
Expression of Estrogen Receptor- and Progesterone Receptor-Regulating MicroRNAs in Breast Cancer

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Samples
2.2. MicroRNA Isolation
2.3. MiRNA Reverse Transcription and Real-Time PCR (RT-PCR)
2.4. Statistical Analysis
3. Results
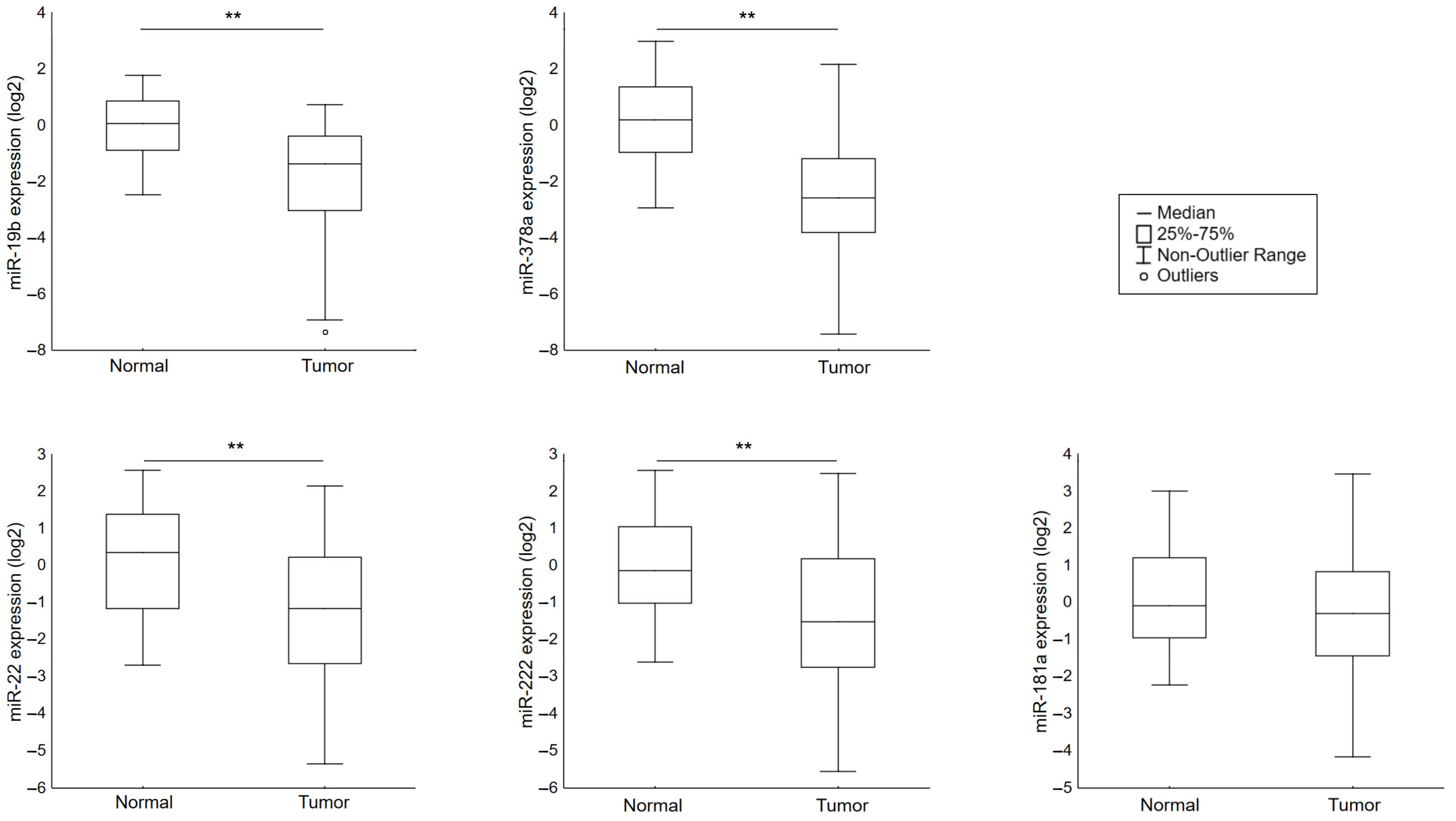
3.1. Analysis of miR-19b, miR-22, miR-222, miR-378a, miR-181a Expression in Breast Cancer
3.2. Analysis of the Association of MiR-19b, MiR-22, MiR-222, MiR-378a, and MiR-181a Expression with ER, PR, HER2 Status and Ki-67 Index
3.3. Expression of MiR-19b, MiR-22, MiR-222, MiR-378a, and MiR-181a in Relation to Clinicopathologic Features of ER- and/or PR-Positive BC and ER-and-PR-Negative BC
3.4. Expression of MiR-19b, MiR-22, MiR-222, MiR-378a, MiR-181a in Relation to Clinicopathologic Features of HER2-Positive BC or HER2-Negative BC
3.5. Expression of MiR-19b, MiR-22, MiR-222, MiR-378a, and MiR-181a in Relation to Clinicopathologic Features of Tumors with the Ki-67 < 14% or ≥ 14%.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagai, H.; Kim, Y.H. Cancer prevention from the perspective of global cancer burden patterns. J. Thorac. Dis. 2017, 9, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Rosso, K.; Nathanson, S.D. Pathogenesis, prevention, diagnosis and treatment of breast cancer. World J. Clin. Oncol. 2014, 5, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Cicatiello, L.; Addeo, R.; Sasso, A.; Altucci, L.; Petrizzi, V.B.; Borgo, R.; Cancemi, M.; Caporali, S.; Caristi, S.; Scafoglio, C.; et al. Estrogens and progesterone promote persistent CCND1 gene activation during G1 by inducing transcriptional derepression via c-Jun/c-Fos/estrogen receptor (progesterone receptor) complex assembly to a distal regulatory element and recruitment of cyclin D1 to its own gene promoter. Mol. Cell. Biol. 2004, 24, 7260–7274. [Google Scholar]
- Hyder, S.M. The role of steroid hormones on the regulation of vascular endothelial growth factor. Am. J. Pathol. 2002, 161, 345–346. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Mayer, J.A.; Mazumdar, A.; Fertuck, K.; Kim, H.; Brown, M.; Brown, P.H. Estrogen induces c-myc gene expression via an upstream enhancer activated by the estrogen receptor and the AP-1 transcription factor. Mol. Endocrinol. 2011, 25, 1527–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Song, S.; Jiang, Z.F.; Sun, B.; Wang, T.; Zhang, S.h.; Wu, S.h. Receptor conversion in metastatic breast cancer: A prognosticator of survival. Oncotarget 2016, 7, 71887–71903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, H.; Zhang, H.; Kong, Q.; Jiang, Y. Mechanisms for estrogen receptor expression in human cancer. Exp. Hematol. Oncol. 2018, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, J.M.; Clark, G.M.; Osborne, C.K.; Allred, D.C. Estrogen receptor status by immunohistochemistry is superior to the ligand-binding assay for predicting response to adjuvant endocrine therapy in breast cancer. J. Clin. Oncol. 1999, 17, 1474–1481. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, T.S.; Kononchuk, V.V.; Yakovleva, A.K.; Alekseenok, E.Y.; Sidorov, S.V.; Gulyaeva, L.F. Association between lymph node status and expression levels of androgen receptor, miR-185, miR-205, and miR-21 in breast cancer subtypes. Int. J. Breast Cancer 2020, 2020, 3259393. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 2005, 33, e179. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar]
