van Hagen, P.M.; Hartwig, N.G.; Trip, M.; Warris, A.; De Vries, E.; Barendregt, B.H.; Pico, I.; Hop, W.; et al. B-cell replication history and somatic hypermutation status identify distinct pathophysiologic backgrounds in common variable immunodeficiency. Blood 2011, 118, 6814–6823. [Google Scholar] [CrossRef]
- Kalina, T.; Stuchly, J.; Janda, A.; Hrušák, O.; Růžičková, S.; Šedivá, A.; Litzman, J.; Vlkova, M. Profiling of polychromatic flow cytometry data on B-cells reveals patients’ clusters in common variable immunodeficiency. Cytom. Part A 2009, 75, 902–909. [Google Scholar] [CrossRef]
- Knox, J.J.; Buggert, M.; Kardava, L.; Seaton, K.E.; Eller, M.A.; Canaday, D.H.; Robb, M.L.; Ostrowski, M.A.; Deeks, S.G.; Slifka, M.K.; et al. T-bet+ B cells are induced by human viral infections and dominate the HIV gp140 response. JCI Insight 2017, 2, e92943. [Google Scholar] [CrossRef] [Green Version]
- Jenks, S.A.; Cashman, K.S.; Zumaquero, E.; Marigorta, U.M.; Patel, A.V.; Wang, X.; Tomar, D.; Woodruff, M.C.; Simon, Z.; Bugrovsky, R.; et al. Distinct effector B cells induced by unregulated toll-like receptor 7 contribute to pathogenic responses in systemic lupus erythematosus. Immunity 2018, 49, 725–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lederer, K.; Castaño, D.; Gómez Atria, D.; Oguin, T.H.; Wang, S.; Manzoni, T.B.; Muramatsu, H.; Hogan, M.J.; Amanat, F.; Cherubin, P.; et al. SARS-CoV-2 mRNA Vaccines Foster Potent Antigen-Specific Germinal Center Responses Associated with Neutralizing Antibody Generation. Immunity 2020, 53, 1281–1295. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.N.; Devlin, J.C.; Buus, T.B.; Koide, A.; Cornelius, A.; Samanovic, M.I.; Herrera, A.; Zhang, C.; Desvignes, L.; Odum, N.; et al. Discrete immune response signature to SARS-CoV-2 mRNA vaccination versus infection. medRxiv 2021. [Google Scholar] [CrossRef]
- Casulli, S.; Coignard-Biehler, H.; Amazzough, K.; Shoai-Tehrani, M.; Bayry, J.; Mahlaoui, N.; Elbim, C.; Kaveri, S.V. Defective functions of polymorphonuclear neutrophils in patients with common variable immunodeficiency. Immunol. Res. 2014, 60, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, C.M.; Muskus, C.; Arango, J.; Patino, P.; Montoya, C.J. Quantitative and functional evaluation of innate immune responses in patients with common variable immunodeficiency. J. Investig. Allergol. Clin. Immunol. 2011, 21, 207–215. [Google Scholar] [PubMed]
- Park, J.; Munagala, I.; Xu, H.; Blankenship, D.; Maffucci, P.; Chaussabel, D.; Banchereau, J.; Pascual, V.; Cunningham-Rundles, C. Interferon Signature in the Blood in Inflammatory Common Variable Immune Deficiency. PLoS ONE 2013, 8, e74893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizi, G.; Rezaei, N.; Kiaee, F.; Tavakolinia, N.; Yazdani, R.; Mirshafiey, A.; Aghamohammadi, A. T Cell Abnormalities in Common Variable Immunodeficiency. J. Investig. Allergol. Clin. Immunol. 2016, 26, 233–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viallard, J.-F.; Ruiz, C.; Guillet, M.; Pellegrin, J.-L.; Moreau, J.-F. Perturbations of the CD8+ T-cell repertoire in CVID patients with complications. Results Immunol. 2013, 3, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlkova, M.; Ticha, O.; Nechvatalova, J.; Kalina, T.; Litzman, J.; Mauri, C.; Blair, P.A. Regulatory B cells in CVID patients fail to suppress multifunctional IFN-γ+TNF-α+CD4+ T cells differentiation. Clin. Immunol. 