
Liver Regeneration by Hematopoietic Stem Cells: Have We Reached the End of the Road?


Abstract
:1. Introduction
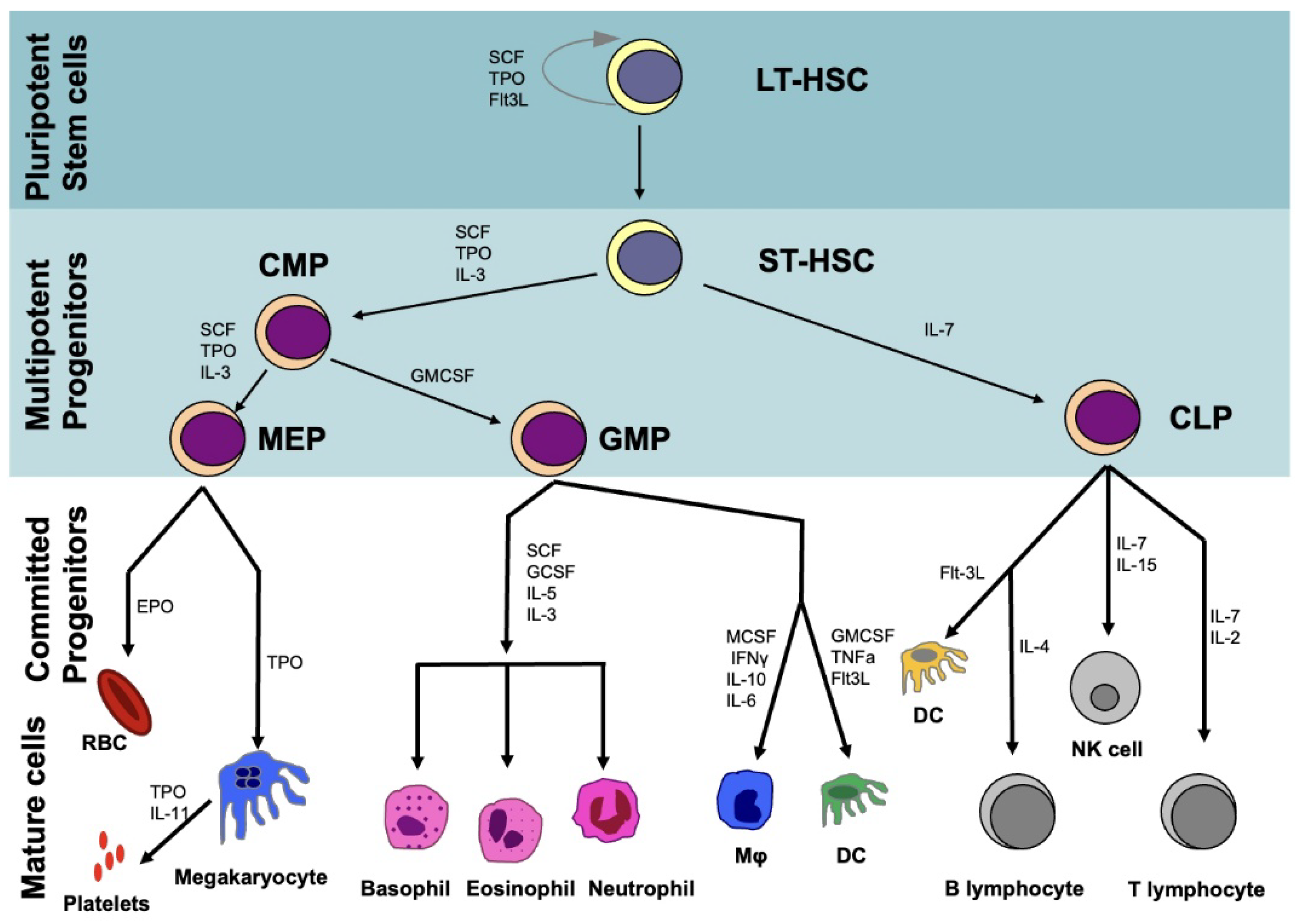
2. Hematopoietic System
2.1. Hematopoietic Stem Cells (HSCs)
2.2. Embryonic Hematopoiesis
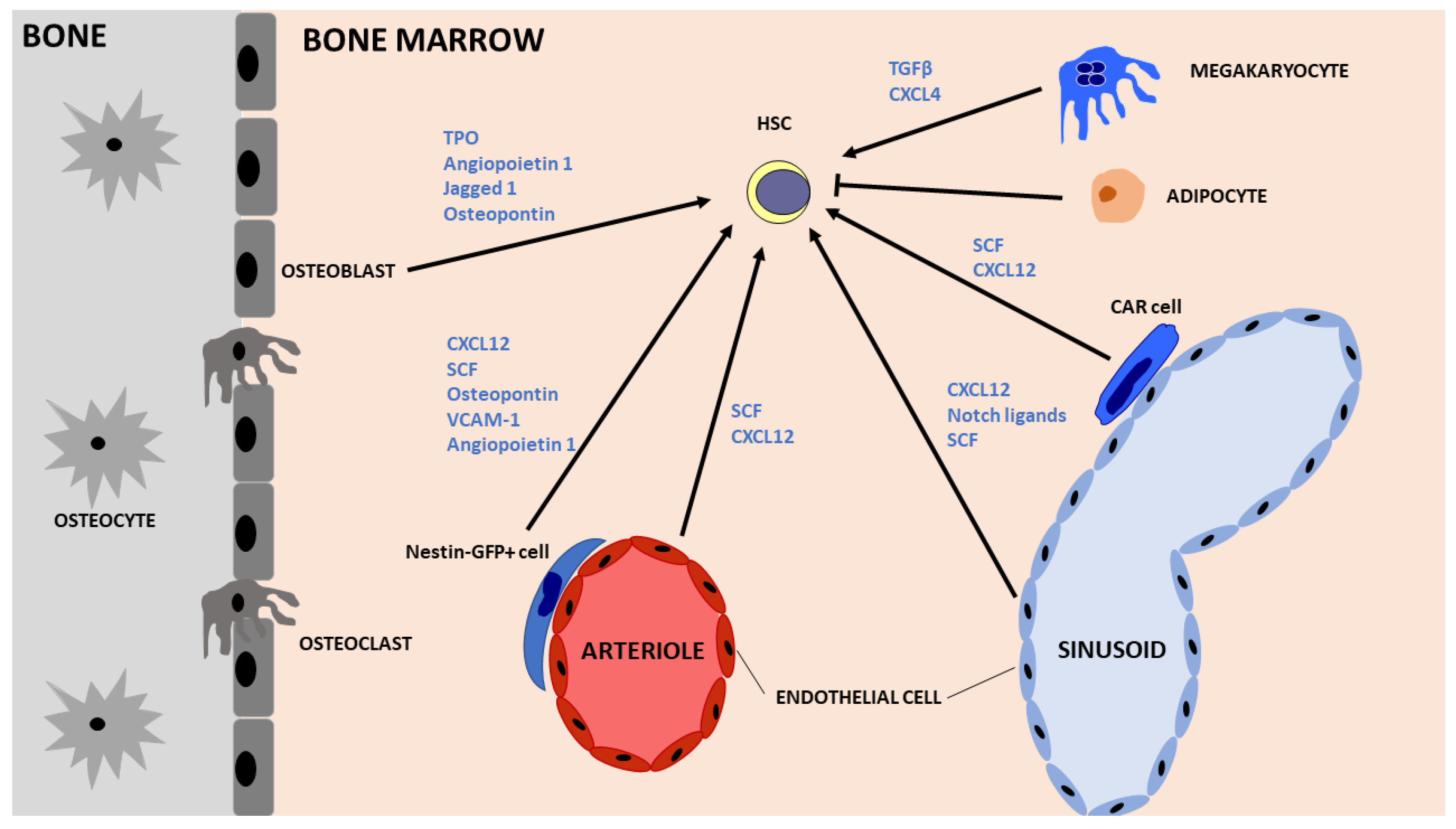
3. Fetal Liver Crosstalk with HSCs
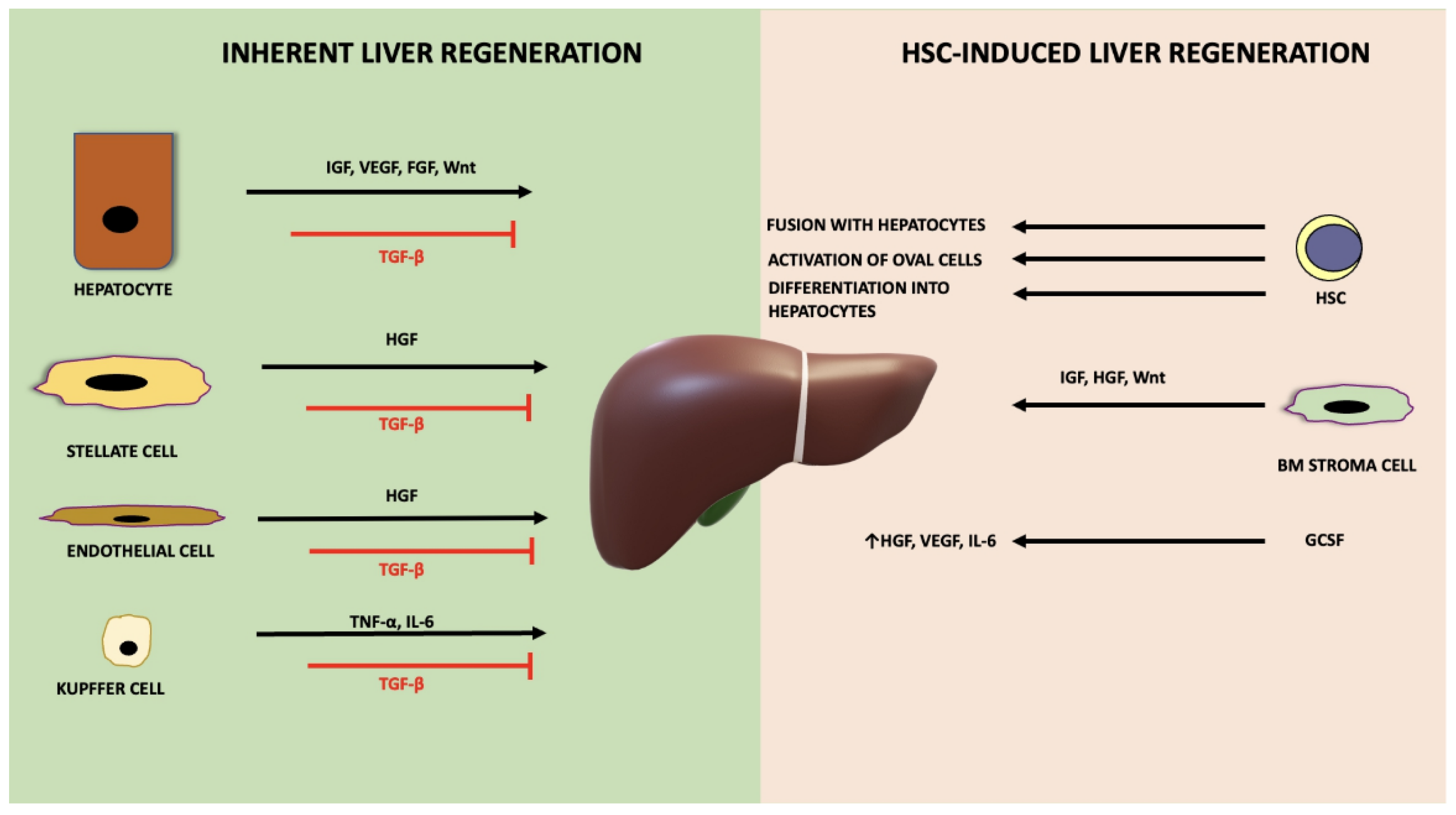
4. Inherent Liver Regeneration
5. HSC-Mediated Liver Regeneration
6. Clinical Trials of HSC Transplantation or Mobilisation in Patients with Liver Disease
7. Discussion
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Trefts, E.; Gannon, M.; Wasserman, D.H. The Liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Furuya, K.; Zheng, Y.-W.; Oda, T. Novel Alternative Transplantation Therapy for Orthotopic Liver Transplantation in Liver Failure: A Systematic Review. World J. Transplant. 2020, 10, 64–78. [Google Scholar] [CrossRef]
- Iansante, V.; Mitry, R.R.; Filippi, C.; Fitzpatrick, E.; Dhawan, A. Human Hepatocyte Transplantation for Liver Disease: Current Status and Future Perspectives. Pediatr. Res. 2018, 83, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Giancotti, A.; D’Ambrosio, V.; Corno, S.; Pajno, C.; Carpino, G.; Amato, G.; Vena, F.; Mondo, A.; Spiniello, L.; Monti, M.; et al. Current Protocols and Clinical Efficacy of Human Fetal Liver Cell Therapy in Patients with Liver Disease: A Literature Review. Cytotherapy 2022, 24, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Nikokiraki, C.; Psaraki, A.; Roubelakis, M.G. The Potential Clinical Use of Stem/Progenitor Cells and Organoids in Liver Diseases. Cells 2022, 11, 1410. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, J.; Hackl, V.; Esser, H.; Meszaros, A.T.; Fodor, M.; Öfner, D.; Troppmair, J.; Schneeberger, S.; Hautz, T. Cell-Based Regeneration and Treatment of Liver Diseases. Int. J. Mol. Sci. 2021, 22, 10276. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Prolif. 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Trohatou, O.; Roubelakis, M.G. Mesenchymal Stem/Stromal Cells in Regenerative Medicine: Past, Present, and Future. Cell. Reprogramming 2017, 19, 217–224. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, S.; Shi, X.; Cao, H.; Li, L. A Pooled Analysis of Mesenchymal Stem Cell-Based Therapy for Liver Disease. Stem Cell Res. Ther. 2018, 9, 72. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, Y.; Wu, X.; Xu, X.; Niu, J. The Assessment of Mesenchymal Stem Cells Therapy in Acute on Chronic Liver Failure and Chronic Liver Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Clinical Trials. Stem Cell Res. Ther. 2022, 13, 204. [Google Scholar] [CrossRef]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afshari, A.; Shamdani, S.; Uzan, G.; Naserian, S.; Azarpira, N. Different Approaches for Transformation of Mesenchymal Stem Cells into Hepatocyte-like Cells. Stem Cell Res. Ther. 2020, 11, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Yuan, L.; An, Z.; Shi, D.; Xin, J.; Jiang, J.; Ren, K.; Chen, J.; Guo, B.; Zhou, X.; et al. DLL4 Restores Damaged Liver by Enhancing HBMSC Differentiation into Cholangiocytes. Stem Cell Res. 2020, 47, 101900. [Google Scholar] [CrossRef]
- Tsuchiya, A.; Takeuchi, S.; Watanabe, T.; Yoshida, T.; Nojiri, S.; Ogawa, M.; Terai, S. Mesenchymal Stem Cell Therapies for Liver Cirrhosis: MSCs as “Conducting Cells” for Improvement of Liver Fibrosis and Regeneration. Inflamm. Regen. 2019, 39, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psaraki, A.; Ntari, L.; Karakostas, C.; Korrou-Karava, D.; Roubelakis, M.G. Extracellular Vesicles Derived from Mesenchymal Stem/Stromal Cells: The Regenerative Impact in Liver Diseases. Hepatology 2022, 75, 1590–1603. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Hannan, N.R.F.; Segeritz, C.-P.; Touboul, T.; Vallier, L. Production of Hepatocyte-like Cells from Human Pluripotent Stem Cells. Nat. Protoc. 2013, 8, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Basma, H.; Soto–Gutiérrez, A.; Yannam, G.R.; Liu, L.; Ito, R.; Yamamoto, T.; Ellis, E.; Carson, S.D.; Sato, S.; Chen, Y.; et al. Differentiation and Transplantation of Human Embryonic Stem Cell–Derived Hepatocytes. Gastroenterology 2009, 136, 990–999.e4. [Google Scholar] [CrossRef] [Green Version]
- Tolosa, L.; Caron, J.; Hannoun, Z.; Antoni, M.; López, S.; Burks, D.; Castell, J.V.; Weber, A.; Gomez-Lechon, M.-J.; Dubart-Kupperschmitt, A. Transplantation of HESC-Derived Hepatocytes Protects Mice from Liver Injury. Stem Cell Res. Ther. 2015, 6, 246. [Google Scholar] [CrossRef] [Green Version]
- Tasnim, F.; Xing, J.; Huang, X.; Mo, S.; Wei, X.; Tan, M.-H.; Yu, H. Generation of Mature Kupffer Cells from Human Induced Pluripotent Stem Cells. Biomaterials 2019, 192, 377–391. [Google Scholar] [CrossRef]
- Povero, D.; Pinatel, E.M.; Leszczynska, A.; Goyal, N.P.; Nishio, T.; Kim, J.; Kneiber, D.; de Araujo Horcel, L.; Eguchi, A.; Ordonez, P.M.; et al. Human Induced Pluripotent Stem Cell–Derived Extracellular Vesicles Reduce Hepatic Stellate Cell Activation and Liver Fibrosis. JCI Insight 2019, 4, e125652. [Google Scholar] [CrossRef]
- Liu, X.; Li, W.; Fu, X.; Xu, Y. The Immunogenicity and Immune Tolerance of Pluripotent Stem Cell Derivatives. Front. Immunol. 2017, 8, 645. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, M.; Hayashizaki, Y.; Murakawa, Y. Genomic Instability of IPSCs: Challenges Towards Their Clinical Applications. Stem Cell Rev. Rep. 2017, 13, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of Putative Progenitor Endothelial Cells for Angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef]
- Lan, L.; Liu, R.; Qin, L.-Y.; Cheng, P.; Liu, B.-W.; Zhang, B.-Y.; Ding, S.-Z.; Li, X.-L. Transplantation of Bone Marrow-Derived Endothelial Progenitor Cells and Hepatocyte Stem Cells from Liver Fibrosis Rats Ameliorates Liver Fibrosis. World J. Gastroenterol. 2018, 24, 237–247. [Google Scholar] [CrossRef]
