
Functions of Forkhead Box O on Glucose Metabolism in Abalone Haliotis discus hannai and Its Responses to High Levels of Dietary Lipid


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Gene Cloning, Sequence Analysis, Tissue Distribution, and Function Analysis of foxo
2.2.1. Animals and Sampling
2.2.2. Cloning of foxo and Sequence Analysis
2.2.3. Tissue Distribution
2.2.4. The foxo Interfering
2.2.5. Quantitative Real-Time PCR (qPCR)
2.3. Feeding Trial
2.3.1. Experimental Diets
2.3.2. Feeding and Sampling
2.3.3. Biochemical Parameters
2.3.4. Quantitative Real-Time PCR (qPCR)
2.3.5. Western Blot Analysis
2.4. Statistical Analysis
3. Results
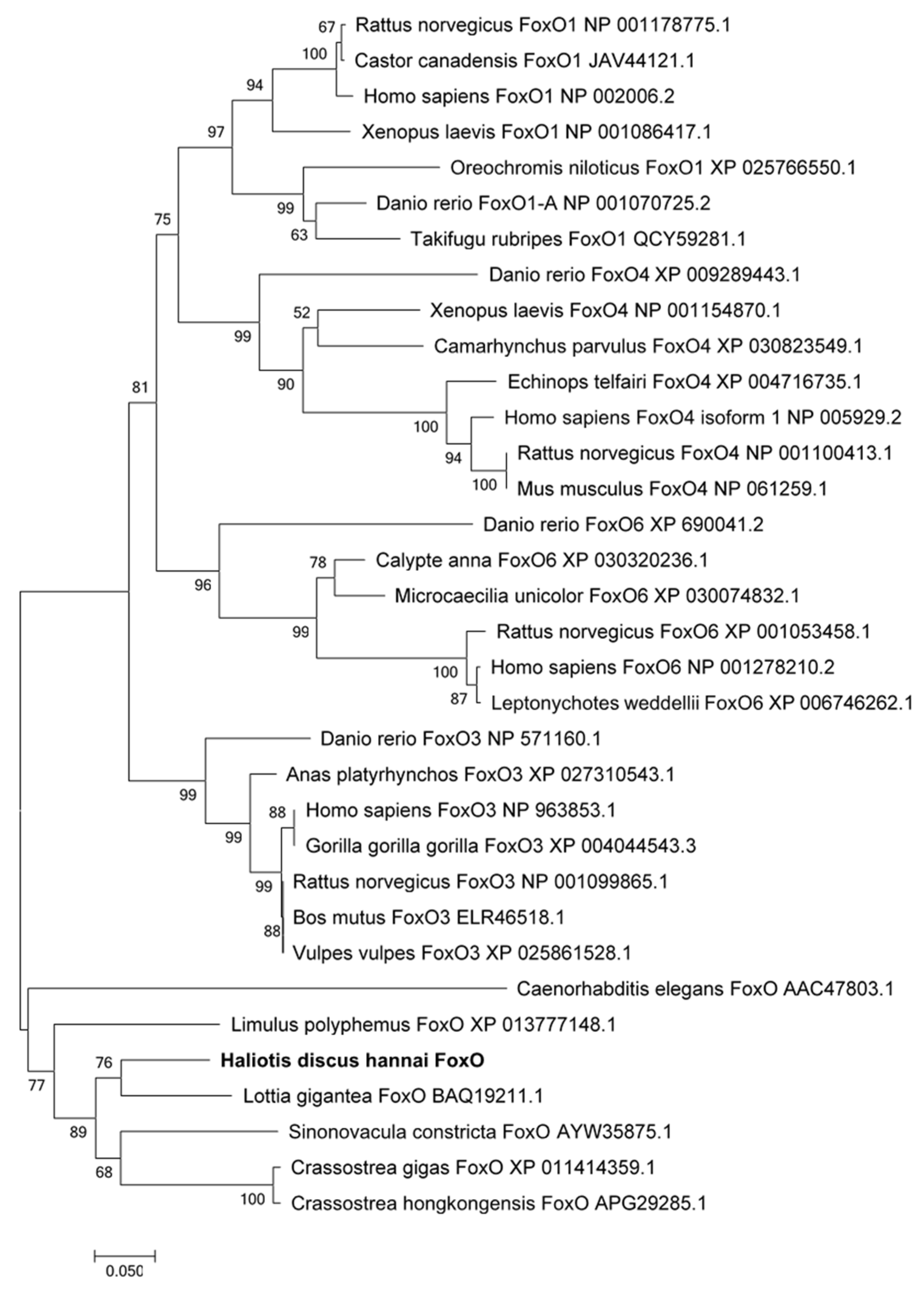
3.1. Identification of foxo in Abalone
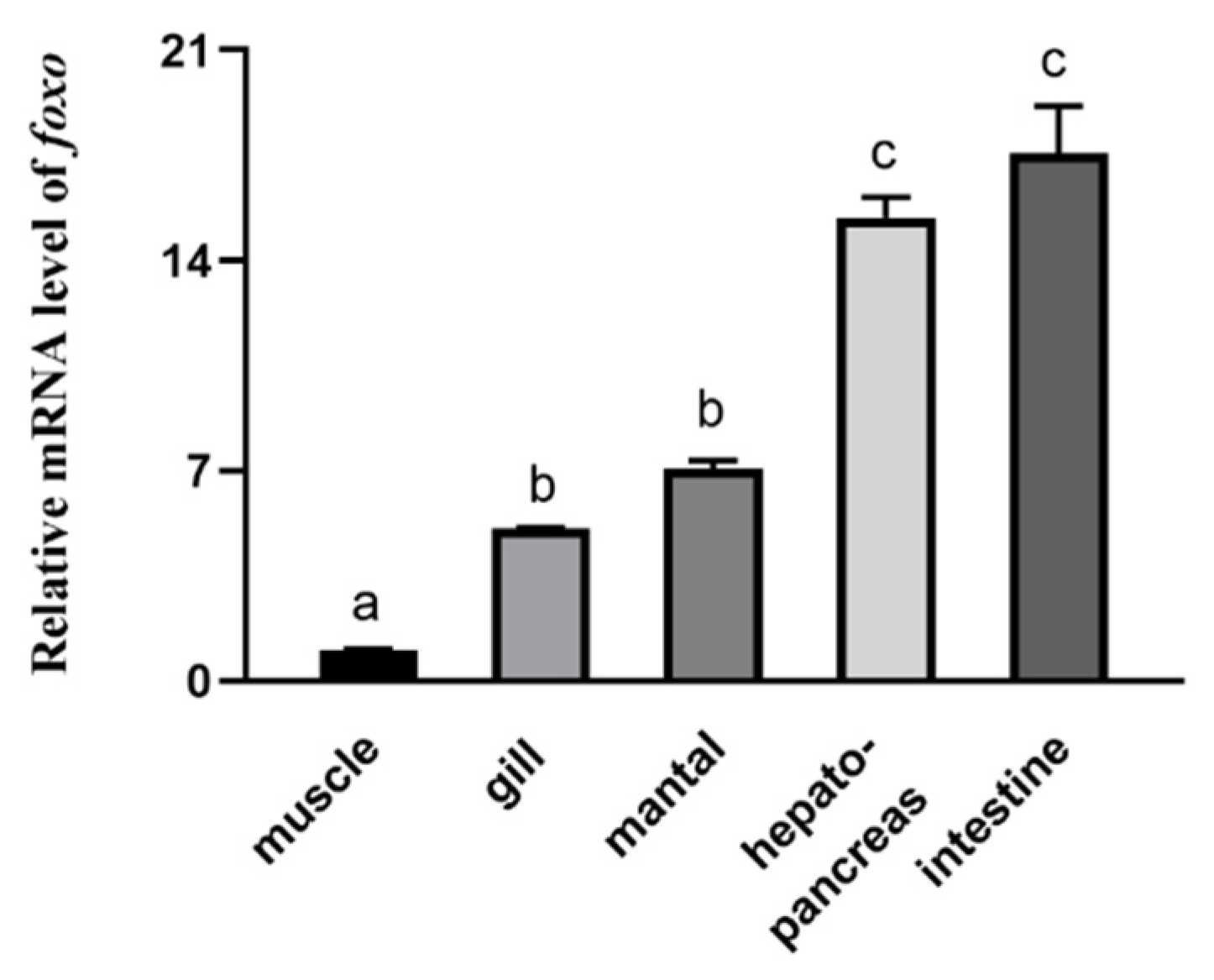
3.2. Tissue Distribution of foxo in Abalone
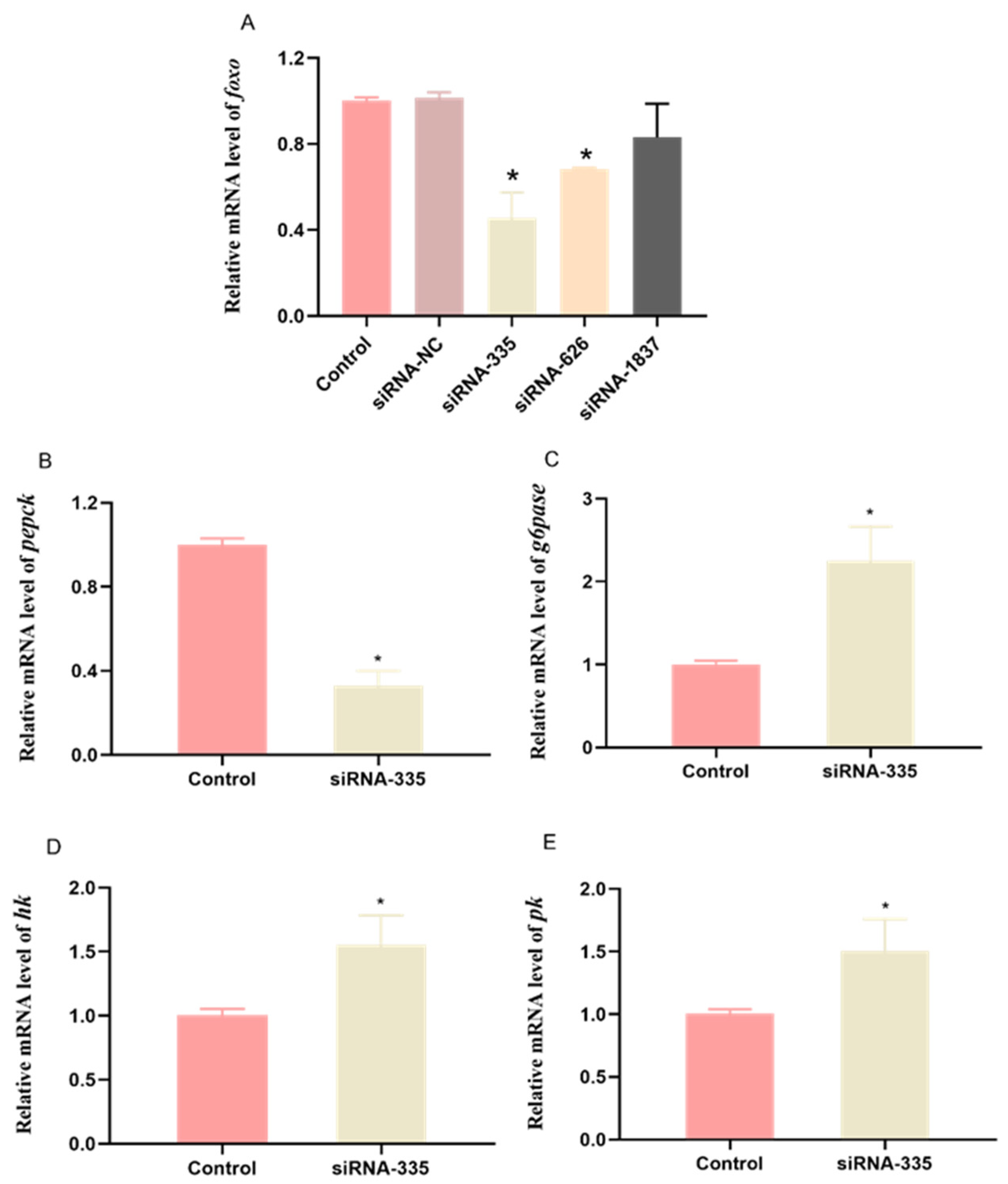
3.3. Knockdown of foxo In Vivo in Abalone
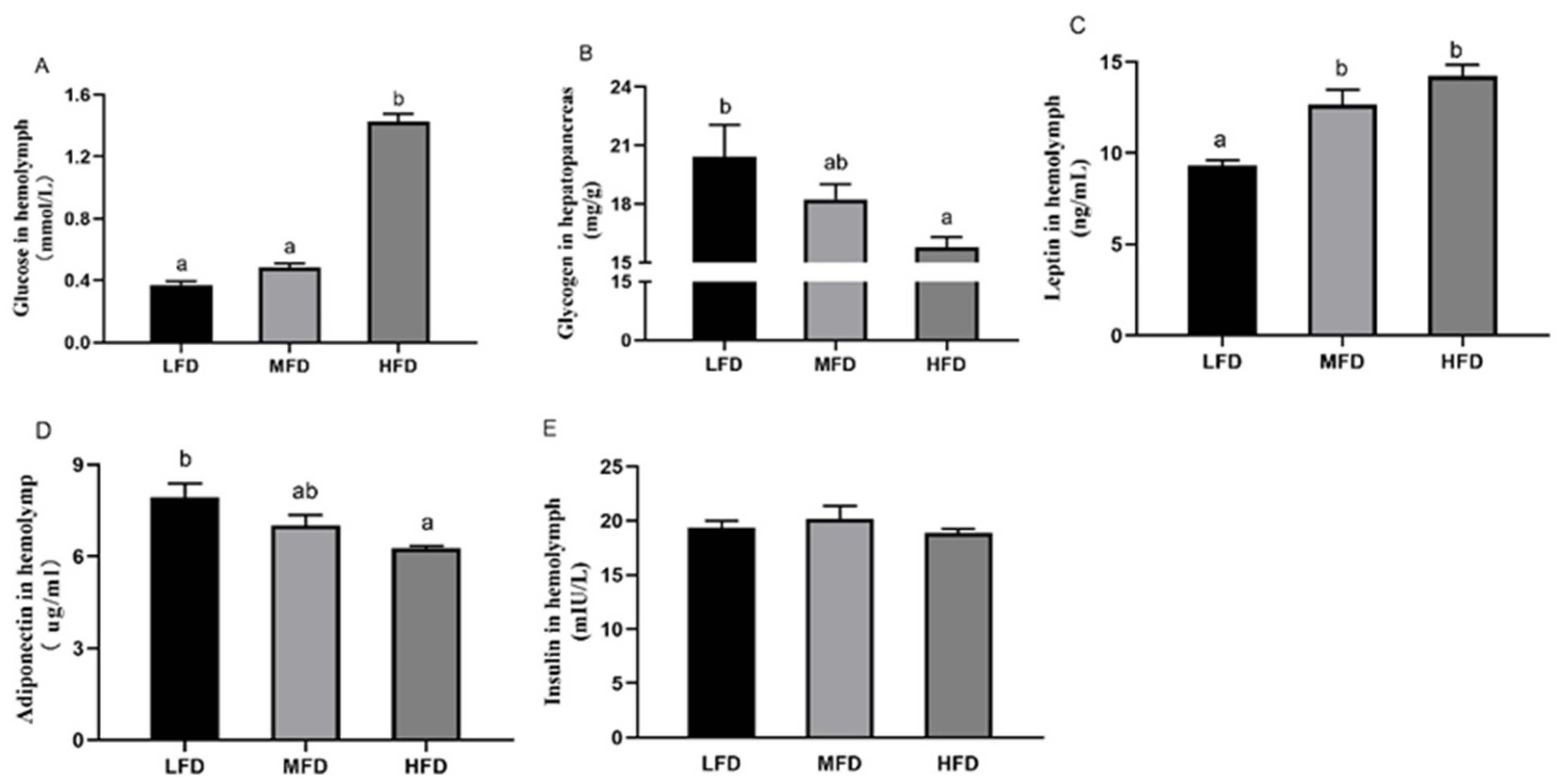
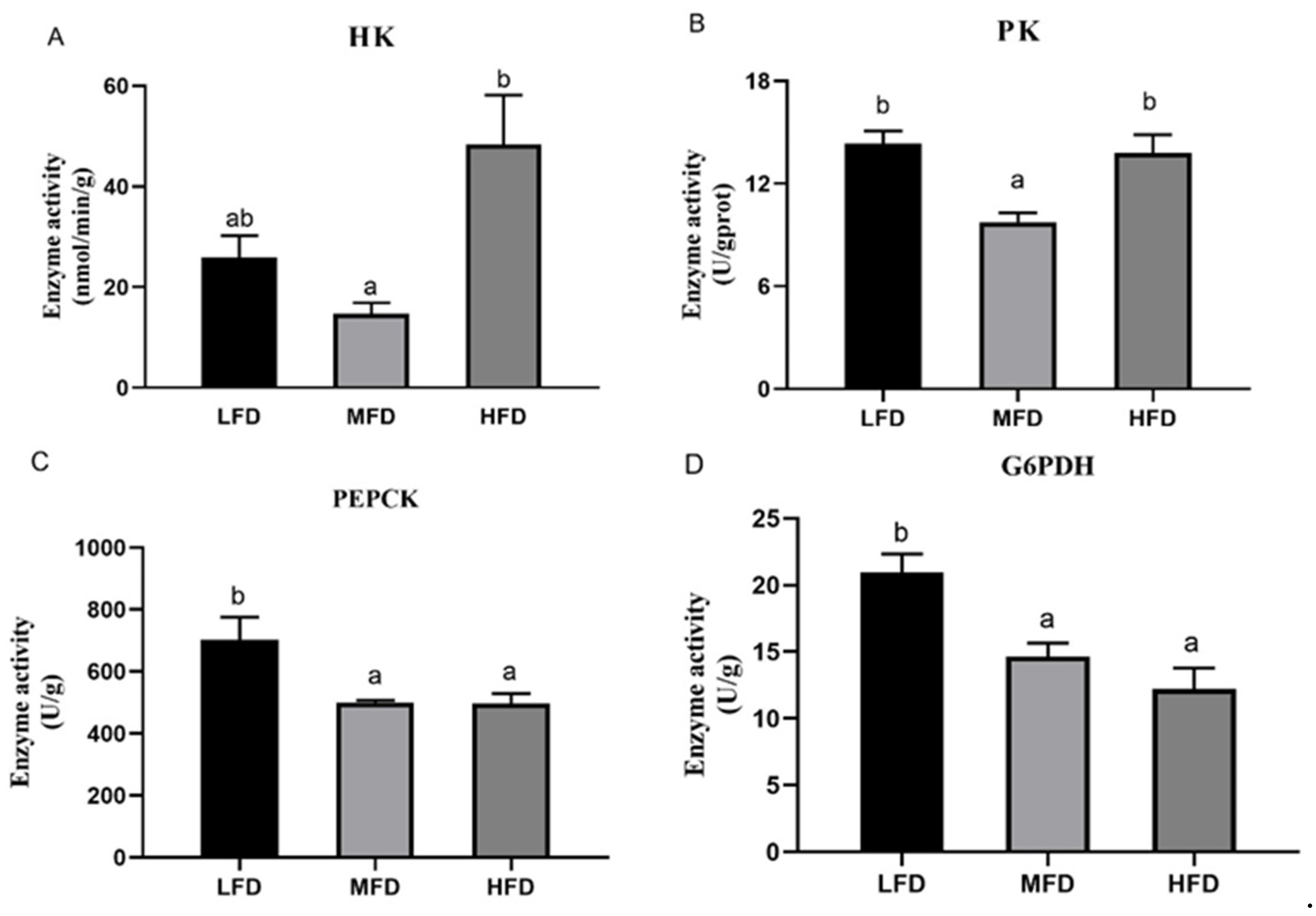
3.4. Hemolymph and Hepatopancreas Parameters after Feeding Trial
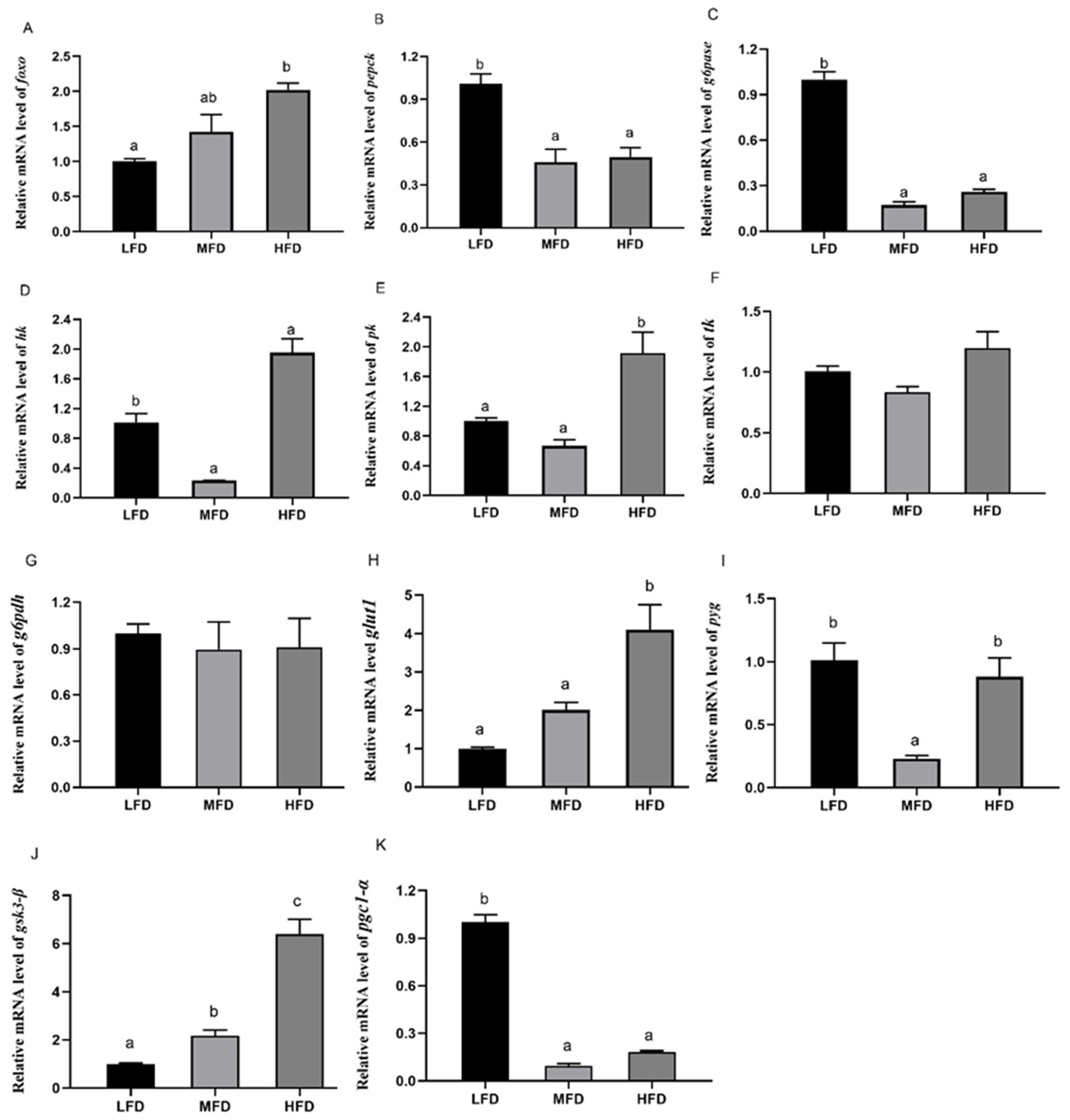
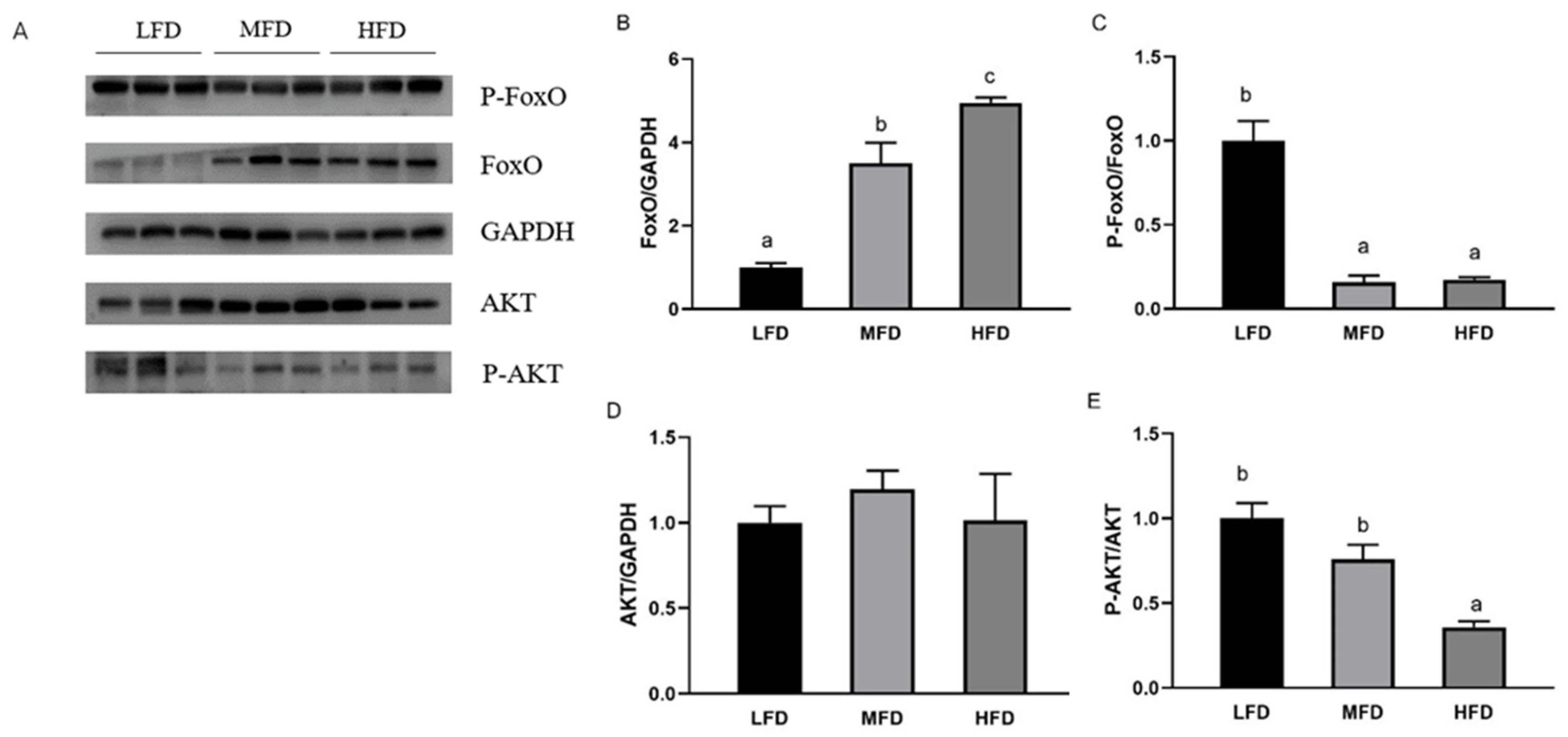
3.5. Effects of Dietary Lipid on Gene and Protein Expressions in Hepatopancreas
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Accili, D.; Arden, K.C. FoxOs at the crossroads of cellular metabolism, differentiation, and transformation. Cell 2004, 117, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Furukawa-Hibi, Y.; Kobayashi, Y.; Chen, C.; Motoyama, N. FOXO transcription factors in cell-cycle regulation and the response to oxidative stress. Antioxid. Redox Signal. 2005, 7, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Dong, H.H. FoxO1: A Conductor of Insulin Signaling to Glucose and Lipid Metabolism; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Zhang, W.; Patil, S.; Chauhan, B.; Guo, S.; Unterman, T.G. FoxO1 regulates multiple metabolic pathways in the liver: Effects on gluconeogenic, glycolytic, and lipogenic gene expression. J. Biol. Chem. 2006, 281, 10105–10117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postic, C.; Dentin, R.; Girard, J. Role of the liver in the control of carbohydrate and lipid homeostasis. Diabetes Metab. 2004, 30, 398–408. [Google Scholar] [CrossRef]
- Kim, D.H.; Perdomo, G.; Zhang, T.; Slusher, S.; Lee, S.; Phillips, B.E.; Fan, Y.; Giannoukakis, N.; Gramignoli, R.; Strom, S. FoxO6 Integrates Insulin Signaling with Gluconeogenesis in the Liver. Diabetes 2011, 60, 2763–2774. [Google Scholar] [CrossRef] [Green Version]
- Calabuig-Navarro, V.; Yamauchi, J.; Lee, S.; Zhang, T.; Liu, Y.Z.; Sadlek, K.; Coudriet, G.M.; Piganelli, J.D.; Jiang, C.L.; Miller, R. Forkhead Box O6 (FoxO6) Depletion Attenuates Hepatic Gluconeogenesis and Protects against Fat-induced Glucose Disorder in Mice. J. Biol. Chem. 2015, 290, 15581–15594. [Google Scholar] [CrossRef] [Green Version]