2015, 160, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Ochtrop, M.L.G.; Goldacker, S.; May, A.M.; Rizzi, M.; Draeger, R.; Hauschke, D.; Stehfest, C.; Warnatz, K.; Goebel, H.; Technau-Ihling, K.; et al. T and B lymphocyte abnormalities in bone marrow biopsies of common variable immunodeficiency. Blood 2011, 118, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameratunga, R.; Longhurst, H.; Steele, R.; Lehnert, K.; Leung, E.; Brooks, A.E.; Woon, S.-T. Common Variable Immunodeficiency Disorders, T-Cell Responses to SARS-CoV-2 Vaccines, and the Risk of Chronic COVID-19. J. Allergy Clin. Immunol. Pract. 2021, 9, 3575–3583. [Google Scholar] [CrossRef]
- Kinoshita, H.; Durkee-Shock, J.; Jensen-Wachspress, M.; Kankate, V.V.; Lang, H.; Lazarski, C.A.; Keswani, A.; Webber, K.C.; Montgomery-Recht, K.; Walkiewicz, M.; et al. Robust Antibody and T Cell Responses to SARS-CoV-2 in Patients with Antibody Deficiency. J. Clin. Immunol. 2021, 41, 1146–1153. [Google Scholar] [CrossRef]
- Friedmann, D.; Goldacker, S.; Peter, H.-H.; Warnatz, K. Preserved Cellular Immunity Upon Influenza Vaccination in Most Patients with Common Variable Immunodeficiency. J. Allergy Clin. Immunol. Pract. 2020, 8, 2332–2340. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health Circular “Vaccination of Individuals with SARS-CoV.2 Infection”. Available online: https://www.trovanorme.salute.gov.it/norme/renderNormsanPdf?anno=2021&codLeg=79033&parte=1%20&serie=null (accessed on 16 September 2021).




{kind=link}
{kind=link}
{kind=link}
| Group 1 Immunized n = 38 | Group 2 Convalescents n = 34 | Group 3 Convalescent/Immunized n = 20 | Groups 1 vs. 2 p-Value | Groups 1 vs. 3 p-Value | Groups 2 vs. 3 p-Value | |
|---|---|---|---|---|---|---|
| Sex, n (%) | 25 (66) | 16 (47) | 12 (60) | 0.157 | 0.776 | 0.408 |
| Age, years, median (IQR) | 54.5 (42.5–60.0) | 49.5 (44.7–59.2) | 49.5 (44.2–62.7) | 0.840 | 0.973 | 0.943 |
| PAD-related complication | ||||||
| Autoimmunity, n (%) | 18 (47) | 9 (26) | 7 (35) | 0.094 | 0.414 | 0.550 |
| Chronic lung disease, n (%) | 8 (21) | 20 (58) | 10 (50) | 0.001 | 0.036 | 0.560 |
| Cancer, n (%) | 2 (5) | 2 (6) | 2 (10) | 1.000 | 0.602 | 0.622 |
| Immunosuppressive treatment, n (%) | 10 (26) | 3 (9) | 2 (10) | 0.069 | 0.186 | 1.000 |
| IgG, g/L, median (IQR) | 6.9 (5.8–8.1) | 7.5 (6.5–8.8) | 7.6 (7.2–8.8) | 0.069 | 0.122 | 0.881 |
| IgA, mg/dL, median (IQR) | 7.0 (0–25.0) | 6.0 (0–28.0) | 4.5 (0–12.8) | 0.541 | 0.599 | 0.401 |
| IgM, mg/dL, median (IQR) | 19.6 (4–42) | 20.0 (5–25.1) | 9.5 (4–25) | 0.576 | 0.355 | 0.648 |
| CD3+, cell/mm3, median (IQR) | 924 (748–1512) | 974 (675–1548) | 879 (639–1412) | 0.513 | 0.255 | 0.335 |
| CD4+, cell/mm3, median (IQR) | 452 (356–811) | 511 (356–811) | 479 (335–838) | 0.756 | 0.801 | 0.992 |
| CD8+, cell/mm3, median (IQR) | 421 (186–692) | 369 (269–710) | 297 (242–521) | 0.741 | 0.281 | 0.095 |
| CD19+, cell/mm3, median (IQR) | 87 (20–184) | 91 (29–172) | 55 (30–131) | 0.564 | 0.150 | 0.272 |
| CD19+CD27+ IgM–IgD–, %, median (IQR) | 3.5 (1–7.7) | 2.0 (0–5.0) | 2.5 (0–5.5) | 0.649 | 0.921 | 0.771 |
| CD19+CD27+IgM–IgD–, cell/mm3, median (IQR) | 1 (0–5.8) | 1.6 (0–4.1) | 2.3 (0.1–4.6) | 0.480 | 0.474 | 0.739 |