- Kaur, S.; Tripathi, D.; Dongre, K.; Garg, V.; Rooge, S.; Mukopadhyay, A.; Sakhuja, P.; Sarin, S.K. Increased Number and Function of Endothelial Progenitor Cells Stimulate Angiogenesis by Resident Liver Sinusoidal Endothelial Cells (SECs) in Cirrhosis through Paracrine Factors. J. Hepatol. 2012, 57, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, E.; Kin, M.; Torimura, T.; Nakamura, T.; Kumemura, H.; Hanada, S.; Hisamoto, T.; Yoshida, T.; Kawaguchi, T.; Baba, S.; et al. Endothelial Progenitor Cell Transplantation Improves the Survival Following Liver Injury in Mice. Gastroenterology 2006, 130, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Liu, Z.-D.; Wu, N.; Cong, X.; Fei, R.; Chen, H.-S.; Wei, L. Transplanted Endothelial Progenitor Cells Ameliorate Carbon Tetrachloride-Induced Liver Cirrhosis in Rats. Liver Transplant. 2009, 15, 1092–1100. [Google Scholar] [CrossRef]
- D’Avola, D.; Fernández-Ruiz, V.; Carmona-Torre, F.; Méndez, M.; Pérez-Calvo, J.; Prósper, F.; Andreu, E.; Herrero, J.I.; Iñarrairaegui, M.; Fuertes, C.; et al. Phase 1–2 Pilot Clinical Trial in Patients with Decompensated Liver Cirrhosis Treated with Bone Marrow–Derived Endothelial Progenitor Cells. Transl. Res. 2017, 188, 80–91.e2. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhou, L.; Ma, X.; Ma, W.; Wang, C.; Lu, Y.; Chen, Y.; An, L.; An, W.; Yang, Y. Monitoring of Intrasplenic Hepatocyte Transplantation for Acute-on-Chronic Liver Failure: A Prospective Five-Year Follow-Up Study. Transplant. Proc. 2014, 46, 192–198. [Google Scholar] [CrossRef]
- Bilir, B.; Guinette, D.; Karrer, F.; Kumpe, D.; Krysl, J.; Stephens, J.; Mcgavran, L.; Ostrowska, A.; Durham, J. Hepatocyte Transplantation in Acute Liver Failure. Liver Transplant. 2000, 6, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Pareja, E.; Gomez-Lechon, M.J.; Cortes, M.; Bonora-Centelles, A.; Castell, J.V.; Mir, J. Human Hepatocyte Transplantation in Patients with Hepatic Failure Awaiting a Graft. Eur. Surg. Res. 2013, 50, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Overturf, K.; Al-Dhalimy, M.; Tanguay, R.; Brantly, M.; Ou, C.-N.; Finegold, M.; Grompe, M. Hepatocytes Corrected by Gene Therapy are Selected in Vivo in a Murine Model of Hereditary Tyrosinaemia Type I. Nat. Genet. 1996, 12, 266–273. [Google Scholar] [CrossRef]
- Ding, J.; Yannam, G.R.; Roy-Chowdhury, N.; Hidvegi, T.; Basma, H.; Rennard, S.I.; Wong, R.J.; Avsar, Y.; Guha, C.; Perlmutter, D.H.; et al. Spontaneous Hepatic Repopulation in Transgenic Mice Expressing Mutant Human A1-Antitrypsin by Wild-Type Donor Hepatocytes. J. Clin. Investig. 2011, 121, 1930–1934. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A.; Chaijitraruch, N.; Fitzpatrick, E.; Bansal, S.; Filippi, C.; Lehec, S.C.; Heaton, N.D.; Kane, P.; Verma, A.; Hughes, R.D.; et al. Alginate Microencapsulated Human Hepatocytes for the Treatment of Acute Liver Failure in Children. J. Hepatol. 2020, 72, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Habibullah, C.M.; Syed, I.H.; Qamar, A.; Taher-Uz, Z. Human fetal hepatocyte transplantation in patients with fulminant hepatic failure. Transplantation 1994, 58, 951–952. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Habeeb, A.; Parveen, N.; Naseem, B.; Babu, R.P.; Capoor, A.K.; Habibullah, C.M. Peritoneal Transplantation of Human Fetal Hepatocytes for the Treatment of Acute Fatty Liver of Pregnancy: A Case Report. Trop. Gastroenterol. 2005, 25, 141–143. [Google Scholar]
- Patterson, A.M.; Pelus, L.M. G-CSF in Stem Cell Mobilization: New Insights, New Questions. Ann. Blood 2016, 2, 10. [Google Scholar] [CrossRef]
- Sawai, C.M.; Babovic, S.; Upadhaya, S.; Knapp, D.J.H.F.; Lavin, Y.; Lau, C.M.; Goloborodko, A.; Feng, J.; Fujisaki, J.; Ding, L.; et al. Hematopoietic Stem Cells are the Major Source of Multilineage Hematopoiesis in Adult Animals. Immunity 2016, 45, 597–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Zheng, Z.; Cheng, T. New Paradigms on Hematopoietic Stem Cell Differentiation. Protein Cell 2020, 11, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, A.C.; Igarashi, K.J.; Nakauchi, H. Haematopoietic Stem Cell Self-Renewal In Vivo and Ex Vivo. Nat. Rev. Genet. 2020, 21, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Gurudutta, G. Epigenetic Regulation of Hematopoietic Stem Cells. Int. J. Stem Cells 2016, 9, 36–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouzolles, M.; Oburoglu, L.; Taylor, N.; Zimmermann, V.S. Hematopoietic Stem Cell Lineage Specification. Curr. Opin. Hematol. 2016, 23, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.S.; Hu, W.; Park, C.Y. The Role of MicroRNAs in Hematopoietic Stem Cell and Leukemic Stem Cell Function. Ther. Adv. Hematol. 2011, 2, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K.; Mauch, P.; Vergilio, J.-A.; Sackstein, R.; Down, J.D. Distribution of Hematopoietic Stem Cells in the Bone Marrow According to Regional Hypoxia. Proc. Natl. Acad. Sci. USA 2007, 104, 5431–5436. [Google Scholar] [CrossRef] [Green Version]
- Nombela-Arrieta, C.; Pivarnik, G.; Winkel, B.; Canty, K.J.; Harley, B.; Mahoney, J.E.; Park, S.-Y.; Lu, J.; Protopopov, A.; Silberstein, L.E. Quantitative Imaging of Haematopoietic Stem and Progenitor Cell Localization and Hypoxic Status in the Bone Marrow Microenvironment. Nat. Cell Biol. 2013, 15, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.; Frenette, P.S. Haematopoietic Stem Cell Activity and Interactions with the Niche. Nat. Rev. Mol. Cell Biol. 2019, 20, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Luis, T.C.; Naber, B.A.E.; Roozen, P.P.C.; Brugman, M.H.; de Haas, E.F.E.; Ghazvini, M.; Fibbe, W.E.; van Dongen, J.J.M.; Fodde, R.; Staal, F.J.T. Canonical Wnt Signaling Regulates Hematopoiesis in a Dosage-Dependent Fashion. Cell Stem Cell 2011, 9, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamma, R.; Ribatti, D. Bone Niches, Hematopoietic Stem Cells, and Vessel Formation. Int. J. Mol. Sci. 2017, 18, 151. [Google Scholar] [CrossRef] [Green Version]
- Notta, F.; Doulatov, S.; Laurenti, E.; Poeppl, A.; Jurisica, I.; Dick, J.E. Isolation of Single Human Hematopoietic Stem Cells Capable of Long-Term Multilineage Engraftment. Science 2011, 333, 218–221. [Google Scholar] [CrossRef]
- Anjos-Afonso, F.; Currie, E.; Palmer, H.G.; Foster, K.E.; Taussig, D.C.; Bonnet, D. CD34− Cells at the Apex of the Human Hematopoietic Stem Cell Hierarchy Have Distinctive Cellular and Molecular Signatures. Cell Stem Cell 2013, 13, 161–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjos-Afonso, F.; Buettner, F.; Mian, S.A.; Rhys, H.; Perez-Lloret, J.; Garcia-Albornoz, M.; Rastogi, N.; Ariza-McNaughton, L.; Bonnet, D. Single Cell Analyses Identify a Highly Regenerative and Homogenous Human CD34+ Hematopoietic Stem Cell Population. Nat. Commun. 2022, 13, 2048. [Google Scholar] [CrossRef]