- Haeusler, R.A.; Hartil, K.; Vaitheesvaran, B.; Arrieta-Cruz, I.; Knight, C.M.; Cook, J.R.; Kammoun, H.L.; Febbraio, M.A.; Gutierrez-Juarez, R.; Kurland, I.J. Integrated control of hepatic lipogenesis versus glucose production requires FoxO transcription factors. Nat. Commun. 2014, 5, 5190. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, I.S.; Zhang, W.; Wasserman, D.H.; Liew, C.W.; Liu, J.; Paik, J.; Depinho, R.A.; Stolz, D.B.; Kahn, C.R.; Schwartz, M.W. FoxO1 integrates direct and indirect effects of insulin on hepatic glucose production and glucose utilization. Nat. Commun. 2015, 6, 7079. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Zhang, Y.; Deng, K.; Liu, G.; Gu, Z.; Liu, J.; Luo, K.; Zhang, W.; Mai, K. Forkhead box O1 in turbot Scophthalmus maximus: Molecular characterization, gene structure, tissue distribution and the role in glucose metabolism. Gene 2019, 708, 49–56. [Google Scholar] [CrossRef]
- Sun, J.; Xiao, P.Z.; Chang, Z.G.; Ji, H.; Du, Z.Y.; Chen, L.Q. Forkhead box O1 in grass carp Ctenopharyngodon idella: Molecular characterization, gene structure, tissue distribution and mRNA expression in insulin-inhibited adipocyte lipolysis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 204, 76–84. [Google Scholar] [CrossRef]
- Li, S.; Sang, C.; Zhang, J.; Li, Z.; Chen, N. Molecular cloning, expression profiling of adipose triglyceride lipase (ATGL) and forkhead box O1 (FoxO1), and effects of dietary carbohydrate level on their expression in hybrid grouper (Epinephelus fuscoguttatus × E. lanceolatus). Aquaculture 2018, 492, 103–122. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, Z.; Zhou, T.; Yang, Y.; Dunhama, R.; Liu, Z. FOXO genes in channel catfish and their response after bacterial infection. Dev. Comp. Immunol. 2019, 97, 38–44. [Google Scholar] [CrossRef]
- Alessi, D.R.; Barry Caudwell, F.; Andjelkovic, M.; Hemmings, B.A.; Cohen, P. Molecular basis for the substrate specificity of protein kinase B; comparison with MAPKAP kinase-1 and p70 S6 kinase. FEBS Lett. 1996, 399, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Dobson, A.J.; Ezcurra, M.; Flanagan, C.E.; Summerfield, A.C.; Piper, M.D.W.; Gems, D.; Alic, N. Nutritional Programming of Lifespan by FOXO Inhibition on Sugar-Rich Diets. Cell Rep. 2017, 18, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Unterman, T.G. Regulation of Hepatic Glucose Metabolism by FoxO Proteins, an Integrated Approach. Curr. Top. Dev. Biol. 2017, 127, 119–147. [Google Scholar]
- Guo, H.; Xia, M.; Zou, T.; Ling, W.; Zhong, R.; Zhang, W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J. Nutr. Biochem. 2012, 23, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Barma, P.; Dey, D.; Basu, D.; Roy, S.S.; Bhattacharya, S. Nutritionally induced insulin resistance in an Indian perch: A possible model for type 2 diabetes. Curr. Sci. 2006, 90, 188–194. [Google Scholar]
- Velasco, C.; Comesaña, S.; Conde-Sieira, M.; Míguez, J.M.; Soengas, J.L. The short-term presence of oleate or octanoate alters the phosphorylation status of Akt, AMPK, mTOR, CREB, and FoxO1 in liver of rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2018, 219–220, 17–25. [Google Scholar] [CrossRef]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Simakov, O.; Marletaz, F.; Cho, S.-J.; Edsinger-Gonzales, E.; Havlak, P.; Hellsten, U.; Kuo, D.