

Exploring the Diet-Gut Microbiota-Epigenetics Crosstalk Relevant to Neonatal Diabetes


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
3. An Overview of NDM
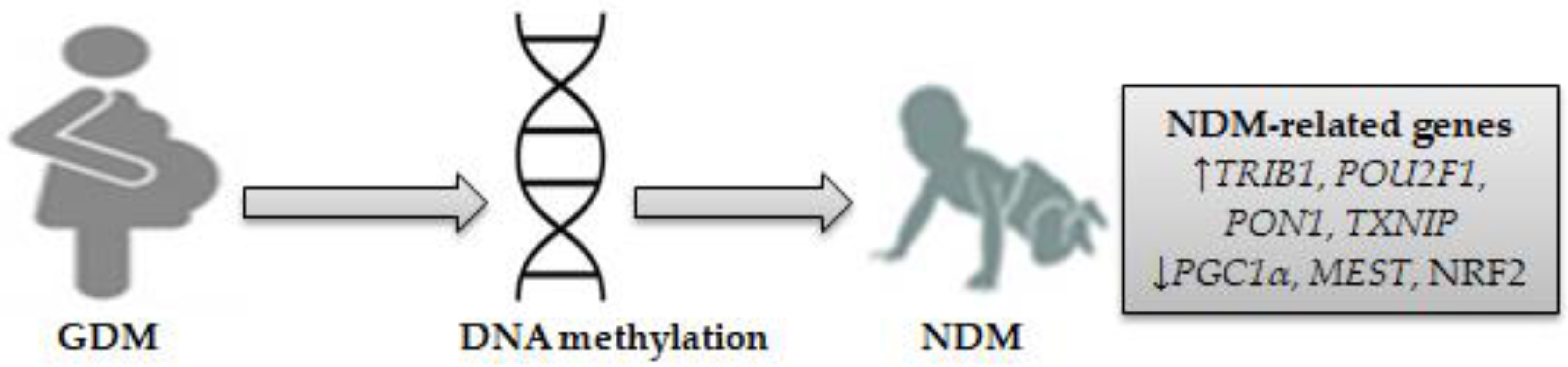
4. Epigenetic Modifications and NDM Gene Expression Profiles in Neonates Exposed to GDM
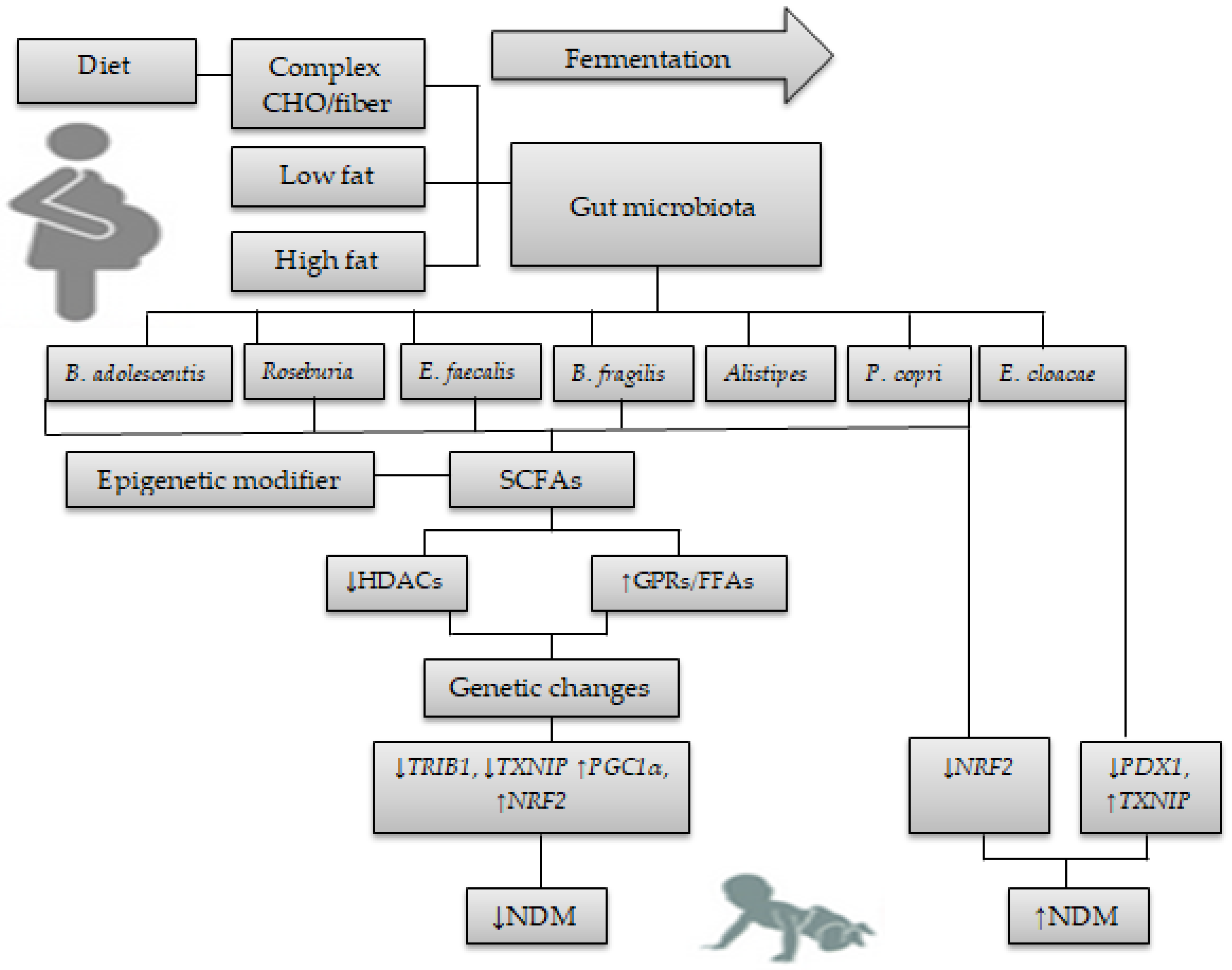
5. Diet, Gut Microbiota, and Epigenetic Crosstalk Alters Gene Expression in NDM
5.1. Diet Alters Gut Microbiota in GDM and NDM
5.2. Gut Microbiota and Its Metabolites Alter Gene Expression in NDM
5.2.1. Actinobacteria
5.2.2. Firnicutes
5.2.3. Bacteroidetes
5.2.4. Enterobacteriaceae
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABCC8 | ATP-binding cassette transporter subfamily C member 8 |
| cAMP | Cyclic AMP |
| CHO | Carbohydrate |
| COX-2 | Cyclooxygenase-2 |
| CPA | Carboxypeptidase A |
| CpG | Cytosine-phosphate-guanine dinucleotides |
| CRP | C-reactive peptide |
| CXCL | C-X-C motif chemokine ligand |
| DCs | Dendritic cells |
| DMGs | Differential methylation genes |
| DNA-PK | DNA-dependent protein kinase |
| EGR-1 | Early Growth Response Gene-1 |
| EIF2AK3 | Eukaryotic translation initiation factor 2-α kinase 3 |
| FFAs | Free fatty acid receptors |
| FoxO1 | Forkhead box O1 |
| FoxP3 | Forkhead box P3 |
| GATA6 | GATA binding protein 6 |
| GCK | Glucokinase |
| GDM | Gestational diabetes |
| GHs | Glycoside hydrolases |
| GLIS3 | GLI-similar 3 |
| GLP-1 | Glucagon-like peptide 1 |
| GPRs | Activate G-protein coupled receptors |
| H2O2 | Hydrogen peroxide |
| HbA1c | Hemoglobin A1c |
| HDACs | Histone deacetylases |
| HDL | High density lipoprotein |
| HLA | Human leukocyte antigen |
| HMOs | Human milk oligosaccharides |
| HOMA-IR | Homeostatic model assessment of insulin resistance |
| IECs | Intestinal epithelial cells |
| IGF | Insulin-like growth factor |
| IL | Interleukins |
| IFN-γ | Interferon-γ |
| lncRNA | Non-coding RNAs |
| INS | Preproinsulin |
| INSR | Insulin receptor |
| IR | Insulin resistance |
| JAK | Janus kinase |
| JNK | c-jun N-terminal kinase |
| KATP | ATP-sensitive potassium |
| KCNJ11 | Potassium inwardly rectifying channel, subfamily J, member 11 |
| LBP | LPS binding protein |
| LDL | Low density lipoprotein |
| LPS | Lipopolysaccharide |
| LRBA | LPS-responsive beige-like anchor |
| MAPK | Mitogen-activated protein kinase |
| MEST | Mesoderm-specific transcript |
| NDM | Neonatal diabetes |
| NF-kB | Nuclear factor-kappa B |
| NLRP3 | Nucleotide-binding oligomerization domain-like receptor family pyrin domain containing 3 |
| iNOS | Nitric oxide synthase |
| NRF2 | NF-E2 related factor 2 |
| OS | Oxidative stress |
| P53 | Protein 53 |
| PDX1 | Pancreatic and duodenal homeobox l |
| PGC1α | PPAR-γ coactivator-1-α |
| PI3K | Phosphatidylinositol-3 kinase |
| PKA PLAG1 | Protein kinase A PLAG1 like zinc finger 1 |
| PNDM | Permanent neonatal diabetes |
| PON1 | Paraoxonase 1 |
| POU2F1 PPP1R13L | POU class 2 homeobox 1 protein phosphatase 1 regulatory subunit 13 like |
| PUFA | Polyunsaturated fatty acid |
| ROS | Reactive oxygen species |
| SCFAs | Short-chain fatty acids |
| SFA | Saturated fatty acid |
| SLC2A2 | Solute carrier family 2 member 2 |
| SLC19A2 | Solute carrier family 19, member 2 |
| SOCS3 | Suppressor of cytokine signaling 3 |
| SRS | Silver-Russell syndrome |
| Srx-1 | Sulfiredoxin-1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| T1D | Type 1 diabetes |
| T2D | Type 2 diabetes |
| TGF-β | Transforming growth factor-β |
| TJ | Tight junction |
| TLR | Toll like receptor |
| TNDM | Transient neonatal diabetes |
| TNF-α | Tumor necrosis factor-α |
| Treg | Regulatory T |
| TRIB1 | Tribbles homolog 1 |
| TXN | Thioredoxin |
| TXNIP | Thioredoxin interacting protein |
| VEGF | Vascular endothelial growth factor |
| VIPR1 | Vasoactive intestinal peptide receptor |
| VLCKD | Very low-calorie ketogenic diet |
| ZEP57 | Zinc finger protein 57 homolog |
References
- Edghill, E.L.; Dix, R.J.; Flanagan, S.E.; Bingley, P.J.; Hattersley, A.T.; Ellard, S.; Gillespie, K.M. HLA genotyping supports a nonautoimmune etiology in patients diagnosed with diabetes under the age of 6 months. Diabetes 2006, 55, 1895–1898. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Cabezas, O.; Ellard, S. Diabetes mellitus in neonates and infants: Genetic heterogeneity, clinical approach to diagnosis, and therapeutic options. Horm. Res. Paediatr. 2013, 80, 137–146. [Google Scholar] [CrossRef]
- Flanagan, S.E.; Haapaniemi, E.; Russell, M.A.; Caswell, R.; Allen, H.L.; De Franco, E.; McDonald, T.J.; Rajala, H.; Ramelius, A.; Barton, J.; et al. Activating germline mutations in STAT3 cause early-onset multi-organ autoimmune disease. Nat. Genet. 2014, 46, 812–814. [Google Scholar] [CrossRef]
