Tree Surface Temperature in a Primary Tropical Rain Forest

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Studied Species
2.2. Dominant Mature Tree Species
2.3. Saplings of 28 Tree Species
2.4. Thermal Imaging of the Canopy Dominant Tree Species
2.5. Thermal Imaging and Stomatal Conductance of 28 Tree Species Samplings
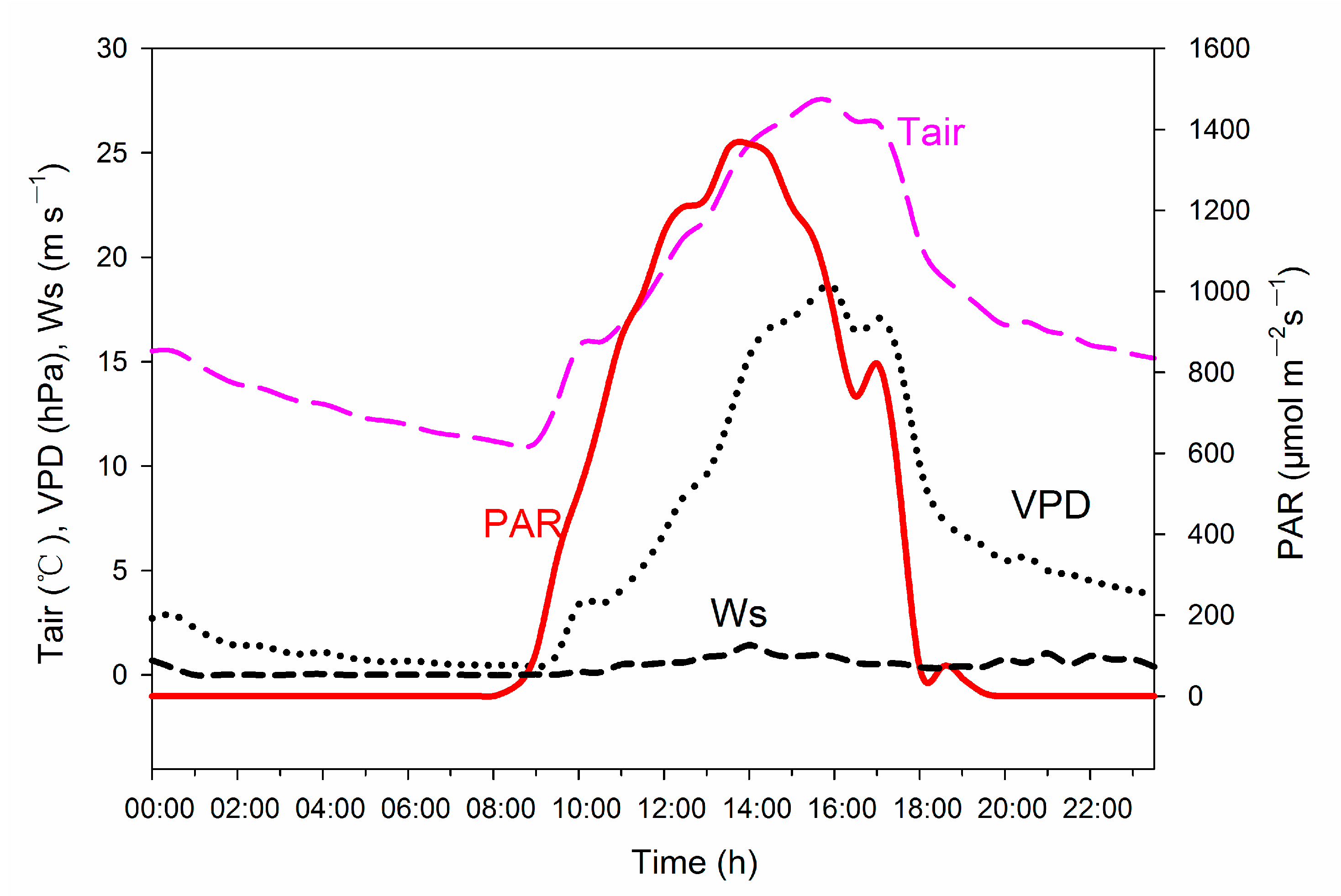
2.6. Environmental Data and Soil Moisture
2.7. Evaluate the Canopy Temperature Change