- Gilam, A.; Shai, A.; Ashkenazi, I.; Sarid, L.A.; Drobot, A.; Bickel, A.; Shomron, N. MicroRNA regulation of progesterone receptor in breast cancer. Oncotarget 2017, 8, 25963–25976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Li, J.; Sun, M.; Sun, L.; Zhang, X. miRNA expression in breast cancer varies with lymph node metastasis and other clinicopathologic features. IUBMB Life 2014, 66, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Horie-Inoue, K.; Ueno, T.; Suzuki, T.; Sato, W.; Shigekawa, T.; Osaki, A.; Saeki, T.; Berezikov, E.; Mano, H.; et al. miR-378a-3p modulates tamoxifen sensitivity in breast cancer MCF-7 cells through targeting GOLT1A. Sci. Rep. 2015, 5, 13170. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Yu, D.; Wei, N.; Fu, H.; Cai, T.; Huang, Y.; Wu, C.; Zheng, X.; Du, Q.; Lin, D.; et al. An estrogen receptor alpha suppressor, microRNA-22, is downregulated in estrogen receptor alpha-positive human breast cancer cell lines and clinical samples. FEBS J. 2010, 277, 1684–1694. [Google Scholar] [CrossRef]
- Zou, Q.; Tang, Q.; Pan, Y.; Wang, X.; Dong, X.; Liang, Z.; Huang, D. MicroRNA-22 inhibits cell growth and metastasis in breast cancer via targeting of SIRT1. Exp. Ther. Med. 2017, 14, 1009–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stinson, S.; Lackner, M.R.; Adai, A.T.; Yu, N.; Kim, H.J.; O’Brien, C.; Spoerke, J.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. TRPS1 targeting by miR-221/222 promotes the epithelial-to-mesenchymal transition in breast cancer. Sci. Signal. 2011, 4, 41. [Google Scholar]
- Chernyy, V.; Pustylnyak, V.; Kozlov, V.; Gulyaeva, L. Increased expression of miR-155 and miR-222 is associated with lymph node positive status. J. Cancer 2018, 9, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Han, S.H.; Kim, H.J.; Gwak, J.M.; Kim, M.; Chung, Y.R.; Park, S.Y. MicroRNA-222 Expression as a predictive marker for tumor progression in hormone receptor-positive breast cancer. J. Breast Cancer 2017, 20, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Yoshimoto, N.; Toyama, T.; Takahashi, S.; Sugiura, H.; Endo, Y.; Iwasa, M.; Fujii, Y.; Yamashita, H. Distinct expressions of microRNAs that directly target estrogen receptor α in human breast cancer. Breast Cancer Res. Treat. 2011, 30, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Lei, Q.; Wang, Y.; Wang, Z.; Xie, G.; Zhong, X.; Wang, Y.; Chen, N.; Qiu, Y.; Pu, T.; et al. Downregulation of miR-221-3p and upregulation of its target gene PARP1 are prognostic biomarkers for triple negative breast cancer patients and associated with poor prognosis. Oncotarget 2017, 8, 108712–108725. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Guo, L.; Jiang, F.; Li, L.; Li, Z.; Chen, F. Analysis of the miRNA-mRNA-lncRNA networks in ER+ and ER- breast cancer cell lines. J. Cell. Mol. Med. 2015, 19, 2874–2887. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, J.; Ma, Z.; Zhang, F.; Yu, W. miR-19b serves as a prognostic biomarker of breast cancer and promotes tumor progression through PI3K/AKT signaling pathway. Onco Targets Ther. 2018, 11, 4087–4095. [Google Scholar] [CrossRef] [Green Version]