| Atypical MBC (ATM) CD19+CD24-CD27–CD38–CD21–, % of CD19+, median (IQR) | 3.69 (2.8–8.1) | 3.4 (2.3–5.2) | 3.1 (2.9–4.1) | 0.469 | 0.613 | 0.863 |
| ID | Age (Range) | Time Range of SARS-CoV-2 Infection | SARS-CoV-2 Infection Severity | SARS-CoV-2Associated Symptoms | Pneumonia | Days of SARS-CoV-2 qPCR Positivity | Additional COVID-19 Specific Therapy | Outcome | BNT162b2 Immunization |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 30–39 | March–May 2020 | moderate | fever and cough dyspnea | yes | 45 | lopinavir/ritonavir, tocilizumab, and dexamethasone | recovery | no |
| 2 | 50–59 | June–September 2020 | asymptomatic | no | 79 | no | recovery | yes, 180 days from infection | |
| 3 | 40–49 | January–April 2021 | asymptomatic | no | 30 | no | recovery | no | |
| 4 | 60–60 | October–December 2020 | asymptomatic | no | 14 | no | recovery | no | |
| 5 | 50–59 | October–December 2020 | asymptomatic | no | 81 | no | recovery | no | |
| 6 | 50–59 | June–September 2020 | asymptomatic | no | 51 | no | recovery | no | |
| 7 | 30–39 | January–April 2021 | moderate | fever, cough, and dyspnea | no | 23 | no | recovery | no |
| 8 | 40–49 | October–December 2020 | moderate | fever, cough, and dyspnea | yes | 75 | lopinavir/ritonavir, tocilizumab, and dexamethasone | dyspnoea and O2 therapy | no |
| 9 | 30–39 | January–April 2021 | mild | fever | no | 21 | dexamethasone | recovery | no |
| 10 | 40–49 | January–April 2021 | moderate | fever, cough, and dyspnea | yes | 22 | remdesivir and dexamethasone | recovery | no |
| 11 | 60–69 | January–April 2021 | mild | fever | no | 17 | bamlanivimab/ etesevimab | recovery | no |
| 12 | 50–59 | January–April 2021 | moderate | fever, cough, and dyspnea | yes | 112 | calsirimab/imdevimab, remdesivir, and dexamethasone | recovery | no |
| 13 | 40–49 | January–April 2021 | moderate | fever, cough, and dyspnea | yes | 40 | calsirimab/imdevimab, remdesivir, and dexamethasone | dyspnoea and O2 therapy | no |
| 14 | 60–69 | January–April 2021 | asymptomatic | no | 33 | bamlanivimab/ etesevimab | recovery | no | |
| 15 | 50–59 | January–April 2021 | asymptomatic | no | 23 | bamlanivimab | recovery | no | |
| 16 | 40–49 | October–December 2020 | asymptomatic | no | NA | no | recovery | yes, 150 days from infection | |
| 17 | 30–39 | October–December 2020 | asymptomatic | no | NA | no | recovery | yes, 150 days from infection | |
| 18 | 40–49 | January–April 2021 | moderate | fever and mild dyspnea | yes | 49 | dexamethasone and azithromycin | recovery | yes, 120 days from infection |
| 19 | 20–29 | October–December 2020 | asymptomatic | no | 15 | no | recovery | yes, 120 days from infection | |
| 20 | 50–59 | March–May 2020 | severe | fever, cough, and severe dyspnea | yes | 10 | lopinavir/ritonavir, azithromycin, and hydroxychloroquine | recovery | yes, 330 days from infection |
| 21 | 60–69 | October–December 2020 | moderate | fever and mild dyspnea | yes | 60 | NA | recovery | yes, 150 days from infection |
| 22 | 50–59 | January–April 2021 | mild | ageusia | no | 9 | no | recovery | yes, 90 days from infection |
| 23 | 50–59 | October–December 2020 | mild | fever | no | NA | no | recovery | yes, 180 days from infection |
| 24 | 40–49 | January–April 2021 | asymptomatic | no | 22 | no | recovery | yes, 90 days from infection | |