- Knapp, D.J.H.F.; Hammond, C.A.; Hui, T.; van Loenhout, M.T.J.; Wang, F.; Aghaeepour, N.; Miller, P.H.; Moksa, M.; Rabu, G.M.; Beer, P.A.; et al. Single-Cell Analysis Identifies a CD33+ Subset of Human Cord Blood Cells with High Regenerative Potential. Nat. Cell Biol. 2018, 20, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Tavian, M.; Hallais, M.F.; Peault, B. Emergence of Intraembryonic Hematopoietic Precursors in the Pre-Liver Human Embryo. Development 1999, 126, 793–803. [Google Scholar] [CrossRef]
- Palis, J.; Robertson, S.; Kennedy, M.; Wall, C.; Keller, G. Development of Erythroid and Myeloid Progenitors in the Yolk Sac and Embryo Proper of the Mouse. Development 1999, 126, 5073–5084. [Google Scholar] [CrossRef] [PubMed]
- Vink, C.S.; Mariani, S.A.; Dzierzak, E. Embryonic Origins of the Hematopoietic System: Hierarchies and Heterogeneity. Hemasphere 2022, 6, e737. [Google Scholar] [CrossRef]
- Ajami, B.; Samusik, N.; Wieghofer, P.; Ho, P.P.; Crotti, A.; Bjornson, Z.; Prinz, M.; Fantl, W.J.; Nolan, G.P.; Steinman, L. Single-Cell Mass Cytometry Reveals Distinct Populations of Brain Myeloid Cells in Mouse Neuroinflammation and Neurodegeneration Models. Nat. Neurosci. 2018, 21, 541–551. [Google Scholar] [CrossRef]
- Soares-da-Silva, F.; Freyer, L.; Elsaid, R.; Burlen-Defranoux, O.; Iturri, L.; Sismeiro, O.; Pinto-do-Ó, P.; Gomez-Perdiguero, E.; Cumano, A. Yolk Sac, but Not Hematopoietic Stem Cell–Derived Progenitors, Sustain Erythropoiesis throughout Murine Embryonic Life. J. Exp. Med. 2021, 218, e20201729. [Google Scholar] [CrossRef]
- McGrath, K.E.; Frame, J.M.; Fegan, K.H.; Bowen, J.R.; Conway, S.J.; Catherman, S.C.; Kingsley, P.D.; Koniski, A.D.; Palis, J. Distinct Sources of Hematopoietic Progenitors Emerge before HSCs and Provide Functional Blood Cells in the Mammalian Embryo. Cell Rep. 2015, 11, 1892–1904. [Google Scholar] [CrossRef] [Green Version]
- Tavian, M.; Coulombel, L.; Luton, D.; Clemente, H.S.; Dieterlen-Lièvre, F.; Péault, B. Aorta-Associated CD34+ Hematopoietic Cells in the Early Human Embryo. Blood 1996, 87, 67–72. [Google Scholar] [CrossRef]
- Bertrand, J.Y.; Chi, N.C.; Santoso, B.; Teng, S.; Stainier, D.Y.R.; Traver, D. Haematopoietic Stem Cells Derive Directly from Aortic Endothelium during Development. Nature 2010, 464, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisset, J.-C.; van Cappellen, W.; Andrieu-Soler, C.; Galjart, N.; Dzierzak, E.; Robin, C. In Vivo Imaging of Haematopoietic Cells Emerging from the Mouse Aortic Endothelium. Nature 2010, 464, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Hemmati, H.D.; Wandycz, A.M.; Weissman, I.L. The Purification and Characterization of Fetal Liver Hematopoietic Stem Cells. Proc. Natl. Acad. Sci. USA 1995, 92, 10302–10306. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xu, C.; Asada, N.; Frenette, P.S. The Hematopoietic Stem Cell Niche: From Embryo to Adult. Development 2018, 145, dev139691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares-da-Silva, F.; Peixoto, M.; Cumano, A.; Pinto-do-Ó, P. Crosstalk Between the Hepatic and Hematopoietic Systems During Embryonic Development. Front. Cell Dev. Biol. 2020, 8, 612. [Google Scholar] [CrossRef]
- Hirsch, E.; Iglesias, A.; Potocnik, A.J.; Hartmann, U.; Fässler, R. Impaired Migration but Not Differentiation of Haematopoietic Stem Cells in the Absence of Β1 Integrins. Nature 1996, 380, 171–175. [Google Scholar] [CrossRef]
- Roy, V.; Verfaillie, C.M. Expression and Function of Cell Adhesion Molecules on Fetal Liver, Cord Blood and Bone Marrow Hematopoietic Progenitors. Exp. Hematol. 1999, 27, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Y.; Miyanishi, M.; Wang, S.K.; Yamazaki, S.; Sinha, R.; Kao, K.S.; Seita, J.; Sahoo, D.; Nakauchi, H.; Weissman, I.L. Hoxb5 Marks Long-Term Haematopoietic Stem Cells and Reveals a Homogenous Perivascular Niche. Nature 2016, 530, 223–227. [Google Scholar] [CrossRef]
- Ara, T.; Tokoyoda, K.; Sugiyama, T.; Egawa, T.; Kawabata, K.; Nagasawa, T. Long-Term Hematopoietic Stem Cells Require Stromal Cell-Derived Factor-1 for Colonizing Bone Marrow during Ontogeny. Immunity 2003, 19, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Ema, H.; Nakauchi, H. Expansion of Hematopoietic Stem Cells in the Developing Liver of a Mouse Embryo. Blood 2000, 95, 2284–2288. [Google Scholar] [CrossRef] [Green Version]
- Borge, O.; Ramsfjell, V.; Veiby, O.; Murphy, M.J.; Lok, S.; Jacobsen, S. Thrombopoietin, but Not Erythropoietin Promotes Viability and Inhibits Apoptosis of Multipotent Murine Hematopoietic Progenitor Cells in Vitro. Blood 1996, 88, 2859–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit-Cocault, L.; Volle-Challier, C.; Fleury, M.; Péault, B.; Souyri, M. Dual Role of Mpl Receptor during the Establishment of Definitive Hematopoiesis. Development 2007, 134, 3031–3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, K.S.; Kulkeaw, K.; Nakanishi, Y.; Sugiyama, D. Expression of Cytokine and Extracellular Matrix MRNAs in Fetal Hepatic Stellate Cells. Genes Cells 2017, 22, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Chou, S.; Flygare, J.; Lodish, H.F. Fetal Hepatic Progenitors Support Long-Term Expansion of Hematopoietic Stem Cells. Exp. Hematol. 2013, 41, 479–490.e4. [Google Scholar] [CrossRef]
- Ciriza, J.; Thompson, H.; Petrosian, R.; Manilay, J.O.; García-Ojeda, M.E. The Migration of Hematopoietic Progenitors from the Fetal Liver to the Fetal Bone Marrow: Lessons Learned and Possible Clinical Applications. Exp. Hematol. 2013, 41, 411–423. [Google Scholar] [CrossRef]
- Wittig, O.; Paez-Cortez, J.; Cardier, J.E. Liver Sinusoidal Endothelial Cells Promote B Lymphopoiesis from Primitive Hematopoietic Cells. Stem Cells Dev. 2010, 19, 341–350. [Google Scholar] [CrossRef]