-H.; Larsson, T.; Lv, J.; Arendt, D. Insights into bilaterian evolution from three spiralian genomes. Nature 2013, 493, 526–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Xiao, S.; Zhang, Y.; Zhang, Y.; Yu, Z. ChAkt1 involvement in orchestrating the immune and heat shock responses in Crassostrea hongkongensis: Molecular cloning and functional characterization. Fish Shellfish Immunol. 2015, 47, 1015. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Niu, D.; Wei, K.; Dong, Z.; Li, J. Polymorphisms in the FOXO gene are associated with growth traits in the Sanmen breeding population of the razor clam Sinonovacula constricta. Aquac. Fish 2018, 3, 177–183. [Google Scholar] [CrossRef]
- Mai, K.; Mercer, J.P.; Donlon, J. Comparative studies on the nutrition of two species of abalone, Haliotis tuberculata L. and Haliotis discus hannai Ino. III: Response of abalone to various levels of dietary lipid. Aquaculture 1995, 134, 65–80. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, D.; Chen, F.; Ma, S.; Zhou, W.; Zhang, W.; Mai, K. Lipid deposition in abalone Haliotis discus hannai affected by dietary lipid levels through AMPKα2/PPARα and JNK/mTOR/SREBP-1c pathway. Aquaculture 2021, 532, 736040. [Google Scholar] [CrossRef]
- Cheng, P.; Liu, X.; Zhang, G.; He, J. Cloning and expression analysis of a HSP70 gene from Pacific abalone (Haliotis discus hannai). Fish Shellfish Immunol. 2007, 22, 77–87. [Google Scholar] [CrossRef]
- Hu, J.J.; Chen, Y.L.; Duan, X.K.; Jin, T.C.; Li, Y.; Zhang, L.J.; Liu, G.M.; Cao, M.J. Involvement of clip-domain serine protease in the anti-Vibrio immune response of abalone (Haliotis discus hannai)-Molecular cloning, characterization and functional analysis. Fish Shellfish Immunol. 2018, 72, 210–219. [Google Scholar] [CrossRef]
- Varma, D.; Bülow, M.H.; Pesch, Y.Y.; Loch, G.; Hoch, M. Forkhead, a new cross regulator of metabolism and innate immunity downstream of TOR in Drosophila. J. Insect Physiol. 2014, 69, 80–88. [Google Scholar] [CrossRef]
- Murphy, C.T.; Mccarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277. [Google Scholar] [CrossRef] [PubMed]
- Süren-Castillo, S.; Abrisqueta, M.; Maestro, J.L. FoxO inhibits juvenile hormone biosynthesis and vitellogenin production in the German cockroach. Insect Biochem. Mol. Biol. 2012, 42, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Fink, C.; Hoffmann, J.; Knop, M.; Li, Y.; Isermann, K.; Roeder, T. Intestinal FoxO signaling is required to survive oral infection in Drosophila. Mucosal Immunol. 2016, 9, 927–936. [Google Scholar] [CrossRef]
- Kops, G.J.; Dansen, T.B.; Polderman, P.E.; Saarloos, I.; Wirtz, K.W.; Coffer, P.J.; Huang, T.-T.; Bos, J.L.; Medema, R.H.; Burgering, B.M. Forkhead transcription factor FOXO3a protects quiescent cells from oxidative stress. Nature 2002, 419, 316–321. [Google Scholar] [CrossRef]
- Furuyama, T.; Nakazawa, T.; Nakano, I.; Mori, N. Identification of the differential distribution patterns of mRNAs and consensus binding sequences for mouse DAF-16 homologues. Biochem. J. 2000, 349, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Polakof, S.; Mommsen, T.P.; Soengas, J.L. Glucosensing and glucose homeostasis: From fish to mammals. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 160, 123–149. [Google Scholar]