| 25 | 40–49 | January–April 2021 | asymptomatic | no | 9 | no | recovery | yes, 120 days from infection | |
| 26 | 50–59 | October–December 2020 | asymptomatic | no | 16 | no | recovery | yes, 120 days from infection | |
| 27 | 40–49 | October–December 2020 | mild | fever and cough | no | 23 | dexamethasone and azithromycin | recovery | yes, 150 days from infection |
| 28 | 60–69 | October–December 2020 | mild | fever and arthralgia | no | 20 | no | recovery | yes, 150 days from infection |
| 29 | ≥70 | October–December 2020 | moderate | fever, mild dyspnea, and ageusia | no | 27 | dexamethasone and azithromycin | dyspnoea | yes, 90 days from infection |
| 30 | 40–49 | October–December 2020 | asymptomatic | no | 30 | no | recovery | yes, 150 days from infection | |
| 31 | 20–29 | January–April 2021 | mild | ageusia and anosmia | no | 59 | no | recovery | yes, 120 days from infection |
| 32 | 60–69 | October–December 2020 | mild | anosmia | no | 34 | no | recovery | yes, 150 days from infection |
| 33 | 60–69 | January–April 2021 | mild | arthralgia | no | 12 | no | dyspnoea | yes, 120 days from infection |
| 34 | 40–49 | October–December 2020 | severe | fever and moderate dyspnea | yes | 53 | dexamethasone and azithromycin | recovery | yes, 120 days from infection |
| IgG S1 (OD Ratio) | MBC S + (% of CD24 + CD27 + CD38–Inside CD19 + Cells) | ATM S + (% of CD24–CD27–CD38–CD21–Inside CD19+ Cells) | |
|---|---|---|---|
| Baseline, median (IQR) | 0.11 (0.08–0.18) | 0 (0–0.98) | 0 (0–0) |
| Immunized, median (IQR) | 0.30 (0.09–5.39) | 0.07 (0–0.13) | 0 (0–0.27) |
| Convalescent, median (IQR) | 2.5 (1.07–4.04) | 0.16 (0.22) | 0 (0–0) |
| Convalescent/immunized median (IQR) | 9.6 (2.75–12.33) | 0.25 (1.17–0.31) | 0.50 (0.39–1.40) |
| MoAbs treatment, median (IQR) | 6.91 (6.56–9.42) | Nap | Nap |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pulvirenti, F.; Fernandez Salinas, A.; Milito, C.; Terreri, S.; Piano Mortari, E.; Quintarelli, C.; Di Cecca, S.; Lagnese, G.; Punziano, A.; Guercio, M.; et al. B Cell Response Induced by SARS-CoV-2 Infection Is Boosted by the BNT162b2 Vaccine in Primary Antibody Deficiencies. Cells 2021, 10, 2915. https://doi.org/10.3390/cells10112915
Pulvirenti F, Fernandez Salinas A, Milito C, Terreri S, Piano Mortari E, Quintarelli C, Di Cecca S, Lagnese G, Punziano A, Guercio M, et al. B Cell Response Induced by SARS-CoV-2 Infection Is Boosted by the BNT162b2 Vaccine in Primary Antibody Deficiencies. Cells. 2021; 10(11):2915. https://doi.org/10.3390/cells10112915
Chicago/Turabian StylePulvirenti, Federica, Ane Fernandez Salinas, Cinzia Milito, Sara Terreri, Eva Piano Mortari, Concetta Quintarelli, Stefano Di Cecca, Gianluca Lagnese, Alessandra Punziano, Marika Guercio, and et al. 2021. "B Cell Response Induced by SARS-CoV-2 Infection Is Boosted by the BNT162b2 Vaccine in Primary Antibody Deficiencies" Cells 10, no. 11: 2915. https://doi.org/10.3390/cells10112915
APA StylePulvirenti, F., Fernandez Salinas, A., Milito, C., Terreri, S., Piano Mortari, E., Quintarelli, C., Di Cecca, S., Lagnese, G., Punziano, A., Guercio, M., Bonanni, L., Auria, S., Villani, F., Albano, C., Locatelli, F., Spadaro, G., Carsetti, R., & Quinti, I. (2021). B Cell Response Induced by SARS-CoV-2 Infection Is Boosted by the BNT162b2 Vaccine in Primary Antibody Deficiencies. Cells, 10(11), 2915. https://doi.org/10.3390/cells10112915