- Johnson, M.B.; De Franco, E.; Allen, H.L.; Senani, A.A.; Elbarbary, N.; Siklar, Z.; Berberoglu, M.; Imane, Z.; Haghighi, A.; Razavi, Z.; et al. Recessively inherited LRBA mutations cause autoimmunity presenting as neonatal diabetes. Diabetes 2017, 66, 2316–2322. [Google Scholar] [CrossRef] [PubMed]
- Nishimaki, S.; Yukawa, T.; Makita, Y.; Honda, H.; Kikuchi, N.; Minamisawa, S.; Yokota, S. Transient neonatal diabetes mellitus in extremely preterm infant. Arch. Dis. Child.-Fetal Neonatal Ed. 2008, 93, F240–F241. [Google Scholar] [CrossRef]
- Busiah, K.; Auger, J.; Fauret-Amsellem, A.-L.; Dahan, S.; Pouvreau, N.; Cavé, H.; Polak, M.; Mitanchez, D. Differentiating transient idiopathic hyperglycaemia and neonatal diabetes mellitus in preterm infants. Horm. Res. Paediatr. 2015, 84, 68–72. [Google Scholar] [CrossRef]
- Siklar, Z.; Ellard, S.; Okulu, E.; Berberoğlu, M.; Young, E.; Erdeve, S.S.; Mungan, I.A.; Hacihamdioğlu, B.; Erdeve, O.; Arsan, S.; et al. Transient neonatal diabetes with two novel mutations in the KCNJ11 gene and response to sulfonylurea treatment in a preterm infant. J. Pediatr. Endocrinol. Metab. 2011, 24, 1077–1080. [Google Scholar] [CrossRef]
- Besser, R.E.J.; Flanagan, S.E.; Mackay, D.G.J.; Temple, I.K.; Shepherd, M.H.; Shields, B.M.; Ellard, S.; Hattersley, A.T. Prematurity and genetic testing for neonatal diabetes. Pediatrics 2016, 138, e20153926. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, I.; Iqbal, F.; Thorne, R.F. Editorial: Relationship between gestational and neonatal diabetes mellitus. Front. Endocrinol. 2022, 13, 1060147. [Google Scholar] [CrossRef]
- Perez-Muñoz, M.E.; Arrieta, M.C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.W.; Clemente, J.C.; Peter, I.; Loos, R.J.F. The prenatal gut microbiome: Are we colonized with bacteria in utero? Pediatr. Obes. 2017, 12, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Senn, V.; Bassler, D.; Choudhury, R.; Scholkmann, F.; Righini-Grunder, F.; Vuille-dit-Bille, R.N.; Restin, T. Microbial colonization from the fetus to early childhood—A comprehensive review. Front. Cell Infect. Microbiol. 2020, 10, 573735. [Google Scholar] [CrossRef] [PubMed]
- Alsharairi, N.A. The infant gut microbiota and risk of asthma: The effect of maternal nutrition during pregnancy and lactation. Microorganisms 2020, 8, 1119. [Google Scholar] [CrossRef]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef]
- Nuriel-Ohayon, M.; Neuman, H.; Koren, O. Microbial changes during pregnancy, birth, and infancy. Front. Microbiol. 2016, 7, 1031. [Google Scholar] [CrossRef]
- Jeong, S. Factors influencing development of the infant microbiota: From prenatal period to early infancy. Clin. Exp. Pediatr. 2022, 65, 438–447. [Google Scholar] [CrossRef]
- Alsharairi, N.A. The role of short-chain fatty acids in the interplay between a very low-calorie ketogenic diet and the infant gut microbiota and its therapeutic implications for reducing asthma. Int. J. Mol. Sci. 2020, 21, 9580. [Google Scholar] [CrossRef]
- Alsharairi, N.A. The role of short-chain fatty acids in mediating very low-calorie ketogenic diet-infant gut microbiota relationships and its therapeutic potential in obesity. Nutrients 2021, 13, 3702. [Google Scholar] [CrossRef]
- Alsharairi, N.A. The therapeutic role of short-chain fatty acids mediated very low-calorie ketogenic diet-gut microbiota relationships in paediatric inflammatory bowel diseases. Nutrients 2022, 14, 4113. [Google Scholar] [CrossRef] [PubMed]
- Alsharairi, N.A. Therapeutic potential of gut microbiota and its metabolite short-chain fatty acids in neonatal necrotizing enterocolitis. Life 2023, 13, 561. [Google Scholar] [CrossRef]
- Hu, J.; Nomura, Y.; Bashir, A.; Fernandez-Hernandez, H.; Itzkowitz, S.; Pei, Z.; Stone, J.; Loudon, H.; Peter, I. Diversified microbiota of meconium is affected by maternal diabetes status. PLoS ONE 2013, 8, e78257. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Nie, Y.; Shao, R.; Duan, S.; Jiang, Y.; Wang, M.; Xing, Z.; Sun, Q.; Liu, X.; Xu, W. Diversified gut microbiota in newborns of mothers with gestational diabetes mellitus. PLoS ONE 2018, 13, e0205695. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zheng, J.; Shi, W.; Du, N.; Xu, X.; Zhang, Y.; Ji, P.; Zhang, F.; Jia, Z.; Wang, Y.; et al. Dysbiosis of maternal and neonatal microbiota associated with gestational diabetes mellitus. Gut 2018, 67, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Hasain, Z.; Mokhtar, N.M.; Kamaruddin, N.A.; Ismail, N.A.M.; Razalli, N.H.; Gnanou, J.V.; Ali, R.A.R. Gut microbiota and gestational diabetes mellitus: A review of host-gut microbiota interactions and their therapeutic potential. Front. Cell Infect. Microbiol. 2020, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Qin, Y.; Chen, M.; Zhang, Y.; Wang, X.; Dong, T.; Chen, G.; Sun, X.; Lu, T.; White, R.A.; et al. Gestational diabetes mellitus is associated with the neonatal gut microbiota and metabolome. BMC Med. 2021, 19, 120. [Google Scholar] [CrossRef]