- Xiong, X.; Tao, R.; DePinho, R.A.; Dong, X.C. Deletion of hepatic FoxO1/3/4 genes in mice significantly impacts on glucose metabolism through downregulation of gluconeogenesis and upregulation of glycolysis. PLoS ONE 2013, 8, e74340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haeusler, R.A.; Kaestner, K.H.; Accili, D. FoxOs function synergistically to promote glucose production. J. Biol. Chem. 2010, 285, 35245–35248. [Google Scholar] [CrossRef] [Green Version]
- Barthel, A.; Schmoll, D.; Unterman, T.G. FoxO proteins in insulin action and metabolism. Trends Endocrinol. Metab. 2005, 16, 183–189. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, H.; Park, J.M.; Lm, S.S.; Bae, J.S.; Kim, M.Y.; Yoon, H.G.; Cha, U.Y.; Kim, K.S.; Ahn, Y.H. Interrelationship between Liver X Receptor α, Sterol Regulatory Element-binding Protein-1c, Peroxisome Proliferator-activated Receptor γ, and Small Heterodimer Partner in the Transcriptional Regulation of Glucokinase Gene Expression in Liver. J. Biol. Chem. 2009, 284, 15071–15083. [Google Scholar] [CrossRef] [Green Version]
- Roth, U.; Curth, K.; Unterman, T.G.; Kietzmann, T. The Transcription Factors HIF-1 and HNF-4 and the Coactivator p300 Are Involved in Insulin-regulated Glucokinase Gene Expression via the Phosphatidylinositol 3-Kinase/Protein Kinase B Pathway. J. Biol. Chem. 2004, 279, 2623–2631. [Google Scholar] [CrossRef] [Green Version]
- Koyama, M.; Furukawa, F.; Koga, Y.; Funayama, S.; Okumura, S.I. Gluconeogenesis and glycogen metabolism during development of Pacific abalone, Haliotis discus hannai. AJP Regul. Integr. Comp. Physiol. 2020, 318, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Couto, A.; Enes, P.; Peres, H.; Oliva-Teles, A. Effect of water temperature and dietary starch on growth and metabolic utilization of diets in gilthead sea bream (Sparus aurata) juveniles. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Deng, K.; Pan, M.; Zhang, Y.; Sampath, W.; Zhang, W.; Mai, K. Molecular adaptations of glucose and lipid metabolism to different levels of dietary carbohydrates in juvenile Japanese flounder Paralichthys olivaceus. Aquac. Nutr. 2019, 26, 516–527. [Google Scholar] [CrossRef]
- Petersen, K.F.; Shulman, G.I. Etiology of insulin resistance. Am. J. Med. 2006, 119, S10–S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Ginsberg, H.N. Increased very low density lipoprotein (VLDL) secretion, hepatic steatosis, and insulin resistance. Trends Endocrinol. Metab. 2011, 22, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraegen, E.W.; Clark, P.W.; Jenkins, A.B.; Daley, E.A.; Chisholm, D.J.; Storlien, L.H. Development of muscle insulin resistance after liver insulin resistance in high-fat–fed rats. Diabetes 1991, 40, 1397–1403. [Google Scholar] [CrossRef]
- Shu, L.; Hou, G.; Zhao, H.; Huang, W.; Ma, H. Resveratrol improves high-fat diet-induced insulin resistance in mice by downregulating the lncRNA NONMMUT008655.2. Am. J. Transl. Res. 2020, 12, 1–18. [Google Scholar] [PubMed]
- Lu, K.L.; Xu, W.N.; Wang, L.N.; Zhang, D.D.; Zhang, C.N.; Liu, W.B. Hepatic β-Oxidation and Regulation of Carnitine Palmitoyltransferase (CPT) I in Blunt Snout Bream Megalobrama amblycephala Fed a High Fat Diet. PLoS ONE 2014, 9, e93135. [Google Scholar] [CrossRef] [Green Version]