- Yin, R.; Guo, L.; Gu, J.; Li, C.; Zhang, W. Overexpressing miR-19b-1 suppress breast cancer growth by inhibiting tumor microenvironment induced angiogenesis. Int. J. Biochem. Cell Biol. 2018, 97, 43–51. [Google Scholar] [CrossRef]
- Anfossi, S.; Giordano, A.; Gao, H.; Cohen, E.N.; Tin, S.; Wu, Q.; Garza, R.J.; Debeb, B.G.; Alvarez, R.H.; Valero, V.; et al. High serum miR-19a levels are associated with inflammatory breast cancer and are predictive of favorable clinical outcome in patients with metastatic HER2+ inflammatory breast cancer. PLoS ONE 2014, 9, e83113. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Toyama, T.; Sugiura, H.; Fujii, Y.; Yamashita, H. miR-206 Expression is down-regulated in estrogen receptor alpha-positive human breast cancer. Cancer Res. 2008, 68, 5004–5008. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hong, F.; Yu, Z. Decreased expression of microRNA-206 in breast cancer and its association with disease characteristics and patient survival. J. Int. Med. Res. 2013, 41, 596–602. [Google Scholar] [CrossRef]
- Ahmad, A.; Ginnebaugh, K.R.; Sethi, S.; Chen, W.; Ali, R.; Mittal, S.; Sarkar, F.H. miR-20b is up-regulated in brain metastases from primary breast cancers. Oncotarget 2015, 6, 12188–12195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, L.; Giamas, G.; Jacob, J.; Coombes, R.C.; Lucchesi, W.; Thiruchelvam, P.; Barton, G.; Jiao, L.R.; Wait, R.; Waxman, J.; et al. The estrogen receptor-alpha-induced microRNA signature regulates itself and its transcriptional response. Proc. Natl. Acad. Sci. USA 2009, 106, 15732–15737. [Google Scholar] [CrossRef] [Green Version]
- Luengo-Gil, G.; García-Martínez, E.; Chaves-Benito, A.; Conesa-Zamora, P.; Navarro-Manzano, E.; González-Billalabeitia, E.; García-Garre, E.; Martínez-Carrasco, A.; Vicente, V.; Ayala de la Peña, F. Clinical and biological impact of miR-18a expression in breast cancer after neoadjuvant chemotherapy. Cell Oncol. 2019, 42, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Tokar, T.; Pastrello, C.; Rossos, A.E.M.; Abovsky, M.; Hauschild, A.C.; Tsay, M.; Lu, R.; Jurisica, I. mirDIP 4.1-integrative database of human microRNA target predictions. Nucleic Acids Res. 2018, 46, D360–D370. [Google Scholar] [CrossRef] [PubMed]
- Pandey, D.P.; Picard, D. miR-22 inhibits estrogen signaling by directly targeting the estrogen receptor alpha mRNA. Mol. Cell. Biol. 2009, 29, 3783–3790. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.J.; Lin, J.; Yang, H.; Kong, W.; He, L.; Ma, X.; Coppola, D.; Cheng, J.Q. MicroRNA-221/222 negatively regulates estrogen receptor alpha and is associated with tamoxifen resistance in breast cancer. J. Biol. Chem. 2008, 283, 31079–31086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, H.; Chuang, T.D.; Luo, X.; Chegini, N. Endometrial miR-181a and miR-98 expression is altered during transition from normal into cancerous state and target PGR, PGRMC1, CYP19A1, DDX3X, and TIMP3. J. Clin. Endocrinol. Metab. 2012, 97, E1316-26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toms, D.; Xu, S.; Pan, B.; Wu, D.; Li, J. Progesterone receptor expression in granulosa cells is suppressed by microRNA-378-3p. Mol. Cell. Endocrinol. 