- Ferrocino, I.; Ponzo, V.; Gambino, R.; Zarovska, A.; Leone, F.; Monzeglio, C.; Goitre, I.; Rosato, R.; Romano, A.; Grassi, G.; et al. Changes in the gut microbiota composition during pregnancy in patients with gestational diabetes mellitus (GDM). Sci. Rep. 2018, 8, 12216. [Google Scholar] [CrossRef]
- Sugino, K.Y.; Hernandez, T.L.; Barbour, L.A.; Kofonow, J.M.; Frank, D.N.; Friedman, J.E. A maternal higher-complex carbohydrate diet increases bifidobacteria and alters early life acquisition of the infant microbiome in women with gestational diabetes mellitus. Front. Endocrinol. 2022, 13, 921464. [Google Scholar] [CrossRef]
- Kocova, M. Genetic spectrum of neonatal diabetes. Balkan J. Med. Genet. 2020, 23, 5–15. [Google Scholar] [CrossRef]
- Greeley, S.A.W.; Tucker, S.E.; Naylor, R.N.; Bell, G.I.; Philipson, L.H. Neonatal diabetes mellitus: A model for personalized medicine. Trends Endocrinol. Metab. 2010, 21, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Greeley, S.A.W.; Naylor, R.N.; Philipson, L.H.; Bell, G.I. Neonatal diabetes: An expanding list of genes allows for improved diagnosis and treatment. Curr. Diabetes Rep. 2011, 11, 519–532. [Google Scholar] [CrossRef]
- Naylor, R.N.; Greeley, S.A.W.; Bell, G.I.; Philipson, L.H. Genetics and pathophysiology of neonatal diabetes mellitus. J. Diabetes Investig. 2011, 2, 158–169. [Google Scholar] [CrossRef]
- Vaxillaire, M.; Bonnefond, A.; Froguel, P. The lessons of early-onset monogenic diabetes for the understanding of diabetes pathogenesis. Best. Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Zübarioğlu, A.U.; Bülbül, A.; Uslu, H.S. Neonatal diabetes mellitus. Sisli Etfal Hastan. Tip Bülteni 2018, 52, 71–78. [Google Scholar] [CrossRef]
- Dahl, A.; Kumar, S. Recent Advances in Neonatal Diabetes. Diabetes Metab. Syndr. Obes. 2020, 13, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Colclough, K.; Gloyn, A.L.; Pollin, T.I. Monogenic diabetes: A gateway to precision medicine in diabetes. J. Clin. Investig. 2021, 131, e142244. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.; Boonen, S.E.; Dahl, C.; Hahnemann, J.M.D.; Mackay, D.J.D.G.; Tümer, Z.; Grønskov, K.; Temple, I.K.; Guldberg, P.; Tommerup, N. Genome-wide DNA methylation analysis of transient neonatal diabetes type 1 patients with mutations in ZFP57. BMC Med. Genet. 2016, 17, 29. [Google Scholar] [CrossRef] [PubMed]
- Mackay, D.J.G.; Temple, I.K. Transient neonatal diabetes mellitus type 1. Am. J. Med. Genet. C Semin. Med. Genet. 2010, 154C, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Akerman, I.; Maestro, M.A.; De Franco, E.; Grau, V.; Flanagan, S.; García-Hurtado, J.; Mittler, G.; Ravassard, P.; Piemonti, L.; Ellard, S.; et al. Neonatal diabetes mutations disrupt a chromatin pioneering function that activates the human insulin gene. Cell Rep. 2021, 35, 108981. [Google Scholar] [CrossRef]
- Hammoud, B.; Greeley, S.A.W. Growth and development in monogenic forms of neonatal diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2022, 29, 65–77. [Google Scholar] [CrossRef]
- Johnson, M.B.; Patel, K.A.; De Franco, E.; Hagopian, W.; Killian, M.; McDonald, T.J.; Tree, T.I.M.; Domingo-Vila, C.; Hudson, M.; Hammersley, S.; et al. Type 1 diabetes can present before the age of 6 months and is characterized by autoimmunity and rapid loss of beta cells. Diabetologia 2020, 63, 2605–2615. [Google Scholar] [CrossRef]
- Yan, J.; Yang, H. Gestational diabetes mellitus, programing and epigenetics. J. Matern. Fetal Neonatal Med. 2014, 27, 1266–1269. [Google Scholar] [CrossRef] [PubMed]
- Hjort, L.; Martino, D.; Grunnet, L.G.; Naeem, H.; Maksimovic, J.; Olsson, A.H.; Zhang, C.; Ling, C.; Olsen, S.F.; Saffery, R.; et al. Gestational diabetes and maternal obesity are associated with epigenome-wide methylation changes in children. JCI Insight 2018, 3, e122572. [Google Scholar] [CrossRef] [PubMed]
- Hjort, L.; Novakovic, B.; Grunnet, L.G.; Maple-Brown, L.; Damm, P.; Desoye, G.; Saffery, R. Diabetes in pregnancy and epigenetic mechanisms-how the first 9 months from conception might affect the child’s epigenome and later risk of disease. Lancet Diabetes Endocrinol. 2019, 7, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Kakongoma, N.; Hua, W.; Xu, J.; Wang, Y.; He, S.; Gu, H.; Shi, J.; Hu, W. DNA methylation and expression profiles of placenta and umbilical cord blood reveal the characteristics of gestational diabetes mellitus patients and offspring. Clin. Epigenetics 2022, 14, 69. [Google Scholar] [CrossRef] [PubMed]