- Li, Y.; Lu, L.; Cai, X. Liver Regeneration and Cell Transplantation for End-Stage Liver Disease. Biomolecules 2021, 11, 1907. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.A.; Glorioso, J.M.; Nyberg, S.L. Liver Regeneration. Transl. Res. 2014, 163, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalopoulos, G.K. Liver Regeneration. J. Cell. Physiol. 2007, 213, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Ma, K.; Zhang, J.; Qatanani, M.; Cuvillier, J.; Liu, J.; Dong, B.; Huang, X.; Moore, D.D. Nuclear Receptor-Dependent Bile Acid Signaling is Required for Normal Liver Regeneration. Science 2006, 312, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.H.; Kalinichenko, V.V.; Holterman, A.-X.L.; Wang, X. Transcription Factors in Liver Development, Differentiation, and Regeneration. Hepatology 2003, 38, ajhep09034. [Google Scholar] [CrossRef] [PubMed]
- Kiseleva, Y.V.; Antonyan, S.Z.; Zharikova, T.S.; Tupikin, K.A.; Kalinin, D.V.; Zharikov, Y.O. Molecular Pathways of Liver Regeneration: A Comprehensive Review. World J. Hepatol. 2021, 13, 270–290. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Cho, G.-S.; Han, C.; Park, D.-H.; Park, H.-K.; Woo, D.-H.; Kim, J.-H. Current Understanding of Stem Cell and Secretome Therapies in Liver Diseases. Tissue Eng. Regen. Med. 2017, 14, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Steiling, H.; Wüstefeld, T.; Bugnon, P.; Brauchle, M.; Fässler, R.; Teupser, D.; Thiery, J.; Gordon, J.I.; Trautwein, C.; Werner, S. Fibroblast Growth Factor Receptor Signalling is Crucial for Liver Homeostasis and Regeneration. Oncogene 2003, 22, 4380–4388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, E.; Sakisaka, S.; Matsuo, K.; Tanikawa, K.; Sata, M. Expression and Role of Vascular Endothelial Growth Factor in Liver Regeneration after Partial Hepatectomy in Rats. J. Histochem. Cytochem. 2001, 49, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desbois-Mouthon, C.; Wendum, D.; Cadoret, A.; Rey, C.; Leneuve, P.; Blaise, A.; Housset, C.; Tronche, F.; le Bouc, Y.; Holzenberger, M. Hepatocyte Proliferation during Liver Regeneration is Impaired in Mice with Liver-specific IGF-1R Knockout. FASEB J. 2006, 20, 773–775. [Google Scholar] [CrossRef] [Green Version]
- Nelsen, C.J.; Rickheim, D.G.; Timchenko, N.A.; Stanley, M.W.; Albrecht, J.H. Transient Expression of Cyclin D1 is Sufficient to Promote Hepatocyte Replication and Liver Growth in Vivo. Cancer Res. 2001, 61, 8564–8568. [Google Scholar] [PubMed]
- Tan, X.; Behari, J.; Cieply, B.; Michalopoulos, G.K.; Monga, S.P.S. Conditional Deletion of β-Catenin Reveals Its Role in Liver Growth and Regeneration. Gastroenterology 2006, 131, 1561–1572. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.-M.; Hou, L.-H.; Dou, G.-R.; Wang, Y.-C.; Hu, X.-B.; He, F.; Feng, F.; Zhang, H.-W.; Liang, Y.-M.; et al. Disruption of the Transcription Factor Recombination Signal-Binding Protein-Jκ (RBP-J) Leads to Veno-Occlusive Disease and Interfered Liver Regeneration in Mice. Hepatology 2009, 49, 268–277. [Google Scholar] [CrossRef]
- Karkampouna, S.; ten Dijke, P.; Dooley, S.; Kruithof-de Julio, M. TGFβ Signaling in Liver Regeneration. Curr. Pharm. Des. 2012, 18, 4103–4113. [Google Scholar] [CrossRef]
- Huck, I.; Gunewardena, S.; Espanol-Suner, R.; Willenbring, H.; Apte, U. Hepatocyte Nuclear Factor 4 Alpha Activation is Essential for Termination of Liver Regeneration in Mice. Hepatology 2019, 70, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.-P.; Jiang, Y.-Z.; Sun, L.-Y.; Zhu, Z.-J. Therapeutic Effect and Safety of Stem Cell Therapy for Chronic Liver Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Stem Cell Res. Ther. 2020, 11, 419. [Google Scholar] [CrossRef]
- Moya, I.M.; Halder, G. Hippo–YAP/TAZ Signalling in Organ Regeneration and Regenerative Medicine. Nat. Rev. Mol. Cell Biol. 2019, 20, 211–226. [Google Scholar] [CrossRef] [PubMed]
- De Silvestro, G.; Vicarioto, M.; Donadel, C.; Menegazzo, M.; Marson, P.; Corsini, A. Mobilization of Peripheral Blood Hematopoietic Stem Cells Following Liver Resection Surgery. Hepatogastroenterology 2004, 51, 805–810. [Google Scholar] [PubMed]
- Fujii, H.; Hirose, T.; Oe, S.; Yasuchika, K.; Azuma, H.; Fujikawa, T.; Nagao, M.; Yamaoka, Y. Contribution of Bone Marrow Cells to Liver Regeneration after Partial Hepatectomy in Mice. J. Hepatol. 2002, 36, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Kollet, O.; Shivtiel, S.; Chen, Y.-Q.; Suriawinata, J.; Thung, S.N.; Dabeva, M.D.; Kahn, J.; Spiegel, A.; Dar, A.; Samira, S.; et al. HGF, SDF-1, and MMP-9 are Involved in Stress-Induced Human CD34+ Stem Cell Recruitment to the Liver. J. Clin. Investig. 2003, 112, 160–169. [Google Scholar] [CrossRef]
- Crosby, H.A.; Lalor, P.F.; Ross, E.; Newsome, P.N.; Adams, D.H. Adhesion of Human Haematopoietic (CD34+) Stem Cells to Human Liver Compartments is Integrin and CD44 Dependent and Modulated by CXCR3 and CXCR4. J. Hepatol. 2009, 51, 734–749. [Google Scholar] [CrossRef]
- King, A.; Houlihan, D.D.; Kavanagh, D.; Haldar, D.; Luu, N.; Owen, A.; Suresh, S.; Than, N.N.; Reynolds, G.; Penny, J.; et al. Sphingosine-1-Phosphate Prevents Egress of Hematopoietic Stem Cells from Liver to Reduce Fibrosis. Gastroenterology 2017, 153, 233–248.e16. [Google Scholar] [CrossRef] [Green Version]
- Forbes, S.J.; Newsome, P.N. Liver Regeneration—Mechanisms and Models to Clinical Application. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 473–485. [Google Scholar] [CrossRef]
- Petersen, B.E.; Bowen, W.C.; Patrene, K.D.; Mars, W.M.; Sullivan, A.K.; Murase, N.; Boggs, S.S.; Greenberger, J.S.; Goff, J.P. Bone Marrow as a Potential Source of Hepatic Oval Cells. Science 1999, 284, 1168–1170. [Google Scholar] [CrossRef]
- Lagasse, E.; Connors, H.; Al-Dhalimy, M.; Reitsma, M.; Dohse, M.; Osborne, L.; Wang, X.; Finegold, M.; Weissman, I.L.; Grompe, M. Purified Hematopoietic Stem Cells Can Differentiate into Hepatocytes In Vivo. Nat. Med. 2000, 6, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Alison, M.R.; Poulsom, R.; Jeffery, R.; Dhillon, A.P.; Quaglia, A.; Jacob, J.; Novelli, M.; Prentice, G.; Williamson, J.; Wright, N.A. Hepatocytes from Non-Hepatic Adult Stem Cells. Nature 2000, 406, 257. [Google Scholar] [CrossRef] [PubMed]