2015, 399, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Lambein, K.; Van Bockstal, M.; Vandemaele, L.; Geenen, S.; Rottiers, I.; Nuyts, A.; Matthys, B.; Praet, M.; Denys, H.; Libbrecht, L. Distinguishing score 0 from score 1+ in HER2 immunohistochemistry-negative breast cancer: Clinical and pathobiological relevance. Am. J. Clin. Pathol. 2013, 140, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Schettini, F.; Chic, N.; Brasó-Maristany, F.; Paré, L.; Pascual, T.; Conte, B.; Martínez-Sáez, O.; Adamo, B.; Vidal, M.; Barnadas, E.; et al. Clinical, pathological, and PAM50 gene expression features of HER2-low breast cancer. NPJ Breast Cancer 2021, 7, 1. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Kurozumi, S.; Yamaguchi, Y.; Kurosumi, M.; Ohira, M.; Matsumoto, H.; Horiguchi, J. Recent trends in microRNA research into breast cancer with particular focus on the associations between microRNAs and intrinsic subtypes. J. Hum. Genet. 2017, 62, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Tang, H.; Liu, X.; Liu, P.; Yang, L.; Xie, X.; Ye, F.; Song, C.; Xie, X.; Wei, W. miR-22 as a prognostic factor targets glucose transporter protein type 1 in breast cancer. Cancer Lett. 2015, 356, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Gold, E.B. The timing of the age at which natural menopause occurs. Obstet. Gynecol. Clin. North. Am. 2011, 38, 425–440. [Google Scholar] [CrossRef] [Green Version]
- Zong, Y.; Zhang, Y.; Sun, X.; Xu, T.; Cheng, X.; Qin, Y. miR-221/222 promote tumor growth and suppress apoptosis by targeting lncRNA GAS5 in breast cancer. Biosci. Rep. 2019, 39, BSR20181859. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Maleki, E.; Ghaedi, K.; Shahanipoor, K.; Karimi Kurdistani, Z. Down-regulation of microRNA-19b in hormone receptor-positive/HER2-negative breast cancer. APMIS 2018, 126, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Tabatabaei, S.N.; Ruan, X.; Hardy, P. The dual regulatory role of miR-181a in breast cancer. Cell. Physiol. Biochem. 2017, 44, 843–856. [Google Scholar] [CrossRef]
- Li, J.Y.; Ou, Z.L.; Yu, S.J.; Gu, X.L.; Yang, C.; Chen, A.X.; Di, G.H.; Shen, Z.Z.; Shao, Z.M. The chemokine receptor CCR4 promotes tumor growth and lung metastasis in breast cancer. Breast Cancer Res. Treat. 2012, 131, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, S.M.; Paplomata, E.; Peake, B.M.; Sanabria, E.; Chen, Z.; Nahta, R. P38 MAPK contributes to resistance and invasiveness of HER2- overexpressing breast cancer. Curr. Med. Chem. 2014, 21, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirouac, D.C.; Du, J.; Lahdenranta, J.; Onsum, M.D.; Nielsen, U.B.; Schoeberl, B.; McDonagh, C.F. HER2+ Cancer Cell Dependence on PI3K vs. MAPK Signaling Axes Is Determined by Expression of EGFR, ERBB3 and CDKN1B. PLoS Comput. Biol. 2016, 12, e1004827. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, C.C.; Suman, V.J.; Goetz, M.P. The emerging role of CDK4/6i in HER2-positive breast cancer. Ther. Adv. Med. Oncol. 2019, 11, 1758835919887665. [Google Scholar] [CrossRef]
- Eichner, L.J.; Perry, M.C.; Dufour, C.R.; Bertos, N.; Park, M.; St-Pierre, J.; Giguère, V. miR-378(∗) mediates metabolic shift in breast cancer cells via the PGC-1β/ERRγ transcriptional pathway. Cell Metab. 2010, 12, 352–361. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Characteristics | ER- And/Or PR-Positive (n = 130) | ER-and-PR-Negative (n = 44) | HER2 2+, 3+ (n = 43) | HER2 1+ (n = 31) | HER2 0 (n = 100) | Triple-Negative (n = 20) | |