- Ott, R.; Stupin, J.H.; Melchior, K.; Schellong, K.; Ziska, T.; Dudenhausen, J.W.; Henrich, W.; Rancourt, R.C.; Plagemann, A. Alterations of adiponectin gene expression and DNA methylation in adipose tissues and blood cells are associated with gestational diabetes and neonatal outcome. Clin. Epigenetics 2018, 10, 131. [Google Scholar] [CrossRef]
- Kang, J.; Lee, C.-N.; Li, H.-Y.; Hsu, K.-H.; Lin, S.-Y. Genome-wide DNA methylation variation in maternal and cord blood of gestational diabetes population. Diabetes Res. Clin. Pract. 2017, 132, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Ruchat, S.-M.; Houde, A.-A.; Voisin, G.; St-Pierre, J.; Perron, P.; Baillargeon, J.-P.; Gaudet, D.; Hivert, M.-F.; Brisson, D.; Bouchard, L. Gestational diabetes mellitus epigenetically affects genes predominantly involved in metabolic diseases. Epigenetics 2013, 9, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Winzell, M.S.; Ahrén, B. Role of VIP and PACAP in islet function. Peptides 2007, 28, 1805–1813. [Google Scholar] [CrossRef]
- Soubeyrand, S.; Martinuk, A.; Naing, T.; Lau, P.; McPherson, R. Role of Tribbles Pseudokinase 1 (TRIB1) in human hepatocyte metabolism. Biochim. Biophys. Acta 2016, 1862, 223–232. [Google Scholar] [CrossRef]
- Popova, P.V.; Vasileva, L.B.; Tkachuk, A.S.; Puzanov, M.V.; Bolotko, Y.A.; Pustozerov, E.A.; Gerasimov, A.S.; Zazerskaya, I.E.; Li, O.A.; Vasilyeva, E.Y.; et al. Association of tribbles homologue 1 gene expression in human umbilical vein endothelial cells with duration of intrauterine exposure to hyperglycaemia. Genet. Res. 2018, 100, e3. [Google Scholar] [CrossRef]
- Sukarieh, R.; Joseph, R.; Leow, S.C.; Li, Y.; Löffler, M.; Aris, I.M.; Tan, J.H.; Teh, A.L.; Chen, L.; Holbrook, J.D.; et al. Molecular pathways reflecting poor intrauterine growth are found in Wharton’s jelly-derived mesenchymal stem cells. Hum. Reprod. 2014, 29, 2287–2301. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liang, R.; Liu, T.; Wang, L.; Zou, J.; Liu, N.; Liu, Y.; Cai, X.; Liu, Y.; Ding, X.; et al. Opposing effects of IL-1β/COX-2/PGE2 pathway loop on islets in type 2 diabetes mellitus. Endocr. J. 2019, 66, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Dong, S.; Wu, L.; Bai, Y.; Bi, X.; Li, Y.; Shu, C. Maternal and placental DNA methylation changes associated with the pathogenesis of gestational diabetes mellitus. Nutrients 2023, 15, 70. [Google Scholar] [CrossRef]
- Hjort, L.; Novakovic, B.; Cvitic, S.; Saffery, R.; Damm, P.; Desoye, G. Placental DNA methylation in pregnancies complicated by maternal diabetes and/or obesity: State of the art and research gaps. Epigenetics 2022, 17, 2188–2208. [Google Scholar] [CrossRef]
- Xie, X.; Gao, H.; Zeng, W.; Chen, S.; Feng, L.; Deng, D.; Qiao, F.-Y.; Liao, L.; McCormick, K.; Ning, Q.; et al. Placental DNA methylation of peroxisome-proliferator-activated receptor-γ co-activator-1α promoter is associated with maternal gestational glucose level. Clin. Sci. 2015, 129, 385–394. [Google Scholar] [CrossRef]
- Ruschke, K.; Fishbein, L.; Dietrich, A.; Klöting, N.; Tönjes, A.; Oberbach, A.; Fasshauer, M.; Jenkner, J.; Schön, M.R.; Stumvoll, M.; et al. Gene expression of PPARgamma and PGC-1alpha in human omental and subcutaneous adipose tissues is related to insulin resistance markers and mediates beneficial effects of physical training. Eur. J. Endocrinol. 2010, 162, 515–523. [Google Scholar] [CrossRef]
- Côté, S.; Gagné-Ouellet, V.; Guay, S.-P.; Allard, C.; Houde, A.-A.; Perron, P.; Baillargeon, J.-P.; Gaudet, D.; Guérin, R.; Brisson, D.; et al. PPARGC1α gene DNA methylation variations in human placenta mediate the link between maternal hyperglycemia and leptin levels in newborns. Clin. Epigenetics 2016, 8, 72. [Google Scholar] [CrossRef]
- Gao, Y.; She, R.; Sha, W. Gestational diabetes mellitus is associated with decreased adipose and placenta peroxisome proliferator-activator receptor γ expression in a Chinese population. Oncotarget 2017, 8, 113928–113937. [Google Scholar] [CrossRef]
- Wang, L.; Fan, H.; Zhou, L.; Wu, Y.; Lu, H.; Luo, J. Altered expression of PGC-1α and PDX1 and their methylation status are associated with fetal glucose metabolism in gestational diabetes mellitus. Biochem. Biophys. Res. Commun. 2018, 501, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Higazy, R.; Gauda, E.B. PGC-1α activity and mitochondrial dysfunction in preterm infants. Front. Physiol. 2022, 13, 997619. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Leahy, A.A.; Monga, N.; Peshavaria, M.; Jetton, T.L.; Leahy, J.L. Peroxisome proliferator-activated receptor γ (PPARγ) and its target genes are downstream effectors of FoxO1 protein in islet β-cells: Mechanism of β-cell compensation and failure. J. Biol. Chem. 2013, 288, 25440–25449. [Google Scholar] [CrossRef]