- Reaven, G.M. Role of insulin resistance in human disease. Diabetes 1988, 37, 1595–1607. [Google Scholar] [CrossRef]
- Luong, N.; Davies, C.R.; Wessells, R.J.; Graham, S.M.; King, M.T.; Veech, R.; Bodmer, R.; Oldham, S.M. Activated FOXO-mediated insulin resistance is blocked by reduction of TOR activity. Cell Metab. 2006, 4, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Nakae, J.; Biggs, W.H.; Kitamura, T.; Cavenee, W.K.; Wright, C.V.; Arden, K.C.; Accili, D. Regulation of insulin action and pancreatic β-cell function by mutated alleles of the gene encoding forkhead transcription factor Foxo1. Nat. Genet. 2002, 32, 245–253. [Google Scholar] [CrossRef]
- Nakae, J.; Oki, M.; Cao, Y. The FoxO transcription factors and metabolic regulation. FEBS Lett. 2008, 582, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Li, P.; Huntley, J.; Chang, J.P.; Arden, K.C.; Olefsky, J.M. FoxO1 haploinsufficiency protects against high-fat diet-induced insulin resistance with enhanced peroxisome proliferator-activated receptor γ activation in adipose tissue. Diabetes 2009, 58, 1275–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Yan, Y.; Tian, H.; Jiang, G.; Li, X.; Liu, W. Resveratrol supplementation improves lipid and glucose metabolism in high-fat diet-fed blunt snout bream. Fish Physiol. Biochem. 2018, 44, 163–173. [Google Scholar] [CrossRef]
- Du, R.-Y.; Chen, J.-X.; Zhu, J.; Feng, J.-Y.; Luo, L.; Lin, S.-M.; Chen, Y.-J. Glucose homeostasis and glucose tolerance were impaired with elevated lipid to starch ratios in practical diets for the omnivorous genetically improved farmed tilapia Oreochromis niloticus. Aquaculture 2020, 523, 735221. [Google Scholar] [CrossRef]
- Figueiredo-Silva, A.C.; Panserat, S.; Kaushik, S.; Geurden, I.; Polakof, S. High levels of dietary fat impair glucose homeostasis in rainbow trout. J. Exp. Biol. 2012, 215, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moller, D.E. New drug targets for type 2 diabetes and the metabolic syndrome. Nature 2001, 414, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Goldstein, J.L. Selective versus total insulin resistance: A pathogenic paradox. Cell Metab. 2008, 7, 95–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asterholm, I.W.; Scherer, P.E. Enhanced metabolic flexibility associated with elevated adiponectin levels. Am. J. Pathol. 2010, 176, 1364–1376. [Google Scholar] [CrossRef]
- Liu, Y.; Turdi, S.; Park, T.; Morris, N.J.; Deshaies, Y.; Xu, A.; Sweeney, G. Adiponectin Corrects High-Fat Diet–Induced Disturbances in Muscle Metabolomic Profile and Whole-Body Glucose Homeostasis. Diabetes 2013, 62, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef]
- Blüher, M. Adipokines—Removing road blocks to obesity and diabetes therapy. Mol. Metab. 2014, 3, 230–240. [Google Scholar] [CrossRef]
- Hotta, K.; Funahashi, T.; Bodkin, N.L.; Ortmeyer, H.K.; Arita, Y.; Hansen, B.C.; Matsuzawa, Y. Circulating Concentrations of the Adipocyte Protein Adiponectin are Decreased in Parallel with Reduced Insulin Sensitivity During the Progression to Type 2 Diabetes in Rhesus Monkeys. Diabetes 2001, 50, 1126–1133. [Google Scholar] [CrossRef] [Green Version]