|---|---|---|---|---|---|---|---|
| Age (mean and range, year) | 60 (27–90) | 52 (38–76) | 56 (39–90) | 57 (44–84) | 58 (27–83) | 54 (38–76) | |
| T stage | T1 | 46 | 18 | 16 | 8 | 40 | 9 |
| T2 | 77 | 23 | 22 | 22 | 56 | 10 | |
| T3 | 2 | 1 | 3 | - | - | - | |
| T4 | 5 | 2 | 2 | 1 | 4 | 1 | |
| N stage | N0 | 70 | 29 | 27 | 11 | 61 | 16 |
| N1 | 46 | 8 | 10 | 15 | 29 | 3 | |
| N2 | 10 | 6 | 5 | 3 | 8 | 1 | |
| N3 | 4 | 1 | 1 | 2 | 2 | - | |
| Ki-67 index | <14% | 73 | 7 | 14 | 14 | 52 | 3 |
| 14–39% | 47 | 19 | 20 | 13 | 33 | 8 | |
| ≥40% | 10 | 18 | 9 | 4 | 15 | 9 | |
| ER score | 0–2 | 4 | 44 | 18 | 7 | 23 | 20 |
| 3–5 | 13 | - | 2 | 1 | 10 | - | |
| 6–8 | 113 | - | 23 | 23 | 67 | - | |
| PR score | 0–2 | 22 | 44 | 24 | 13 | 29 | 20 |
| 3–5 | 34 | - | 7 | 3 | 24 | - | |
| 6–8 | 74 | - | 12 | 15 | 47 | - | |
| HER2 score | 0 | 80 | 20 | - | 100 | 20 | |
| 1 | 23 | 8 | - | 31 | - | - | |
| 2–3 | 27 | 16 | 43 | - | - | - | |
| miRNA | Target (Score Class *) | Other Targets Associated with BC ** | Association with ER or PR Status | Association with Tumor Characteristics |
|---|---|---|---|---|
| hsa-miR-181a-5p | PR; ER (very high) | NLK, GATA6, BCL2, ATM, HIPK2, HRAS, SIRT1, DUSP5, FOS, MTMR3, XIAP | The averages of the expression fold change of miR-181a was significantly lower in the PR-positive group compared to the PR-negative group of BC patients [12]. | MiRNA was downregulated in lymph node metastasis group of BC patients [13]. |
| hsa-miR-378a-3p | PR; ER (medium) | VEGFA, NPNT, MYC, CYP19A1 | Not available, but miR-378a-3p expression was down-regulated in tamoxifen-resistant MCF-7 cells [14]. | Lower expression levels of miR-378a-3p were associated with poor prognosis for tamoxifen-treated patients [14]. |
| hsa-miR-22-3p | ER; PR (high) | BMP7, MYCBP, RGS2, NCOA1, ERBB3, PTEN, SP1 | ER-positive breast tumor specimens had significantly lower miR-22 levels than ER-negative specimens [15]. | Downregulation of miR-22 was significantly associated with the poor differentiation, advanced clinical stage, as well as lymphatic and distant metastasis in breast cancer [16]. |
| hsa-miR-222-3p | ER; PR (high) | STAT5A, MMP1, FOXO3, CDKN1C, FOS, ICAM1, PTEN, ETS1, RECK, TRPS1, CERS2, DKK2 | MiR-221/222 was more abundant in triple-negative tumors than in ER/PR–positive tumors [17]. MiR-222, but not miR-221, led to suppress the expression of the luciferase reporter plasmid carrying the full-length 3′-UTR of ESR1 mRNA signal by more than 40% [15]. | Increased expression of miR-222 was associated with lymph node positive status [18]. High expression levels of miR-222 in patients with ER+ breast cancer were associated with decreased disease-free survival as compared to ER+ breast cancer patients exhibiting low levels of miR-222 [19]. |
| hsa-miR-221-3p | FOXO3, CDKN1C, TBK1, DKK2, BRAP, USP18, ICAM1, FOS, BNIP3, PTEN, ETS1, TRPS1, CERS2, STAT5A, RECK | Expression level of miR-221 was significantly lower in ERa-negative than in ERa-positive tumors [20]. | Low miR-221-3p expression may contribute to the poor outcome of triple negative BC patients [21]. | |