- El Hajj, N.; Pliushch, G.; Schneider, E.; Dittrich, M.; Müller, T.; Korenkov, M.; Aretz, M.; Zechner, U.; Lehnen, H.; Haaf, T. Metabolic programming of MEST DNA methylation by intrauterine exposure to gestational diabetes mellitus. Diabetes 2013, 62, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Weng, X.; Liu, F.; Zhang, H.; Kan, M.; Wang, T.; Dong, M.; Liu, Y. Genome-wide DNA methylation profiling in infants born to gestational diabetes mellitus. Diabetes Res. Clin. Pract. 2018, 142, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Bentley, L.; Nakabayashi, K.; Monk, D.; Beechey, C.; Peters, J.; Birjandi, Z.; Khayat, F.E.; Patel, M.; Preece, M.A.; Stanier, P.; et al. The imprinted region on human chromosome 7q32 extends to the carboxypeptidase A gene cluster: An imprinted candidate for Silver-Russell syndrome. J. Med. Genet. 2003, 40, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Sparago, A.; Cerrato, F.; Riccio, A. Is ZFP57 binding to H19/IGF2:IG-DMR affected in Silver-Russell syndrome? Clin. Epigenetics 2018, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Brereton, R.E.; Nickerson, S.L.; Woodward, K.J.; Edwards, T.; Sivamoorthy, S.; Walters, F.R.V.; Chabros, V.; Marchin, V.; Grumball, T.; Kennedy, D.; et al. Further heterogeneity in Silver-Russell syndrome: PLAG1 deletion in association with a complex chromosomal rearrangement. Am. J. Med. Genet. A 2021, 185, 3136–3145. [Google Scholar] [CrossRef] [PubMed]
- Quilter, C.R.; Cooper, W.N.; Cliffe, K.M.; Skinner, B.M.; Prentice, P.M.; Nelson, L.; Bauer, J.; Ong, K.K.; Constância, M.; Lowe, W.L.; et al. Impact on offspring methylation patterns of maternal gestational diabetes mellitus and intrauterine growth restraint suggest common genes and pathways linked to subsequent type 2 diabetes risk. FASEB J. 2014, 28, 4868–4879. [Google Scholar] [CrossRef]
- Ng, M.C.Y.; Lam, V.K.L.; Tam, C.H.T.; Chan, A.W.H.; So, W.-Y.; Ma, R.C.W.; Zee, B.C.Y.; Waye, M.M.Y.; Mak, W.W.; Hu, C.; et al. Association of the POU class 2 homeobox 1 gene (POU2F1) with susceptibility to Type 2 diabetes in Chinese populations. Diabet. Med. 2010, 27, 1443–1449. [Google Scholar] [CrossRef]
- Wang, P.; Jin, T. Hydrogen peroxide stimulates nuclear import of the POU homeodomain protein Oct-1 and its repressive effect on the expression of Cdx-2. BMC Cell Biol. 2010, 11, 56. [Google Scholar] [CrossRef]
- Primo-Parmo, S.L.; Sorenson, R.C.; Teiber, J.; Du, B.N.L. The human serum paraoxonase/arylesterase gene (PON1) is one member of a multigene family. Genomics 1996, 33, 498–507. [Google Scholar] [CrossRef]
- León, I.; Roldán, S.H.; José Rodrigo, M.; Rodríguez, M.L.; Fisher, J.; Mitchell, C.; Lage-Castellanos, A. The shared mother-child epigenetic signature of neglect is related to maternal adverse events. Front. Physiol. 2022, 13, 966740. [Google Scholar] [CrossRef]
- Pappa, K.I.; Gazouli, M.; Anastasiou, E.; Loutradis, D.; Anagnou, N.P. The Q192R polymorphism of the paraoxonase-1 (PON1) gene is associated with susceptibility to gestational diabetes mellitus in the Greek population. Gynecol. Endocrinol. 2017, 33, 617–620. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.-H.; Liu, Q.-Q.; Chen, M.; Bai, H.; Jiang, C.-Y.; Guan, L.-B.; Fan, P. Lactonase activity and status of paraoxonase 1 and oxidative stress in neonates of women with gestational diabetes mellitus. Pediatr. Res. 2021, 89, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Numaguchi, M.; Caccavello, R.; Kimura, S. Paraoxonase 1 lactonase activity and distribution in the HDL subclasses in the cord blood. Redox Rep. 2014, 19, 124–132. [Google Scholar] [CrossRef]
- Cheng, X.; Siow, R.C.M.; Mann, G.E. Impaired redox signaling and antioxidant gene expression in endothelial cells in diabetes: A role for mitochondria and the nuclear factor-E2-related factor 2-Kelch-like ECH-associated protein 1 defense pathway. Antioxid. Redox Signal. 2011, 14, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Chapple, S.J.; Patel, B.; Puszyk, W.; Sugden, D.; Yin, X.; Mayr, M.; Siow, R.C.M.; Mann, G.E. Gestational diabetes mellitus impairs Nrf2-mediated adaptive antioxidant defenses and redox signaling in fetal endothelial cells in utero. Diabetes 2013, 62, 4088–4097. [Google Scholar] [CrossRef] [PubMed]