- Theise, N.D.; Nimmakayalu, M.; Gardner, R.; Illei, P.B.; Morgan, G.; Teperman, L.; Henegariu, O.; Krause, D.S. Liver from Bone Marrow in Humans. Hepatology 2000, 32, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagers, A.J.; Sherwood, R.I.; Christensen, J.L.; Weissman, I.L. Little Evidence for Developmental Plasticity of Adult Hematopoietic Stem Cells. Science 2002, 297, 2256–2259. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulos, G.; Wang, P.-R.; Russell, D.W. Transplanted Bone Marrow Regenerates Liver by Cell Fusion. Nature 2003, 422, 901–904. [Google Scholar] [CrossRef]
- Wang, X.; Willenbring, H.; Akkari, Y.; Torimaru, Y.; Foster, M.; Al-Dhalimy, M.; Lagasse, E.; Finegold, M.; Olson, S.; Grompe, M. Cell Fusion is the Principal Source of Bone-Marrow-Derived Hepatocytes. Nature 2003, 422, 897–901. [Google Scholar] [CrossRef] [PubMed]
- Kashofer, K.; Siapati, E.K.; Bonnet, D. In Vivo Formation of Unstable Heterokaryons after Liver Damage and Hematopoietic Stem Cell/Progenitor Transplantation. Stem Cells 2006, 24, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Jaiswal, A.K.; Mukhopadhyay, A. Hepatocyte Nuclear Factor-4α Induces Transdifferentiation of Hematopoietic Cells into Hepatocytes. J. Biol. Chem. 2010, 285, 4725–4731. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.-Y.; Collector, M.I.; Baylin, S.B.; Diehl, A.M.; Sharkis, S.J. Hematopoietic Stem Cells Convert into Liver Cells within Days without Fusion. Nat. Cell Biol. 2004, 6, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Muraca, M.; Ferraresso, C.; Vilei, M.T.; Granato, A.; Quarta, M.; Cozzi, E.; Rugge, M.; Pauwelyn, K.A.; Caruso, M.; Avital, I.; et al. Liver Repopulation with Bone Marrow Derived Cells Improves the Metabolic Disorder in the Gunn Rat. Gut 2007, 56, 1725–1735. [Google Scholar] [CrossRef] [PubMed]
- Newsome, P.N.; Johannessen, I.; Boyle, S.; Dalakas, E.; Mcaulay, K.A.; Samuel, K.; Rae, F.; Forrester, L.; Turner, M.L.; Hayes, P.C.; et al. Human Cord Blood-Derived Cells Can Differentiate into Hepatocytes in the Mouse Liver with No Evidence of Cellular Fusion. Gastroenterology 2003, 124, 1891–1900. [Google Scholar] [CrossRef]
- Tang, X.-P. Differentiation of Human Umbilical Cord Blood Stem Cells into Hepatocytes in Vivo and in Vitro. World J. Gastroenterol. 2006, 12, 4014. [Google Scholar] [CrossRef]
- Pedone, E.; Olteanu, V.-A.; Marucci, L.; Muñoz-Martin, M.I.; Youssef, S.A.; de Bruin, A.; Cosma, M.P. Modeling Dynamics and Function of Bone Marrow Cells in Mouse Liver Regeneration. Cell Rep. 2017, 18, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, D.; Estrov, Z.; Raj, S.; Willerson, J.T.; Yeh, E.T.H. Both Cell Fusion and Transdifferentiation Account for the Transformation of Human Peripheral Blood CD34-Positive Cells into Cardiomyocytes In Vivo. Circulation 2004, 110, 3803–3807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida-Porada, G.; Porada, C.D.; Chamberlain, J.; Torabi, A.; Zanjani, E.D. Formation of Human Hepatocytes by Human Hematopoietic Stem Cells in Sheep. Blood 2004, 104, 2582–2590. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, Y.N.; Kim, J.S.; Park, M.-S.; Paik, Y.H.; Seok, J.-Y.; Chung, Y.E.; Kim, H.O.; Kim, K.S.; Ahn, S.H.; et al. Autologous Bone Marrow Infusion Activates the Progenitor Cell Compartment in Patients with Advanced Liver Cirrhosis. Cell Transplant. 2010, 19, 1237–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, B. Mouse A6–Positive Hepatic Oval Cells Also Express Several Hematopoietic Stem Cell Markers. Hepatology 2003, 37, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Weimar, I.S.; Miranda, N.; Muller, E.J.; Hekman, A.; Kerst, J.M.; de Gast, G.C.; Gerritsen, W.R. Hepatocyte Growth Factor/Scatter Factor (HGF/SF) is Produced by Human Bone Marrow Stromal Cells and Promotes Proliferation, Adhesion and Survival of Human Hematopoietic Progenitor Cells (CD34+). Exp. Hematol. 1998, 26, 885–894. [Google Scholar]
- Zachman, D.K.; Goldman, D.C.; Hamlin, K.L.; Guha, C.; Fleming, W.H. Role of Hepatocyte Growth Factor in Endothelial-Dependent Hematopoietic Stem Cell Regeneration. Blood 2014, 124, 4369. [Google Scholar] [CrossRef]
- Goff, J.P.; Shields, D.S.; Petersen, B.E.; Zajac, V.F.; Michalopoulos, G.K.; Greenberger, J.S. Synergistic Effects of Hepatocyte Growth Factor on Human Cord Blood CD34 + Progenitor Cells are the Result of C-met Receptor Expression. Stem Cells 1996, 14, 592–602. [Google Scholar] [CrossRef]
- Baccin, C.; Al-Sabah, J.; Velten, L.; Helbling, P.M.; Grünschläger, F.; Hernández-Malmierca, P.; Nombela-Arrieta, C.; Steinmetz, L.M.; Trumpp, A.; Haas, S. Combined Single-Cell and Spatial Transcriptomics Reveal the Molecular, Cellular and Spatial Bone Marrow Niche Organization. Nat. Cell Biol. 2020, 22, 38–48. [Google Scholar] [CrossRef]
- Fujii, K.; Ishimaru, F.; Kozuka, T.; Matsuo, K.; Nakase, K.; Kataoka, I.; Tabayashi, T.; Shinagawa, K.; Ikeda, K.; Harada, M.; et al. Elevation of Serum Hepatocyte Growth Factor during Granulocyte Colony-Stimulating Factor-Induced Peripheral Blood Stem Cell Mobilization. Br. J. Haematol. 2004, 124, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Carstanjen, D. Interleukin-6 is a Major Effector Molecule of Short-Term G-CSF Treatment Inducing Bone Metabolism and an Acute-Phase Response. Exp. Hematol. 2001, 29, 812–821. [Google Scholar] [CrossRef]
- Garg, V.; Garg, H.; Khan, A.; Trehanpati, N.; Kumar, A.; Sharma, B.C.; Sakhuja, P.; Sarin, S.K. Granulocyte Colony–Stimulating Factor Mobilizes CD34+ Cells and Improves Survival of Patients with Acute-on-Chronic Liver Failure. Gastroenterology 2012, 142, 505–512.e1. [Google Scholar] [CrossRef] [PubMed]
- Ieda, Y.; Fujita, J.; Ieda, M.; Yagi, T.; Kawada, H.; Ando, K.; Fukuda, K. G-CSF and HGF: Combination of Vasculogenesis and Angiogenesis Synergistically Improves Recovery in Murine Hind Limb Ischemia. J. Mol. Cell. Cardiol. 2007, 42, 540–548. [Google Scholar] [CrossRef]
- Piscaglia, A.C.; Shupe, T.D.; Oh, S.; Gasbarrini, A.; Petersen, B.E. Granulocyte–Colony Stimulating Factor Promotes Liver Repair and Induces Oval Cell Migration and Proliferation in Rats. Gastroenterology 2007, 133, 619–631. [Google Scholar] [CrossRef] [Green Version]
- Tsolaki, E.; Athanasiou, E.; Gounari, E.; Zogas, N.; Siotou, E.; Yiangou, M.; Anagnostopoulos, A.; Yannaki, E. Hematopoietic Stem Cells and Liver Regeneration: Differentially Acting Hematopoietic Stem Cell Mobilization Agents Reverse Induced Chronic Liver Injury. Blood Cells Mol. Dis. 2014, 53, 124–132. [Google Scholar] [CrossRef]