| hsa-miR-19b-3p | ER; PR (high) | PTEN, MYLIP, SOCS1, TLR2, CYP19A1, TGFBR2, SMAD4, MYCN | The expression of both miR-19a and b in BC tissue samples with ER-positive status were down-regulated compared to those with ER-negative. The most significant difference was for the MiR-19b [22]. | The miR-19b expression was associated with distant metastasis and TNM stage [23]. MiR-19b-1 stable overexpression in MDA-MB-231 caused tumor growth arresting entirely [24]. |
| hsa-miR-19a-3p | HOXA5, MECP2, PTEN, CCND1, ERBB4, SOCS1, TLR2, TGFBR2, SMAD4, TNF, ABCA1, MYCN | High serum miR-19a levels are associated with inflammatory breast cancer [25]. | ||
| hsa-miR-206 | ER; PR (very high) | NOTCH3, CCND2, CDK4, TKT, CCND1 | The expression of miR-206 was inversely correlated with ER mRNA level in breast cancer tissues [26]. | Decreased miR-206 was significantly associated with advanced clinical stage and lymph node metastasis [27]. |
| hsa-miR-20b-5p | ER; PR (high) | CDKN1A, MYLIP, VEGFA, EPHB4, LIMK1, PTEN | - | High expression of miR-20b associated with breast cancer brain metastasis [28]. |
| hsa-miR-18a-5p | ER; PR (medium) | PTEN, TNFSF11, NR3C1, HIF1A, ATM, TGFBR2, SMAD4, RUNX1, CTGF | Pre-miR-18a levels were significantly higher in ER-positive tumors compared with negative tumors [29]. Expression levels of miR-18a were strongly and negatively correlated with PR protein scores. MiR-18a expression was much higher in ER-negative than in ER-positive tumors [20]. | A relatively high miR-18a expression was associated with a poor prognosis, especially in the luminal subtype [30]. |
| hsa-miR-18b-5p | MDM2, CTGF | MiR-18b expression was not correlated with ERa protein expression [20]. | Low miR-18b expression was significantly associated with improved survival in HER2-negative breast cancer [20]. |
| Characteristics | n | Relative Level * of miRNA and p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| miR-19b | p-Value | miR-22 | p-Value | miR-222 | p-Value | miR-378a | p-Value | miR-181a | p-Value | |||
| ER and PR status | ER+ and/or PR+ | 130 | 0.48 | 0.974 | 0.46 | 0.533 | 0.41 | 0.535 | 0.20 | 0.407 | 0.85 | 0.518 |
| ER− and PR− | 44 | 0.46 | 0.57 | 0.54 | 0.24 | 0.97 | ||||||
| HER2 status | HER2+ | 43 | 0.55 | 0.345 | 0.48 | 0.470 | 0.70 | 0.181 | 0.44 | 0.013 | 1.33 | 0.015 |
| HER2− | 131 | 0.42 | 0.48 | 0.41 | 0.18 | 0.74 | ||||||
| Ki-67 index (%) | <14 | 80 | 0.52 | 0.335 | 0.76 | 0.043 | 0.46 | 0.537 | 0.24 | 0.407 | 0.85 | 0.904 |
| ≥14 | 94 | 0.38 | 0.36 | 0.43 | 0.19 | 0.96 | ||||||
| Age | ≤50 | 58 | 0.59 | 0.238 | 0.97 | 0.002 | 0.54 | 0.283 | 0.36 | 0.003 | 1.24 | 0.133 |
| >50 | 116 | 0.40 | 0.36 | 0.41 | 0.14 | 0.79 | ||||||
| Characteristics | n | Relative Level * of miRNA and p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| miR-19b | p-Value | miR-22 | p-Value | miR-222 | p-Value | miR-378a | p-Value | miR-181a | p-Value | |||
| ER+ and/or PR+ | ||||||||||||
| T stage | T1 | 46 | 0.80 | 0.088 | 0.85 | 0.124 | 0.92 | 0.009 | 0.27 | 0.152 | 1.22 | 0.332 |
| T2–T4 | 84 | 0.36 | 0.39 | 0.29 | 0.18 | 0.80 | ||||||
| N stage | N0 | 70 | 0.58 | 0.173 | 0.70 | 0.301 | 0.44 | 0.241 | 0.24 | 0.593 | 1.20 | 0.492 |
| N1–N3 | 60 | 0.37 | 0.38 | 0.43 | 0.18 | 0.80 | ||||||