- Baumel-Alterzon, S.; Scott, D.K. Regulation of Pdx1 by oxidative stress and Nrf2 in pancreatic beta-cells. Front. Endocrinol. 2022, 13, 1011187. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.W.; Juvinao-Quintero, D.L.; Ronkainen, J.; Ott, R.; Alfano, R.; Canouil, M.; Geurtsen, M.L.; Khamis, A.; Küpers, L.K.; Lim, I.Y.; et al. Maternal glycemic dysregulation during pregnancy and neonatal blood DNA methylation: Meta-analyses of epigenome-wide association studies. Diabetes Care 2022, 45, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Wondafrash, D.Z.; Nire’a, A.T.; Tafere, G.G.; Desta, D.M.; Berhe, D.A.; Zewdie, K.A. Thioredoxin-interacting protein as a novel potential therapeutic target in diabetes mellitus and its underlying complications. Diabetes Metab. Syndr. Obes. 2020, 13, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Basnet, R.; Basnet, T.B.; Basnet, B.B.; Khadka, S. Overview on thioredoxin-interacting protein (TXNIP): A potential target for diabetes intervention. Curr. Drug. Targets 2022, 23, 761–767. [Google Scholar] [CrossRef]
- Sarina; Li, D.F.; Feng, Z.Q.; Du, J.; Zhao, W.H.; Huang, N.; Jia, J.C.; Wu, Z.Y.; Alamusi; Wang, Y.Y.; et al. Mechanism of placenta damage in gestational diabetes mellitus by investigating TXNIP of patient samples and gene functional research in cell line. Diabetes Ther. 2019, 10, 2265–2288. [Google Scholar] [CrossRef]
- Pasternak, Y.; Ohana, M.; Biron-Shental, T.; Cohen-Hagai, K.; Benchetrit, S.; Zitman-Gal, T. Thioredoxin, thioredoxin interacting protein and transducer and activator of transcription 3 in gestational diabetes. Mol. Biol. Rep. 2020, 47, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Velayos, T.; Martínez, R.; Alonso, M.; Garcia-Etxebarria, K.; Aguayo, A.; Camarero, C.; Urrutia, I.; Martínez de LaPiscina, I.; Barrio, R.; Santin, I.; et al. An activating mutation in STAT3 results in neonatal diabetes through reduced insulin synthesis. Diabetes 2017, 66, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, P.; Metos, J.; Babu, P.V.A. Impact of type 1 diabetes on the composition and functional potential of gut microbiome in children and adolescents: Possible mechanisms, current knowledge, and challenges. Gut Microbes 2021, 13, 1926841. [Google Scholar] [CrossRef] [PubMed]
- Del Chierico, F.; Rapini, N.; Deodati, A.; Matteoli, M.C.; Cianfarani, S.; Putignani, L. Pathophysiology of type 1 diabetes and gut microbiota role. Int. J. Mol. Sci. 2022, 23, 14650. [Google Scholar] [CrossRef]
- Yang, Q.; Ouyang, J.; Sun, F.; Yang, J. Short-chain fatty acids: A soldier fighting against inflammation and protecting from tumorigenesis in people with diabetes. Front. Immunol. 2020, 11, 590685. [Google Scholar] [CrossRef]
- Yi, C.; Sun, W.; Ding, L.; Yan, M.; Sun, C.; Qiu, C.; Wang, D.; Wu, L. Short-chain fatty acids weaken ox-LDL-induced cell inflammatory injury by inhibiting the NLRP3/Caspase-1 pathway and affecting cellular metabolism in THP-1 cells. Molecules 2022, 27, 8801. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, Y.; Wang, P.; Huang, Y.; Wang, F. Short-chain fatty acids manifest stimulative and protective effects on intestinal barrier function through the inhibition of NLRP3 inflammasome and autophagy. Cell Physiol. Biochem. 2018, 49, 190–205. [Google Scholar] [CrossRef]
- Yaku, K.; Enami, Y.; Kurajyo, C.; Matsui-Yuasa, I.; Konishi, Y.; Kojima-Yuasa, A. The enhancement of phase 2 enzyme activities by sodium butyrate in normal intestinal epithelial cells is associated with Nrf2 and p53. Mol. Cell Biochem. 2012, 370, 7–14. [Google Scholar] [CrossRef]
- Milani, C.; Mancabelli, L.; Lugli, G.A.; Duranti, S.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Ferretti, P.; Gorfer, V.; et al. Exploring vertical transmission of Bifidobacteria from mother to child. Appl. Environ. Microbiol. 2015, 81, 7078–7087. [Google Scholar] [CrossRef]
- Asnicar, F.; Manara, F.; Zolfo, M.; Truong, D.T.; Scholz, M.; Armanini, F.; Ferretti, P.; Gorfer, V.; Pedrotti, A.; Tett, A.; et al. Studying vertical microbiome transmission from mothers to infants by strain-level metagenomic profiling. mSystems 2017, 2, e00164-16. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, P.; Pasolli, E.; Tett, A.; Asnicar, F.; Gorfer, V.; Fedi, S.; Armanini, F.; Truong, D.T.; Manara, S.; Zolfo, M.; et al. Mother-to-infant microbial transmission from different body sites shapes the developing infant gut microbiome. Cell Host Microbe. 2018, 24, 133–145.e5. [Google Scholar] [CrossRef]
- Koo, H.; McFarland, B.C.; Hakim, J.A.; Crossman, D.K.; Crowley, M.R.; Rodriguez, J.M.; Benveniste, E.N.; Morrow, C.D. An individualized mosaic of maternal microbial strains is transmitted to the infant gut microbial community. R. Soc. Open. Sci. 2020, 7, 192200. [Google Scholar] [CrossRef] [PubMed]