- Ahren, B.; Mansson, S.; Gingerich, R.L.; Havel, P.J. Regulation of plasma leptin in mice: Influence of age, high-fat diet, and fasting. Am. J. Physiol. 1997, 273, R113. [Google Scholar] [CrossRef] [PubMed]
- Reizes, O.; Benoit, S.C.; Clegg, D.J. Leptin Signaling in the Brain; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Lin, S.; Thomas, T.C.; Storlien, L.H.; Huang, X.F. Development of high fat diet-induced obesity and leptin resistance in C57Bl/6J mice. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 2000, 24, 639. [Google Scholar] [CrossRef] [Green Version]
- Spezani, R.; Silva, R.R.D.; Martins, F.F.; Marinho, T.D.S.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. Intermittent fasting, adipokines, insulin sensitivity, and hypothalamic neuropeptides in a dietary overload with high-fat or high-fructose diet in mice. J. Nutr. Biochem. 2020, 83, 108419. [Google Scholar] [CrossRef]
- Ganjam, G.K.; Dimova, E.Y.; Unterman, T.G.; Kietzmann, T. FoxO1 and HNF-4 Are Involved in Regulation of Hepatic Glucokinase Gene Expression by Resveratrol. J. Biol. Chem. 2009, 284, 30783–30797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Nutritional regulation of hepatic glucose metabolism in fish. Fish Physiol. Biochem. 2009, 35, 519–539. [Google Scholar] [CrossRef] [PubMed]
- Peres, H.; Oliva-Teles, A. Utilization of raw and gelatinized starch by European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2002, 205, 287–299. [Google Scholar] [CrossRef]
- Surintorn, B.; Araya, J.; Suksan, K.; Elisabeth, P.J.; Vincent, V.; Christine, B.; Lucie, M.; Stephane, P. Adaptation of Nile tilapia (Oreochromis niloticus) to different levels of dietary carbohydrates: New insights from a long term nutritional study. Aquaculture 2018, 496, 58–65. [Google Scholar]
- Tsutomu, W.; Hiroki, K.; Yusuke, M.; Motohiro, S.; Minoru, O.; Masakiyo, S.; Daisuke, K.; Hiroshi, T.; Toshiyasu, S. Spironolactone improves glucose and lipid metabolism by ameliorating hepatic steatosis and inflammation and suppressing enhanced gluconeogenesis induced by high-fat and high-fructose diet. Endocrinology 2010, 151, 2040–2049. [Google Scholar]
- Schmoll, D.; Walker, K.S.; Alessi, D.R.; Grempler, R.; Burchell, A.; Guo, S.; Walther, R.; Unterman, T.G. Regulation of Glucose-6-phosphatase Gene Expression by Protein Kinase Bα and the Forkhead Transcription Factor FKHR Evidence for insulin response unit-dependent and-independent effects of insulin on promoter activity. J. Biol. Chem. 2000, 275, 36324–36333. [Google Scholar] [CrossRef] [Green Version]
- Yeagley, D.; Guo, S.; Unterman, T.; Quinn, P.G. Gene- and activation-specific mechanisms for insulin inhibition of basal and glucocorticoid-induced insulin-like growth factor binding protein-1 and phosphoenolpyruvate carboxykinase transcription. Roles of forkhead and insulin response sequences. J. Biol. Chem. 2001, 276, 33705–33710. [Google Scholar] [CrossRef] [Green Version]
- Herzig, S.; Long, F.; Jhala, U.S.; Hedrick, S.; Quinn, R.; Bauer, A.; Rudolph, D.; Schutz, G.; Yoon, C.; Puigserver, P. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1. Nature 2001, 413, 179–183. [Google Scholar] [CrossRef]