| Ki-67 index (%) | <M ** | 66 | 0.52 | 0.507 | 0.62 | 0.176 | 0.43 | 0.308 | 0.18 | 0.817 | 0.77 | 0.551 |
| ≥M ** | 64 | 0.38 | 0.34 | 0.40 | 0.22 | 1.03 | ||||||
| ER score | 6–8 | 113 | 0.47 | 0.960 | 0.49 | 0.872 | 0.32 | 0.477 | 0.19 | 0.968 | 0.82 | 0.577 |
| 0–5 | 17 | 0.65 | 0.40 | 0.28 | 0.12 | 0.60 | ||||||
| PR score | 6–8 | 74 | 0.38 | 0.090 | 0.48 | 0.841 | 0.41 | 0.819 | 0.18 | 0.454 | 0.77 | 0.495 |
| 0–5 | 56 | 0.58 | 0.37 | 0.38 | 0.24 | 0.88 | ||||||
| ER− and PR− | ||||||||||||
| T stage | T1 | 18 | 0.48 | 0.858 | 0.78 | 0.625 | 0.60 | 0.321 | 0.34 | 0.808 | 1.01 | 0.775 |
| T2–T4 | 26 | 0.43 | 0.48 | 0.47 | 0.20 | 0.90 | ||||||
| N stage | N0 | 29 | 0.46 | 0.958 | 0.49 | 0.903 | 0.54 | 0.825 | 0.23 | 0.722 | 1.11 | 0.445 |
| N1–N3 | 15 | 0.37 | 0.62 | 0.64 | 0.19 | 0.80 | ||||||
| Ki-67 index (%) | ≤M | 23 | 0.51 | 0.158 | 0.96 | 0.024 | 0.71 | 0.027 | 0.44 | 0.035 | 1.71 | 0.003 |
| >M | 21 | 0.28 | 0.36 | 0.28 | 0.10 | 0.58 | ||||||
| Characteristics | n | Relative Level * of miRNA and p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| miR-19b | p-Value | miR-22 | p-Value | miR-222 | p-Value | miR-378a | p-Value | miR-181a | p-Value | |||
| HER2 1+, 2+, 3+ | ||||||||||||
| T stage | T1 | 24 | 0.83 | 0.048 | 1.30 | 0.037 | 1.00 | 0.004 | 0.63 | 0.001 | 1.58 | 0.044 |
| T2–T4 | 50 | 0.32 | 0.43 | 0.28 | 0.19 | 0.80 | ||||||
| N stage | N0 | 38 | 0.34 | 0.827 | 0.45 | 0.268 | 0.39 | 0.885 | 0.27 | 0.339 | 1.33 | 0.416 |
| N1–N3 | 36 | 0.39 | 0.58 | 0.70 | 0.34 | 0.80 | ||||||
| PR score | 6–8 | 27 | 0.37 | 0.271 | 0.50 | 0.889 | 0.32 | 0.527 | 0.19 | 0.317 | 0.72 | 0.037 |
| 0–5 | 47 | 0.48 | 0.47 | 0.59 | 0.35 | 1.48 | ||||||
| HER2 score | 2–3 | 43 | 0.57 | 0.424 | 0.55 | 0.316 | 0.78 | 0.029 | 0.47 | 0.003 | 1.46 | 0.019 |
| 1 | 31 | 0.34 | 0.50 | 0.32 | 0.18 | 0.66 | ||||||
| HER2 0 | ||||||||||||
| T stage | T1 | 40 | 0.48 | 0.610 | 0.57 | 0.522 | 0.51 | 0.156 | 0.15 | 0.540 | 0.66 | 0.926 |
| T2–T4 | 60 | 0.49 | 0.38 | 0.41 | 0.18 | 0.91 | ||||||
| N stage | N0 | 61 | 0.60 | 0.046 | 0.85 | 0.008 | 0.51 | 0.127 | 0.21 | 0.075 | 0.98 | 0.515 |
| N1–N3 | 39 | 0.34 | 0.30 | 0.41 | 0.13 | 0.66 | ||||||
| PR score | 6–8 | 47 | 0.41 | 0.197 | 0.41 | 0.884 | 0.42 | 0.710 | 0.13 | 0.219 | 0.85 | 0.875 |
| 0–5 | 53 | 0.54 | 0.42 | 0.45 | 0.18 | 0.65 | ||||||
| HER2 0, ER+/PR+ | ||||||||||||
| T stage | T1 | 31 | 0.45 | 0.706 | 0.47 | 0.800 | 0.41 | 0.138 | 0.20 | 0.771 | 0.69 | 0.962 |
| T2–T4 | 49 | 0.46 | 0.36 | 0.28 | 0.13 | 0.85 | ||||||
| N stage | N0 | 45 | 0.61 | 0.034 | 0.85 | 0.009 | 0.40 | 0.144 | 0.24 | 0.108 | 0.74 | 0.677 |
| N1–N3 | 35 | 0.32 | 0.20 | 0.27 | 0.11 | 0.75 | ||||||
| PR score | 6–8 | 47 | 0.41 | 0.342 | 0.41 | 0.504 | 0.42 | 0.504 | 0.13 | 0.550 | 0.85 | 0.355 |
| 0–5 | 33 | 0.52 | 0.35 | 0.28 | 0.19 | 0.64 | ||||||
| Characteristics | n | Relative Level * of miRNA and p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| miR-19b | p-Value | miR-22 | p-Value | miR-222 | p-Value | miR-378a | p-Value | miR-181a | p-Value | |||
| HER2 2+, 3+ | ||||||||||||
| T stage | T1 | 16 | 0.81 | 0.479 | 1.09 | 0.434 | 1.00 | 0.021 | 0.62 | 0.033 | 1.55 | 0.761 |
| T2–T4 | 27 | 0.37 | 0.44 | 0.29 | 0.25 | 1.20 | ||||||
| N stage | N0 | 27 | 0.33 | 0.161 | 0.45 | 0.199 | 0.68 | 0.372 | 0.41 | 0.118 | 1.40 | 0.736 |