- Ponzo, V.; Ferrocino, I.; Zarovska, A.; Amenta, M.B.; Leone, F.; Monzeglio, C.; Rosato, R.; Pellegrini, M.; Gambino, R.; Cassader, M.; et al. The microbiota composition of the offspring of patients with gestational diabetes mellitus (GDM). PLoS ONE 2019, 14, e0226545. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.-S.; Lu, J.-H.; Li, S.-H.; Li, J.-H.; Yuan, M.-Y.; He, J.-R.; Chen, N.-N.; Xiao, W.-Q.; Shen, S.-Y.; Qiu, L.; et al. Connections between the human gut microbiome and gestational diabetes mellitus. Gigascience 2017, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Soderborg, T.K.; Carpenter, C.M.; Janssen, R.C.; Weir, T.L.; Robertson, C.E.; Ir, D.; Young, B.E.; Krebs, N.F.; Hernandez, T.L.; Barbour, L.A.; et al. Gestational diabetes is uniquely associated with altered early seeding of the infant gut microbiota. Front. Endocrinol. 2020, 11, 603021. [Google Scholar] [CrossRef]
- Gopalsamy, G.; Mortimer, E.; Greenfield, P.; Bird, A.R.; Young, G.P.; Christophersen, C.T. Resistant starch is actively fermented by infant faecal microbiota and increases microbial diversity. Nutrients 2019, 11, 1345. [Google Scholar] [CrossRef]
- Nurmi, J.T.; Puolakkainen, P.A.; Rautonen, N.E. Bifidobacterium lactis sp. 420 up-regulates cyclooxygenase (Cox)-1 and down-regulates Cox-2 gene expression in a Caco-2 cell culture model. Nutr. Cancer 2005, 51, 83–892. [Google Scholar] [CrossRef]
- Kawabata, K.; Baba, N.; Sakano, T.; Hamano, Y.; Taira, S.; Tamura, A.; Baba, S.; Natsume, M.; Ishii, T.; Murakami, S.; et al. Functional properties of anti-inflammatory substances from quercetin-treated Bifidobacterium adolescentis. Biosci. Biotechnol. Biochem. 2018, 82, 689–697. [Google Scholar] [CrossRef]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Rochat, F.; Chassard, C. Vertical mother-neonate transfer of maternal gut bacteria via breastfeeding. Environ. Microbiol. 2014, 16, 2891–2904. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yu, D.; Wang, Y.; Yuan, H.; Ning, X.; Rui, B.; Lei, Z.; Yuan, J.; Yan, J.; Li, M. The intestinal dysbiosis of mothers with gestational diabetes mellitus (GDM) and its impact on the gut microbiota of their newborns. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 3044534. [Google Scholar] [CrossRef] [PubMed]
- Arif, S.; Moore, F.; Marks, K.; Bouckenooghe, T.; Dayan, C.M.; Planas, R.; Vives-Pi, M.; Powrie, J.; Tree, T.; Marchetti, O.; et al. Peripheral and islet interleukin-17 pathway activation characterizes human autoimmune diabetes and promotes cytokine-mediated β-cell death. Diabetes 2011, 60, 2112–2119. [Google Scholar] [CrossRef] [PubMed]
- Nepelska, M.; de Wouters, T.; Jacouton, E.; Béguet-Crespel, F.; Lapaque, N.; Doré, J.; Arulampalam, V.; Blottière, H.M. Commensal gut bacteria modulate phosphorylation-dependent PPARγ transcriptional activity in human intestinal epithelial cells. Sci. Rep. 2017, 7, 43199. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, A.; Annapure, U.S. Maternal-neonatal transmission of Enterococcus strains during delivery. Beni-Suef Univ. J. Basic. Appl. Sci. 2019, 8, 25. [Google Scholar] [CrossRef]
- Vitetta, L.; Coulson, S.; Thomsen, M.; Nguyen, T.; Hall, S. Probiotics D-lactic acidosis, oxidative stress and strain specificity. Gut Microbes 2017, 8, 311–322. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Ishii, M.; Hasegawa, S.; Sekimizu, K. Enterococcus faecalis YM0831 suppresses sucrose-induced hyperglycemia in a silkworm model and in humans. Commun. Biol. 2019, 2, 157. [Google Scholar] [CrossRef]
- Are, A.; Aronsson, L.; Wang, S.; Greicius, G.; Lee, Y.K.; Gustafsson, J.-A.; Pettersson, S.; Arulampalam, V. Enterococcus faecalis from newborn babies regulate endogenous PPARgamma activity and IL-10 levels in colonic epithelial cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1943–1948. [Google Scholar] [CrossRef]
- Maqsood, R.; Rodriguez, R.R.C.; Handley, S.A.; Ndao, I.M.; Tarr, P.I.; Warner, B.B.; Lim, E.S.; Holtz, L.R. Discordant transmission of bacteria and viruses from mothers to babies at birth. Microbiome 2019, 7, 156. [Google Scholar] [CrossRef]
- Chen, F.; Gan, Y.; Li, Y.; He, W.; Wu, W.; Wang, K.; Li, Q. Association of gestational diabetes mellitus with changes in gut microbiota composition at the species level. BMC Microbiol. 2021, 21, 147. [Google Scholar] [CrossRef]
- Su, Y.; Wang, H.K.; Gan, X.P.; Chen, L.; Cao, Y.N.; Cheng, D.C.; Zhang, D.Y.; Liu, W.Y.; Li, F.F.; Xu, X.M. Alterations of gut microbiota in gestational diabetes patients during the second trimester of pregnancy in the Shanghai Han population. J. Transl. Med. 2021, 19, 366. [Google Scholar] [CrossRef]
- Song, Z.; Li, S.; Li, R. An investigation into the correlation of intestinal flora with obesity and gestational diabetes mellitus. Comput. Math. Methods Med. 2022, 2022, 5677073. [Google Scholar] [CrossRef] [PubMed]
- Talford, L.E.; Crost, E.H.; Kavanaugh, D.; Juge, N. Mucin glycan foraging in the human gut microbiome. Front. Genet. 2005, 6, 81. [Google Scholar] [CrossRef]