- Puigserver, P.; Rhee, J.; Donovan, J.; Walkey, C.J.; Yoon, J.C.; Oriente, F.; Kitamura, Y.; Altomonte, J.; Dong, H.; Accili, D. Insulin-regulated hepatic gluconeogenesis through FOXO1–PGC-1α interaction. Nature 2003, 423, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Daitoku, H.; Yamagata, K.; Matsuzaki, H.; Hatta, M.; Fukamizu, A. Regulation of PGC-1 promoter activity by protein kinase B and the forkhead transcription factor FKHR. Diabetes 2003, 52, 642–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, S.H.; Satoh, H.; Herzig, S.; Lee, C.H.; Hedrick, S.; Kulkarni, R.; Evans, R.M.; Olefsky, J.; Montminy, M. PGC-1 promotes insulin resistance in liver through PPAR-α-dependent induction of TRB-3. Nat. Med. 2004, 10, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Scarpulla, R.C. Metabolic control of mitochondrial biogenesis through the PGC-1 family regulatory network. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Gross, M.; Lee, D.H.; Holvoet, P.; Himes, J.H.; Shikany, J.M.; Jacobs, D.R. Oxidative Stress and Insulin Resistance. Diabetes Care 2009, 32, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, D.; Yang, C.; Luo, Q. Soy isoflavones improve the oxidative stress induced hypothalamic inflammation and apoptosis in high fat diet-induced obese male mice through PGC1-α pathway. Aging 2020, 12, 8710. [Google Scholar] [CrossRef]
- Gu, L.; Ding, X.; Wang, Y.; Gu, M.; Peng, Y. Spexin alleviates insulin resistance and inhibits hepatic gluconeogenesis via the FoxO1/PGC-1α pathway in high-fat-diet-induced rats and insulin resistant cells. Int. J. Biol. Sci. 2019, 15, 2815–2829. [Google Scholar] [CrossRef] [Green Version]
- Wredenberg, A.; Freyer, C.; Sandström, M.E.; Katz, A.; Wibom, R.; Westerblad, H.; Larsson, N.-G. Respiratory chain dysfunction in skeletal muscle does not cause insulin resistance. Biochem. Biophys. Res. Commun. 2006, 350, 202–207. [Google Scholar] [CrossRef]
- Koistinen, H.A.; Bastard, J.P.; Dusserre, E.; Ebeling, P.; Zegari, N.; Andreelli, F.; Jardel, C.; Donner, M.; Meyer, L.; Moulin, P.; et al. Subcutaneous adipose tissue expression of tumour necrosis factor- α is not associated with whole body insulin resistance in obese nondiabetic or in type-2 diabetic subjects. Eur. J. Clin. Investig. 2000, 30, 302–310. [Google Scholar] [CrossRef]
- Sell, H.; Laurencikiene, J.; Taube, A.; Eckardt, K.; Cramer, A.; Horrighs, A.; Arner, P.; Eckel, J. Chemerin Is a Novel Adipocyte-Derived Factor Inducing Insulin Resistance in Primary Human Skeletal Muscle Cells. Diabetes 2009, 58, 2731–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, M.A.; Sheridan, M.A. New insights into the signaling system and function of insulin in fish. Gen. Comp. Endocrinol. 2011, 173, 227–247. [Google Scholar] [CrossRef]
- Kwon, H.; Pessin, J.E. Adipokines Mediate Inflammation and Insulin Resistance. Front. Endocrinol. 2013, 4, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Guo, Q.; Qin, C.; Shang, R.; Zhang, Z. Sea Buckthorn Fruit Oil Extract Alleviates Insulin Resistance through the PI3K/Akt Signaling Pathway in Type 2 Diabetes Mellitus Cells and Rats. J. Agric. Food Chem. 2017, 65, 1328–1336. [Google Scholar] [CrossRef]