- Zekri, A.-R.N.; Salama, H.; Medhat, E.; Musa, S.; Abdel-Haleem, H.; Ahmed, O.S.; Khedr, H.A.H.; Lotfy, M.M.; Zachariah, K.S.; Bahnassy, A.A. The Impact of Repeated Autologous Infusion of Haematopoietic Stem Cells in Patients with Liver Insufficiency. Stem Cell Res. Ther. 2015, 6, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terai, S.; Ishikawa, T.; Omori, K.; Aoyama, K.; Marumoto, Y.; Urata, Y.; Yokoyama, Y.; Uchida, K.; Yamasaki, T.; Fujii, Y.; et al. Improved Liver Function in Patients with Liver Cirrhosis after Autologous Bone Marrow Cell Infusion Therapy. Stem Cells 2006, 24, 2292–2298. [Google Scholar] [CrossRef]
- Yannaki, E.; Anagnostopoulos, A.; Kapetanos, D.; Xagorari, A.; Iordanidis, F.; Batsis, I.; Kaloyannidis, P.; Athanasiou, E.; Dourvas, G.; Kitis, G.; et al. Lasting Amelioration in the Clinical Course of Decompensated Alcoholic Cirrhosis with Boost Infusions of Mobilized Peripheral Blood Stem Cells. Exp. Hematol. 2006, 34, 1583–1587. [Google Scholar] [CrossRef]
- Gaia, S.; Smedile, A.; Omedè, P.; Olivero, A.; Sanavio, F.; Balzola, F.; Ottobrelli, A.; Abate, M.L.; Marzano, A.; Rizzetto, M.; et al. Feasibility and Safety of G-CSF Administration to Induce Bone Marrow-Derived Cells Mobilization in Patients with End Stage Liver Disease. J. Hepatol. 2006, 45, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.Y.; Levičar, N.; Pai, M.; Bachellier, P.; Dimarakis, I.; Al-Allaf, F.; M’Hamdi, H.; Thalji, T.; Welsh, J.P.; Marley, S.B.; et al. Characterization and Clinical Application of Human CD34+ Stem/Progenitor Cell Populations Mobilized into the Blood by Granulocyte Colony-Stimulating Factor. Stem Cells 2006, 24, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Mohamadnejad, M. Phase 1 Human Trial of Autologous Bone Marrow-Hematopoietic Stem Cell Transplantation in Patients with Decompensated Cirrhosis. World J. Gastroenterol. 2007, 13, 3359. [Google Scholar] [CrossRef] [PubMed]
- Lyra, A.C. Feasibility and Safety of Autologous Bone Marrow Mononuclear Cell Transplantation in Patients with Advanced Chronic Liver Disease. World J. Gastroenterol. 2007, 13, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Han, Y.; Wang, J.; Liu, J.; Hong, L.; Fan, D. Peripheral Blood Monocytes from Patients with HBV Related Decompensated Liver Cirrhosis Can Differentiate into Functional Hepatocytes. Am. J. Hematol. 2007, 82, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Levičar, N.; Pai, M.; Habib, N.A.; Tait, P.; Jiao, L.R.; Marley, S.B.; Davis, J.; Dazzi, F.; Smadja, C.; Jensen, S.L.; et al. Long-Term Clinical Results of Autologous Infusion of Mobilized Adult Bone Marrow Derived CD34+ Cells in Patients with Chronic Liver Disease. Cell Prolif. 2007, 41, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Parveen, N.; Mahaboob, V.S.; Rajendraprasad, A.; Ravindraprakash, H.R.; Venkateswarlu, J.; Rao, S.G.A.; Narusu, M.L.; Khaja, M.N.; Pramila, R.; et al. Safety and Efficacy of Autologous Bone Marrow Stem Cell Transplantation Through Hepatic Artery for the Treatment of Chronic Liver Failure: A Preliminary Study. Transplant. Proc. 2008, 40, 1140–1144. [Google Scholar] [CrossRef]
- Han, Y.; Yan, L.; Han, G.; Zhou, X.; Hong, L.; Yin, Z.; Zhang, X.; Wang, S.; Wang, J.; Sun, A.; et al. Controlled Trials in Hepatitis B Virus-Related Decompensate Liver Cirrhosis: Peripheral Blood Monocyte Transplant versus Granulocyte–Colony-Stimulating Factor Mobilization Therapy. Cytotherapy 2008, 10, 390–396. [Google Scholar] [CrossRef]
- Pai, M.; Zacharoulis, D.; Milicevic, M.N.; Helmy, S.; Jiao, L.R.; Levičar, N.; Tait, P.; Scott, M.; Marley, S.B.; Jestice, K.; et al. Autologous Infusion of Expanded Mobilized Adult Bone Marrow-Derived CD34+ Cells into Patients with Alcoholic Liver Cirrhosis. Am. J. Gastroenterol. 2008, 103, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Salama, H. Autologous CD34 + and CD133 + Stem Cells Transplantation in Patients with End Stage Liver Disease. World J. Gastroenterol. 2010, 16, 5297. [Google Scholar] [CrossRef] [PubMed]
- Salama, H.; Zekri, A.-R.; Zern, M.; Bahnassy, A.; Loutfy, S.; Shalaby, S.; Vigen, C.; Burke, W.; Mostafa, M.; Medhat, E.; et al. Autologous Hematopoietic Stem Cell Transplantation in 48 Patients with End-Stage Chronic Liver Diseases. Cell Transplant. 2010, 19, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Lyra, A.C.; Soares, M.B.P.; da Silva, L.F.M.; Braga, E.L.; Oliveira, S.A.; Fortes, M.F.; Silva, A.G.P.; Brustolim, D.; Genser, B.; dos Santos, R.R.; et al. Infusion of Autologous Bone Marrow Mononuclear Cells through Hepatic Artery Results in a Short-Term Improvement of Liver Function in Patients with Chronic Liver Disease: A Pilot Randomized Controlled Study. Eur. J. Gastroenterol. Hepatol. 2010, 22, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Okumoto, K.; Haga, H.; Nishise, Y.; Ishii, R.; Sato, C.; Watanabe, H.; Okada, A.; Ikeda, M.; Togashi, H.; et al. Potential Therapeutic Application of Intravenous Autologous Bone Marrow Infusion in Patients with Alcoholic Liver Cirrhosis. Stem Cells Dev. 2011, 20, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Spahr, L.; Chalandon, Y.; Terraz, S.; Kindler, V.; Rubbia-Brandt, L.; Frossard, J.-L.; Breguet, R.; Lanthier, N.; Farina, A.; Passweg, J.; et al. Autologous Bone Marrow Mononuclear Cell Transplantation in Patients with Decompensated Alcoholic Liver Disease: A Randomized Controlled Trial. PLoS ONE 2013, 8, e53719. [Google Scholar] [CrossRef]
- Bai, Y.-Q. Outcomes of Autologous Bone Marrow Mononuclear Cell Transplantation in Decompensated Liver Cirrhosis. World J. Gastroenterol. 2014, 20, 8660. [Google Scholar] [CrossRef]
- Liu, L.; Yan, Y.; Zhou, J.; Huang, L.W.; He, C.P.; Ling, K.; Zhou, H.C.; Wen, Q.M.; Wang, X.M. Curative Effect of Combined Lamivudine, Adefovir Dipivoxil, and Stem Cell Transplantation on Decompensated Hepatitis B Cirrhosis. Genet. Mol. Res. 2014, 13, 9336–9342. [Google Scholar] [CrossRef]
- Andreone, P.; Catani, L.; Margini, C.; Brodosi, L.; Lorenzini, S.; Sollazzo, D.; Nicolini, B.; Giordano, R.; Montemurro, T.; Rizzi, S.; et al. Reinfusion of Highly Purified CD133+ Bone Marrow-Derived Stem/Progenitor Cells in Patients with End-Stage Liver Disease: A Phase I Clinical Trial. Dig. Liver Dis. 2015, 47, 1059–1066. [Google Scholar] [CrossRef]