| N1–N3 | 16 | 0.88 | 1.05 | 0.93 | 0.79 | 1.64 | ||||||
| HER2 0, 1+, ER+ and/or PR+ | ||||||||||||
| T stage | T1 | 48 | 0.48 | 0.379 | 0.60 | 0.745 | 0.51 | 0.094 | 0.16 | 0.993 | 0.73 | 0.610 |
| T2–T4 | 83 | 0.40 | 0.39 | 0.32 | 0.18 | 0.77 | ||||||
| N stage | N0 | 72 | 0.56 | 0.028 | 0.67 | 0.090 | 0.43 | 0.131 | 0.19 | 0.292 | 0.77 | 0.680 |
| N1–N3 | 59 | 0.31 | 0.36 | 0.34 | 0.15 | 0.66 | ||||||
| Characteristics | n | Relative Level * of miRNA and p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| miR-19b | p-Value | miR-22 | p-Value | miR-222 | p-Value | miR-378a | p-Value | miR-181a | p-Value | |||
| Ki-67 < 14% | ||||||||||||
| T stage | T1 | 26 | 0.81 | 0.334 | 0.87 | 0.665 | 0.54 | 0.199 | 0.27 | 0.696 | 0.92 | 0.966 |
| T2–T4 | 54 | 0.46 | 0.54 | 0.46 | 0.19 | 0.88 | ||||||
| N stage | N0 | 45 | 0.59 | 0.106 | 0.88 | 0.273 | 0.45 | 0.536 | 0.26 | 0.458 | 1.25 | 0.995 |
| N1–N3 | 35 | 0.42 | 0.58 | 0.46 | 0.19 | 0.83 | ||||||
| PR score | 6–8 | 46 | 0.50 | 0.738 | 0.75 | 0.325 | 0.62 | 0.319 | 0.20 | 0.458 | 0.93 | 0.300 |
| 0–5 | 34 | 0.51 | 0.44 | 0.38 | 0.25 | 0.78 | ||||||
| Ki-67 ≥ 14% | ||||||||||||
| T stage | T1 | 38 | 0.51 | 0.372 | 0.57 | 0.068 | 0.84 | 0.006 | 0.32 | 0.087 | 1.11 | 0.158 |
| T2–T4 | 56 | 0.31 | 0.30 | 0.28 | 0.16 | 0.77 | ||||||
| N stage | N0 | 54 | 0.45 | 0.539 | 0.49 | 0.539 | 0.48 | 0.443 | 0.21 | 0.780 | 1.15 | 0.228 |
| N1–N3 | 40 | 0.32 | 0.34 | 0.45 | 0.16 | 0.64 | ||||||
| PR score | 6–8 | 28 | 0.24 | 0.017 | 0.16 | 0.046 | 0.21 | 0.022 | 0.09 | 0.011 | 0.37 | 0.047 |
| 0–5 | 66 | 0.54 | 0.46 | 0.55 | 0.23 | 0.98 | ||||||
| Observed Change | Associated Tumor Characteristics | ||||
|---|---|---|---|---|---|
| ER+ and/or PR+ | ER− and PR− | HER2+ | HER2 0, ER+ and/or PR+ | Ki-67 ≥ 14% | |
| miR-19b decreased | N1–N3 stages | PR score 6–8 | |||
| miR-22 decreased | Ki-67 > 35% | N1–N3 stages | PR score 6–8 | ||
| miR-222 decreased | stages T2–T4 | Ki-67 > 35% | stages T2–T4 HER2 score 1 | T2–T4 stages PR score 6–8 | |
| miR-378a decreased | Ki-67 > 35% | stages T2–T4 HER2 score 1 | PR score 6–8 | ||
| miR-181a decreased | Ki-67 > 35% | HER2 score 1 | PR score 6–8 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalinina, T.; Kononchuk, V.; Alekseenok, E.; Obukhova, D.; Sidorov, S.; Strunkin, D.; Gulyaeva, L. Expression of Estrogen Receptor- and Progesterone Receptor-Regulating MicroRNAs in Breast Cancer. Genes 2021, 12, 582. https://doi.org/10.3390/genes12040582
Kalinina T, Kononchuk V, Alekseenok E, Obukhova D, Sidorov S, Strunkin D, Gulyaeva L. Expression of Estrogen Receptor- and Progesterone Receptor-Regulating MicroRNAs in Breast Cancer. Genes. 2021; 12(4):582. https://doi.org/10.3390/genes12040582
Chicago/Turabian StyleKalinina, Tatiana, Vladislav Kononchuk, Efim Alekseenok, Darya Obukhova, Sergey Sidorov, Dmitry Strunkin, and Lyudmila Gulyaeva. 2021. "Expression of Estrogen Receptor- and Progesterone Receptor-Regulating MicroRNAs in Breast Cancer" Genes 12, no. 4: 582. https://doi.org/10.3390/genes12040582
APA StyleKalinina, T., Kononchuk, V., Alekseenok, E., Obukhova, D., Sidorov, S., Strunkin, D., & Gulyaeva, L. (2021). Expression of Estrogen Receptor- and Progesterone Receptor-Regulating MicroRNAs in Breast Cancer. Genes, 12(4), 582. https://doi.org/10.3390/genes12040582