- Motawi, T.M.K.; Bustanji, Y.; El-Maraghy, S.A.; Taha, M.O.; Ghussein, M.A.S.A. Naproxen and Cromolyn as New Glycogen Synthase Kinase 3β Inhibitors for Amelioration of Diabetes and Obesity: An Investigation by Docking Simulation and Subsequent In Vitro/In Vivo Biochemical Evaluation. J. Biochem. Mol. Toxicol. 2013, 27, 425–436. [Google Scholar] [CrossRef]
- Lappas, M. GSK3β is increased in adipose tissue and skeletal muscle from women with gestational diabetes where it regulates the inflammatory response. PLoS ONE 2014, 9, e115854. [Google Scholar]
- Shieh, J.M.; Wu, H.T.; Cheng, K.C.; Cheng, J.T. Melatonin ameliorates high fat diet-induced diabetes and stimulates glycogen synthesis via a PKCzeta-Akt-GSK3beta pathway in hepatic cells. J. Pineal Res. 2010, 47, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Agius, L. Role of glycogen phosphorylase in liver glycogen metabolism. Mol. Asp. Med. 2015, 46, 34–45. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Target Sequence |
|---|---|
| siRNA-335 | CAGCTGGTTGGAAGAATTC |
| siRNA-626 | CACGAGCTAGTTCTAATGC |
| siRNA-1837 | AAGCAAGAGTTGTCTCTAG |
| siRNA-NC | TTCTCCGAACGTGTCACGT |
| Genes | 5′/3′ Forward Primer | 5′/3′ Reverse Primer | Accession Number |
|---|---|---|---|
| foxo | AATGGCCCTGTTTCAACCAC | CTTCCCGACTGTAAAGGTGT | MN864138 |
| pepck | TCGACAACAATGGCAAGCTC | CTTGTCTCCGCAACATTCGT | MH220521.1 |
| g6pase | CGTACAAACTGCCTCACTCG | ATTCTCGGGACAATGTTCACAA | LC456704.1 |
| hk | ACGCCAGATCAACTCTCGAA | GCATCACACGCTTGTAGGTCA | MH220519.1 |
| pk | AGGCAAGAACATCCGCATC | CTCCTGCAAGATCTCATCGAAC | MH220522.1 |
| tk | TCCCAGAACGCTTCATCGAG | CACAGCCAATTCCGACACCA | MT887625 |
| g6pdh | TGCCACCATCAGTCTTCGAG | AGATCATGTCCAAACGGCTT | MT551204 |
| gsk3β | AACTGTTCCGAAGTCTTGCAT | ATATCCCGATGACAAACTCCT | FJ435173.1 |
| glut1 | TCCAGTTTGGCTACAATACAGG | CCCATTCCGATCAAAGTAGGTC | MT551207 |
| pyg | TCTCGTGTTCTGTACCCCAA | TGAACCTGCGTACAATGTCC | LC456706.1 |
| pgc1α | CGAAGACCCAGCAGTCACC | TAACGATCAGTGTCACGACCT | MT873877 |
| β-actin | CCTCAAGTACCCCATCGAGCAC | ATCTTCTCCATGTCGTCCCAG | AY380809.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Guo, Y.; Pan, M.; Li, X.; Huang, D.; Liu, Y.; Wu, C.; Zhang, W.; Mai, K. Functions of Forkhead Box O on Glucose Metabolism in Abalone Haliotis discus hannai and Its Responses to High Levels of Dietary Lipid. Genes 2021, 12, 297. https://doi.org/10.3390/genes12020297
Wang L, Guo Y, Pan M, Li X, Huang D, Liu Y, Wu C, Zhang W, Mai K. Functions of Forkhead Box O on Glucose Metabolism in Abalone Haliotis discus hannai and Its Responses to High Levels of Dietary Lipid. Genes. 2021; 12(2):297. https://doi.org/10.3390/genes12020297
Chicago/Turabian StyleWang, Liu, Yanlin Guo, Mingzhu Pan, Xinxin Li, Dong Huang, Yue Liu, Chenglong Wu, Wenbing Zhang, and Kangsen Mai. 2021. "Functions of Forkhead Box O on Glucose Metabolism in Abalone Haliotis discus hannai and Its Responses to High Levels of Dietary Lipid" Genes 12, no. 2: 297. https://doi.org/10.3390/genes12020297
APA StyleWang, L., Guo, Y., Pan, M., Li, X., Huang, D., Liu, Y., Wu, C., Zhang, W., & Mai, K. (2021). Functions of Forkhead Box O on Glucose Metabolism in Abalone Haliotis discus hannai and Its Responses to High Levels of Dietary Lipid. Genes, 12(2), 297. https://doi.org/10.3390/genes12020297