- Sharma, M.; Rao, P.N.; Sasikala, M.; Kuncharam, M.R.; Reddy, C.; Gokak, V.; Raju, B.; Singh, J.R.; Nag, P.; Reddy, D.N. Autologous Mobilized Peripheral Blood CD34 + Cell Infusion in Non-Viral Decompensated Liver Cirrhosis. World J. Gastroenterol. 2015, 21, 7264–7271. [Google Scholar] [CrossRef]
- Deng, Q.; Cai, T.; Zhang, S.; Hu, A.; Zhang, X.; Wang, Y.; Huang, J. Autologous Peripheral Blood Stem Cell Transplantation Improves Portal Hemodynamics in Patients with Hepatitis B Virus-Related Decompensated Cirrhosis. Hepat. Mon. 2015, 15, e32498. [Google Scholar] [CrossRef] [Green Version]
- Al Tayeb, H.; El Dorry, A.; Amer, N.; Mowafy, N.; Zimaity, M.; Bayoumy, E.; Saleh, A.S.A. Autologous Stem Cells Transplantation in Egyptian Patients with Liver Cirrhosis on Top of Hepatitis C Virus. Int. J. Stem Cells 2015, 8, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Mohamadnejad, M.; Vosough, M.; Moossavi, S.; Nikfam, S.; Mardpour, S.; Akhlaghpoor, S.; Ashrafi, M.; Azimian, V.; Jarughi, N.; Hosseini, S.-E.; et al. Intraportal Infusion of Bone Marrow Mononuclear or CD133+ Cells in Patients with Decompensated Cirrhosis: A Double-Blind Randomized Controlled Trial. Stem Cells Transl. Med. 2016, 5, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.J.; Yoon, J.-H.; Kim, W.; Lee, J.M.; Bin Lee, Y.; Cho, Y.; Lee, D.H.; Lee, M.; Yoo, J.-J.; Cho, E.J.; et al. Ultrasound-Guided Percutaneous Portal Transplantation of Peripheral Blood Monocytes in Patients with Liver Cirrhosis. Korean J. Intern. Med. 2017, 32, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsome, P.N.; Fox, R.; King, A.L.; Barton, D.; Than, N.-N.; Moore, J.; Corbett, C.; Townsend, S.; Thomas, J.; Guo, K.; et al. Granulocyte Colony-Stimulating Factor and Autologous CD133-Positive Stem-Cell Therapy in Liver Cirrhosis (REALISTIC): An Open-Label, Randomised, Controlled Phase 2 Trial. Lancet Gastroenterol. Hepatol. 2018, 3, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Esmaeilzadeh, A.; Ommati, H.; Kooshyar, M.M.; Jarahi, L.; Akhavan Rezayat, K.; Saberi, S.; Vosough, M.; Ghassemi, A. Autologous Bone Marrow Stem Cell Transplantation in Liver Cirrhosis after Correcting Nutritional Anomalies, A Controlled Clinical Study. Cell J. 2019, 21, 268–273. [Google Scholar] [CrossRef]
- Cui, L.N.; Wang, X.F.; Sun, R.Q.; Deng, J.; Gao, Z.J.; Zhou, X.M.; Guo, C.C.; Jia, G.; Shang, Y.L.; Yang, C.M.; et al. Study of the Effects of Long-Term Outcomes of Autologous Peripheral Blood Stem Cell Reinfusion in Patients with Decompensated Cirrhosis. Zhonghua Gan Zang Bing Za Zhi 2022, 30, 279–284. [Google Scholar] [CrossRef]
- Chavez-Tapia, N.C.; Mendiola-Pastrana, I.; Ornelas-Arroyo, V.J.; Noreña-Herrera, C.; Vidaña-Perez, D.; Delgado-Sanchez, G.; Uribe, M.; Barrientos-Gutierrez, T. Granulocyte-Colony Stimulating Factor for Acute-on-Chronic Liver Failure: Systematic Review and Meta-Analysis. Ann. Hepatol. 2015, 14, 631–641. [Google Scholar] [CrossRef]
- Zhu, C.-H.; Zhang, D.-H.; Zhu, C.-W.; Xu, J.; Guo, C.-L.; Wu, X.-G.; Cao, Q.-L.; Di, G.-H. Adult Stem Cell Transplantation Combined with Conventional Therapy for the Treatment of End-Stage Liver Disease: A Systematic Review and Meta-Analysis. Stem Cell Res. Ther. 2021, 12, 558. [Google Scholar] [CrossRef]
- Wu, C.-X.; Wang, D.; Cai, Y.; Luo, A.-R.; Sun, H. Effect of Autologous Bone Marrow Stem Cell Therapy in Patients with Liver Cirrhosis: A Meta-Analysis. J. Clin. Transl. Hepatol. 2019, 7, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Yannaki, E.; Athanasiou, E.; Xagorari, A.; Constantinou, V.; Batsis, I.; Kaloyannidis, P.; Proya, E.; Anagnostopoulos, A.; Fassas, A. G-CSF–Primed Hematopoietic Stem Cells or G-CSF per Se Accelerate Recovery and Improve Survival after Liver Injury, Predominantly by Promoting Endogenous Repair Programs. Exp. Hematol. 2005, 33, 108–119. [Google Scholar] [CrossRef]
- Hu, X.-M.; Zhang, Q.; Zhou, R.-X.; Wu, Y.-L.; Li, Z.-X.; Zhang, D.-Y.; Yang, Y.-C.; Yang, R.-H.; Hu, Y.-J.; Xiong, K. Programmed Cell Death in Stem Cell-Based Therapy: Mechanisms and Clinical Applications. World J. Stem Cells 2021, 13, 386–415. [Google Scholar] [CrossRef]
- García Martínez, J.J.; Bendjelid, K. Artificial Liver Support Systems: What is New over the Last Decade? Ann. Intensive Care 2018, 8, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azparren-Angulo, M.; Royo, F.; Gonzalez, E.; Liebana, M.; Brotons, B.; Berganza, J.; Goñi-de-Cerio, F.; Manicardi, N.; Abad-Jordà, L.; Gracia-Sancho, J.; et al. Extracellular Vesicles in Hepatology: Physiological Role, Involvement in Pathogenesis, and Therapeutic Opportunities. Pharmacol. Ther. 2021, 218, 107683. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Tan, W.-F.; Yang, M.-Q.; Li, J.-Y.; Geller, D.A. The Therapeutic Potential of Exosomes Derived from Different Cell Sources in Liver Diseases. Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 322, G397–G404. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | Year | Country | Condition | Design | Patients (Treated/Control) | Cell Type | Cells Injection | Follow-Up Period | Outcome |
|---|---|---|---|---|---|---|---|---|---|
| Terai S. [129] | 2006 | Japan | Liver cirrhosis | Case-control | 9/0 | BM-MNCs (94% CD45+) | 5.2 × 109 | 24 weeks | Improved liver function and ALB levels. A trend towards ascites improvement |
| Yannaki E. [130] | 2006 | Greece | Alcohol-induced liver cirrhosis | Case-control | 2/0 | Autologous BM-HSCs (mobilised CD34+) | 2 × 106/kg | 120 weeks | Improvement of baseline CTP and MELD scores |
| Gaia S. [131] | 2006 | Italy | Severe liver cirrhosis | Case-control | 8/0 | GCSF mobilisation | N/A | 32 weeks | Improvement of baseline CTP and MELD scores in 50% of patients |
| Gordon M. [132] | 2006 | UK | Chronic liver failure | Case-control | 5/0 | Autologous BM-HSCs (mobilised CD34+) | 106–2 × 108 | 60 days | Ν/A |
| Mohamadnejad M. [133] | 2007 | Iran | Decompensated cirrhosis | Case-control | 4/0 | BM-HSCs | 2.5–8 × 106 | 24 weeks | Improvement in ALB levels in 2 patients and MELD scores in 1 patient. |
| Lyra A.C. [134] | 2007 | Brazil | Chronic liver disease | Case-control | 10/0 | BM-MNCs | 108 | 16 weeks | Overall improvement in ALB, TBIL and INR |