- Glover, J.S.; Ticer, T.D.; Engevik, M.A. Characterizing the mucin-degrading capacity of the human gut microbiota. Sci. Rep. 2022, 12, 8456. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-datchary, P.; Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Leite, A.Z.; de Campos Rodrigues, N.; Gonzaga, M.I.; Paiolo, J.C.C.; Arantes de Souza, C.; Stefanutto, N.A.V.; Omori, W.P.; Guariz Pinheiro, D.; Brisotti, J.L.; Junior, E.M.; et al. Detection of increased plasma interleukin-6 levels and prevalence of Prevotella copri and Bacteroides vulgatus in the feces of type 2 diabetes patients. Front. Immunol. 2017, 8, 1107. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.I.; Choi, J.H.; Lee, K.H.; Kim, J.M. Bacteroides fragilis enterotoxin induces sulfiredoxin-1 expression in intestinal epithelial cell lines through a mitogen-activated protein kinases- and Nrf2-dependent pathway, leading to the suppression of apoptosis. Int. J. Mol. Sci. 2020, 21, 5383. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Crusell, M.K.W.; Hansen, T.H.; Nielsen, T.; Allin, K.H.; Rühlemann, M.C.; Damm, P.; Vestergaard, H.; Rørbye, C.; Jørgensen, N.R.; Christiansen, O.B.; et al. Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome 2018, 6, 89. [Google Scholar] [CrossRef]
- Wu, Y.; Bible, P.W.; Long, S.; Ming, W.-K.; Ding, W.; Long, Y.; Wen, X.; Li, X.; Deng, X.; Deng, Y.; et al. Metagenomic analysis reveals gestational diabetes mellitus-related microbial regulators of glucose tolerance. Acta Diabetol. 2020, 57, 569–581. [Google Scholar] [CrossRef]
- Gavin, P.G.; Mullaney, J.A.; Loo, D.; Lê Cao, K.-A.; Gottlieb, P.A.; Hill, M.M.; Zipris, D.; Hamilton-Williams, E.E. Intestinal metaproteomics reveals host-microbiota interactions in subjects at risk for type 1 diabetes. Diabetes Care 2018, 41, 2178–2186. [Google Scholar] [CrossRef]
- d’Hennezel, E.; Abubucker, S.; Murphy, L.O.; Cullen, T.W. Total lipopolysaccharide from the human gut microbiome silences toll-like receptor signaling. mSystems 2017, 2, e00046-17. [Google Scholar] [CrossRef] [PubMed]
- Groh, I.A.M.; Riva, A.; Braun, D.; Sutherland, H.G.; Williams, O.; Bakuradze, T.; Pahlke, G.; Richling, E.; Haupt, L.M.; Griffiths, L.R.; et al. Long-term consumption of anthocyanin-rich fruit juice: Impact on gut microbiota and antioxidant markers in lymphocytes of healthy males. Antioxidants 2020, 10, 27. [Google Scholar] [CrossRef]
- Hernandez-Sanabria, E.; Heiremans, E.; Arroyo, M.C.; Props, R.; Leclercq, L.; Snoeys, J.; Van de Wiele, T. Short-term supplementation of celecoxib-shifted butyrate production on a simulated model of the gut microbial ecosystem and ameliorated in vitro inflammation. NPJ Biofilms Microbiomes 2020, 6, 9. [Google Scholar] [CrossRef]
- Pinto, Y.; Frishman, S.; Turjeman, S.; Eshel, A.; Nuriel-Ohayon, M.; Shrossel, O.; Ziv, O.; Walters, W.; Parsonnet, J.; Ley, C.; et al. Gestational diabetes is driven by microbiota-induced inflammation months before diagnosis. Gut 2023, 72, 918–928. [Google Scholar] [CrossRef]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Tarallo, S.; Ferrero, G.; De Filippis, F.; Francavilla, A.; Pasolli, E.; Panero, V.; Cordero, F.; Segata, N.; Grioni, S.; Pensa, R.G.; et al. Stool microRNA profiles reflect different dietary and gut microbiome patterns in healthy individuals. Gut 2022, 71, 1302–1314. [Google Scholar] [CrossRef]
- Linna-Kuosmanen, S.; Bosch, V.T.; Moreau, P.R.; Bouvy-Liivrand, M.; Niskanen, H.; Kansanen, E.; Kivelä, A.; Hartikainen, J.; Hippeläinen, M.; Kokki, H.; et al. NRF2 is a key regulator of endothelial microRNA expression under proatherogenic stimuli. Cardiovasc. Res. 2021, 117, 1339–1357. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Pagès, J.-M. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Front. Microbiol. 2015, 6, 392. [Google Scholar] [CrossRef]
- Scheithauer, T.P.M.; Herrema, H.; Yu, H.; Bakker, G.J.; Winkelmeijer, M.; Soukhatcheva, G.; Dai, D.; Ma, C.; Havik, S.R.; Balvers, M.; et al. Gut-derived bacterial flagellin induces beta-cell inflammation and dysfunction. Gut Microbes. 2022, 14, 2111951. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsharairi, N.A. Exploring the Diet-Gut Microbiota-Epigenetics Crosstalk Relevant to Neonatal Diabetes. Genes 2023, 14, 1017. https://doi.org/10.3390/genes14051017
Alsharairi NA. Exploring the Diet-Gut Microbiota-Epigenetics Crosstalk Relevant to Neonatal Diabetes. Genes. 2023; 14(5):1017. https://doi.org/10.3390/genes14051017
Chicago/Turabian StyleAlsharairi, Naser A. 2023. "Exploring the Diet-Gut Microbiota-Epigenetics Crosstalk Relevant to Neonatal Diabetes" Genes 14, no. 5: 1017. https://doi.org/10.3390/genes14051017
APA StyleAlsharairi, N. A. (2023). Exploring the Diet-Gut Microbiota-Epigenetics Crosstalk Relevant to Neonatal Diabetes. Genes, 14(5), 1017. https://doi.org/10.3390/genes14051017