| Yan L. [135] | 2007 | China | HBV-related decompensated liver cirrhosis | Case-control | 2/0 | Autologous BM-HSCs (GCSF-mobilised) | 107–108/kg | 18 months | Improvement of baseline CTP score |
| Levicar N. [136] LCER No. 2004/6746 | 2008 | UK | Chronic liver disease | Case-control | 5/0 | Autologous BM-HSCs (GCSF-mobilised CD34+) | 106–2 × 108 | 18 months | Improvement in ALB and AFP levels |
| Khan A.A. [137] | 2008 | India | Liver cirrhosis | Case-control | 4/0 | Autologous BM-HSCs (GCSF-mobilised CD34+) | 0.1 × 108 | 26 weeks | Improvement of baseline CTP and MELD scores |
| Han Y. [138] | 2008 | China | HBV-related decompensated liver cirrhosis | RCT | 20/20 | Autologous BM-MNCs (GCSF-mobilised) | 107–108/kg | 6 months | Improved ALB and CTP score in patients receiving cell transplant |
| Pai M. [139] | 2008 | UK | Severe alcoholic liver cirrhosis | Case-control | 9/0 | Autologous BM-HSCs (GCSF-mobilised CD34+) | 2.3 × 108 | 3 months | Improved TBIL, ALT, AST and CTP score. Some improvement in ascites formation |
| Salama H. [140] | 2010 | Egypt | End-stage liver disease | RCT | 90/50 | Autologous BM-HSCs (mobilised CD34+ and CD133+) | 0.5 × 108 | 24 weeks | Improved liver function and ALB levels |
| Salama H. [141] | 2010 | Egypt | End-stage liver disease | Case-control | 48 | Autologous BM-HSCs (GCSF-mobilised CD34+) | 1 × 109 | 48 weeks | Decrease in ascites; Improvement in ALB, TBIL, INR, ALT |
| Kim J.K. [116] | 2010 | China | Advanced liver cirrhosis | Case-control | 10/0 | Autologous BM-MNCs (80% CD45+) | 0.5–1.5 × 108 | 6 months | Improvement in CPT score and ascites formation |
| Lyra A.C. [142] | 2010 | Brazil | Chronic liver disease | RCT | 15/15 | Autologous BM cells | 3.8 × 108 | 12 months | The MELD score remained stable in treated patients while it increased in the control group. Improvement in ALB and TBIL in the treated group |
| Saito S. [143] | 2011 | Japan | Alcoholic liver cirrhosis | Case-control | 5/5 | Autologous BM-MNCs | 8.0–7.3 × 109 | 24 weeks | Higher ALB and PTA; improved CTP score |
| Garg V. [124] NCT01036932 | 2012 | India | Acute-on-chronic liver failure | RCT | 23/24 | GCSF mobilisation | N/A | 2 months | Improvement in survival, CTP and MELD scores |
| Spahr L. [144] ISRCTN83972743 | 2013 | Switzerland | Decompensated alcoholic liver disease | RCT | 28/30 | Autologous BM-MNCs (GCSF-mobilised) | 0.47 ± 0.15 × 108/kg | 3 months | No improvement in liver function |
| Bai Y.Q. [145] | 2014 | China | HBV-related liver cirrhosis | Case-control | 32/15 | Autologous BM-MNCs | Not reported | 24 months | Improvement in ALB, PTA, fibrinogen, PLT, TBIL and reduction of adverse effects |
| Liu L. [146] | 2014 | China | Hepatitis B and decompensated liver cirrhosis | RCT | 40/37 | Autologous BM-MNCs (GCSF-mobilised) | 3.2 +/−1.6 × 1011 | 4 weeks | Improvement in serum AST, ALT, ALB, and TBIL levels |
| Andreone P. [147] NCT01025622 | 2015 | Italy | End-stage liver disease | Case-control | 12/0 | Autologous BM-HSCs (GCSF-mobilised CD133+) | 5 × 104/kg up to 1 × 106/kg | 12 months | Temporary improvement in MELD score; worsening of CTP score |
| Zekri A.R. [128] NCT01729221 | 2015 | Egypt | HCV-associated liver cirrhosis | RCT | 60/30 | Autologous BM-HSCs (mobilised CD34+) followed by MSC infusion | 0.5 × 108 | 52 weeks | Improvement in baseline CTP in 40% patients. Improvement in ALB, TBIL and INR |
| Sharma M. [148] | 2015 | India | Non-viral decompensated cirrhosis | RCT | 22/23 | Autologous BM-HSCs (GCSF-mobilised CD34+) | N/A | 3 months | Improvement in serum creatinin and MELD scores |
| Deng Q. [149] | 2015 | China | HBV-related decompensated cirrhosis | RCT | 33/35 | Autologous BM-HSCs (GCSF-mobilised CD34+) | 2–4 × 107 | 48 weeks | improvements in liver function (ALB, PTA) and portal vein hemodynamics |
| TayebH. [150] | 2015 | Egypt | HCV-associated liver cirrhosis | RCT | 10/10 | Autologous BM-MNCs (GCSF-mobilised) | 25 × 106–191 × 106 | 3 months | Improvement in ALB levels 1 month post BMT; γ-GT improvement at 3 months; Improved CTP score at 3 months; No statistical improvement in any other liver parameter at 3 months |
| Mohamadnejad M. [151] | 2016 | Iran | Decompensated cirrhosis | RCT | 8 (CD133) 10 (MNC) 9 (CONTROL) | Autologous BM-CD133+ versus BM-MNCs | 2–13 × 108 MNC/2–7 106 CD133+ | 12 months | Improved MELD score in the CD133+ group at 3 mo |
| Yu S.J. [152] NCT01503749 | 2016 | Korea | Decompensated cirrhosis | RCT | 3 (GSCF + CELLS) 3 (GCSF) 3 (CONTROL) | Autologous BM-MNCs (GCSF-mobilised) | 1.67 × 109–2 × 1010 | 6 months | Small improvement in CTP scores at 24 weeks |
| Newsome [153] ISRCTN 91288089 | 2018 | UK | Compensated liver cirrhosis | RCT | 28/26 GCSF/27 control | Autologous BM-HSCs (GCSF-mobilised CD133+) | 0.2 × 10⁶ | 90 days | No improvement in liver dysfunction or fibrosis while adverse events may occur compared with standard care |
| Esmaeilzadeh A. [154] IRCT2014091919217N1 | 2019 | Iran | Decompensated liver cirrhosis | RCT | 10/10 | Autologous BM-MNCs | 8.06 ± 2.5×106 cells/kg | 6 months | Improvement in MELD socre, INR, TBIL, ALB levels after cell transplantation (6 months) |
| Cui L. [155] | 2022 | China | Decompensated liver cirrhosis | 10 yr follow up study | 287/151 | PBSC | Survival was higher in treated group alongside ALB levels, CTP and MELD scores |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siapati, E.K.; Roubelakis, M.G.; Vassilopoulos, G. Liver Regeneration by Hematopoietic Stem Cells: Have We Reached the End of the Road? Cells 2022, 11, 2312. https://doi.org/10.3390/cells11152312
Siapati EK, Roubelakis MG, Vassilopoulos G. Liver Regeneration by Hematopoietic Stem Cells: Have We Reached the End of the Road? Cells. 2022; 11(15):2312. https://doi.org/10.3390/cells11152312
Chicago/Turabian StyleSiapati, Elena Konstantina, Maria G. Roubelakis, and George Vassilopoulos. 2022. "Liver Regeneration by Hematopoietic Stem Cells: Have We Reached the End of the Road?" Cells 11, no. 15: 2312. https://doi.org/10.3390/cells11152312
APA StyleSiapati, E. K., Roubelakis, M. G., & Vassilopoulos, G. (2022). Liver Regeneration by Hematopoietic Stem Cells: Have We Reached the End of the Road? Cells, 11(15), 2312. https://doi.org/10.3390/cells11152312