2.8. Statistical and Analytical Methods
3. Results
3.1. Environmental Conditions
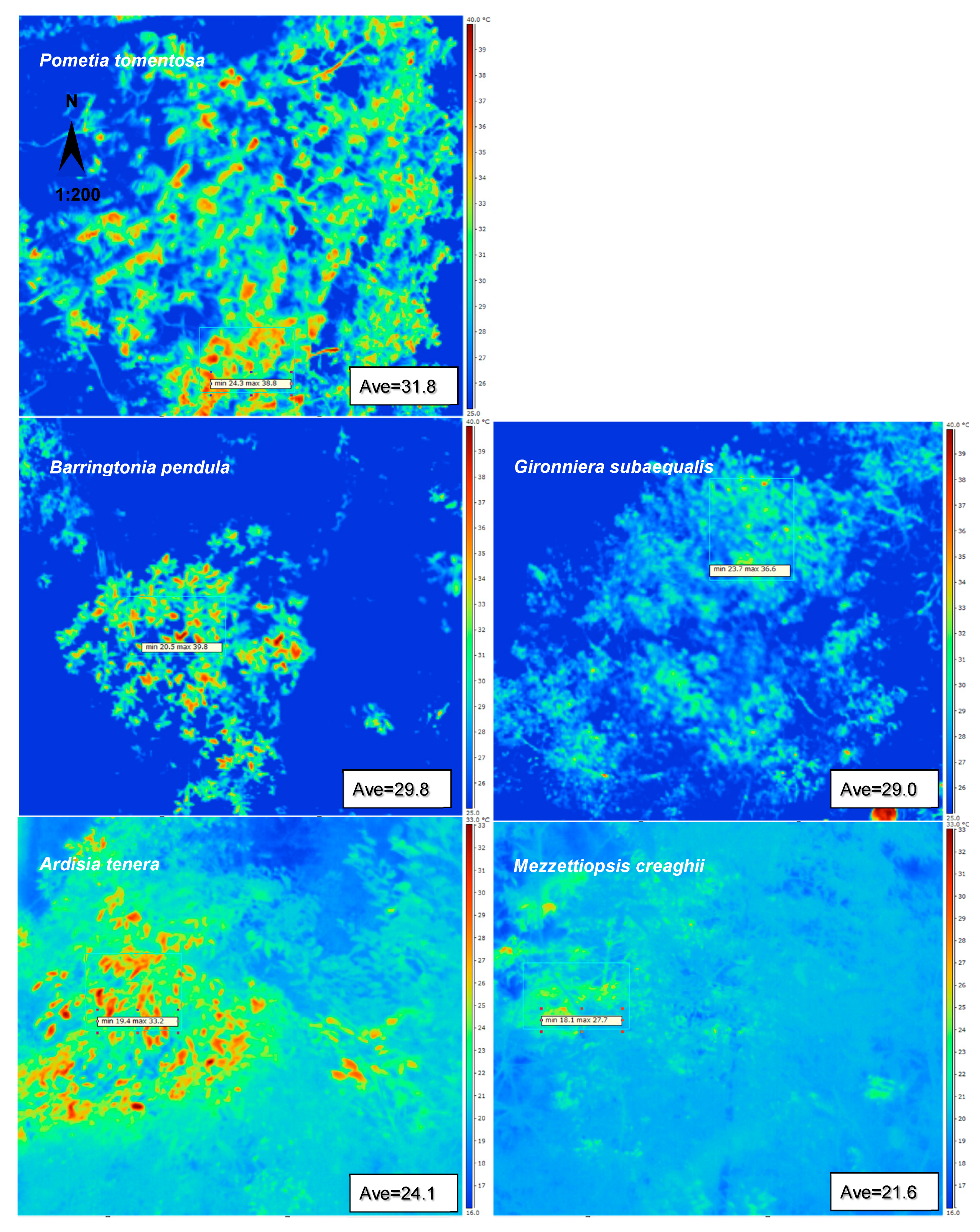
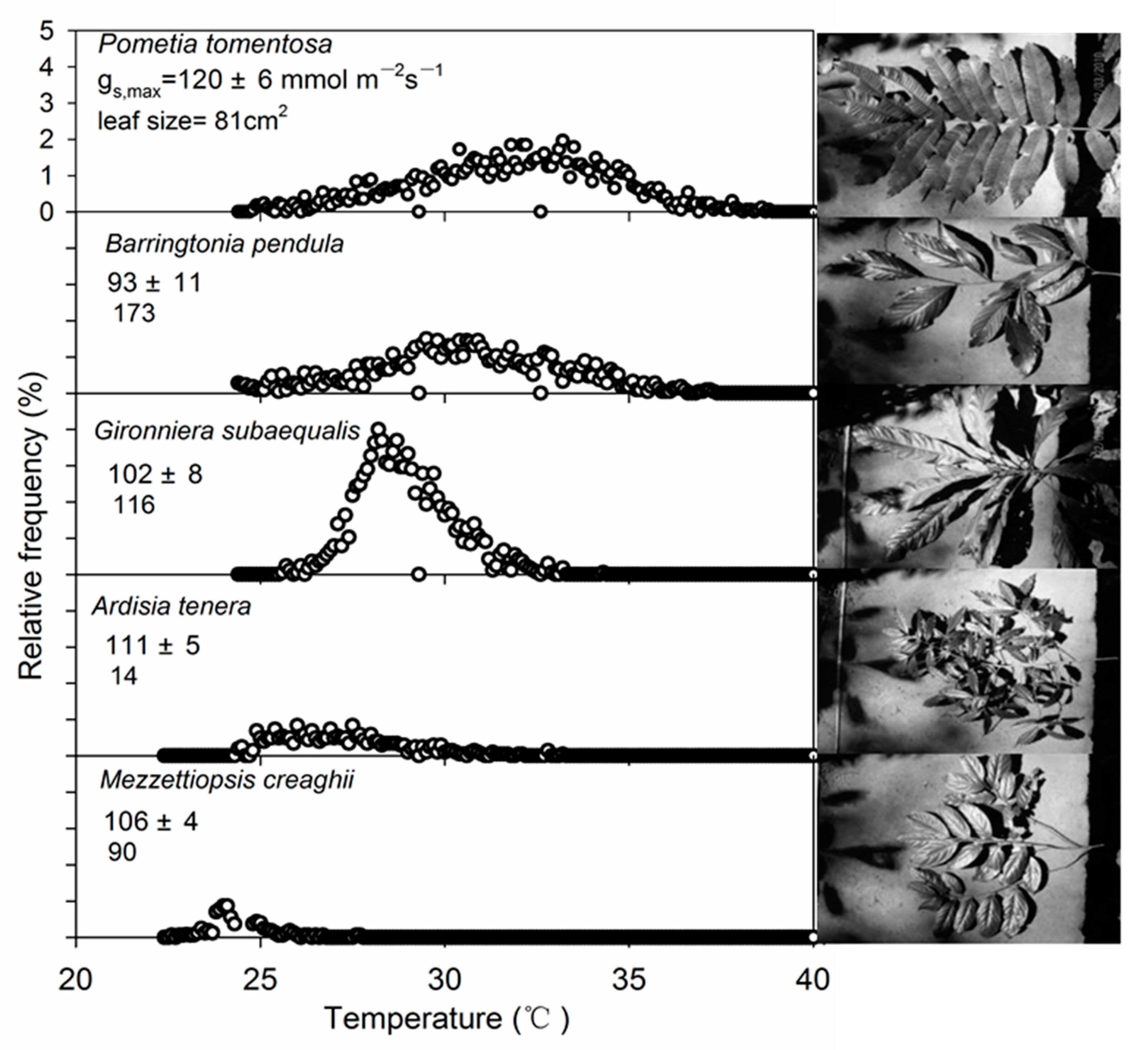
3.2. Spatial and Temporal Temperature Distribution of the Dominant Mature Tree Species
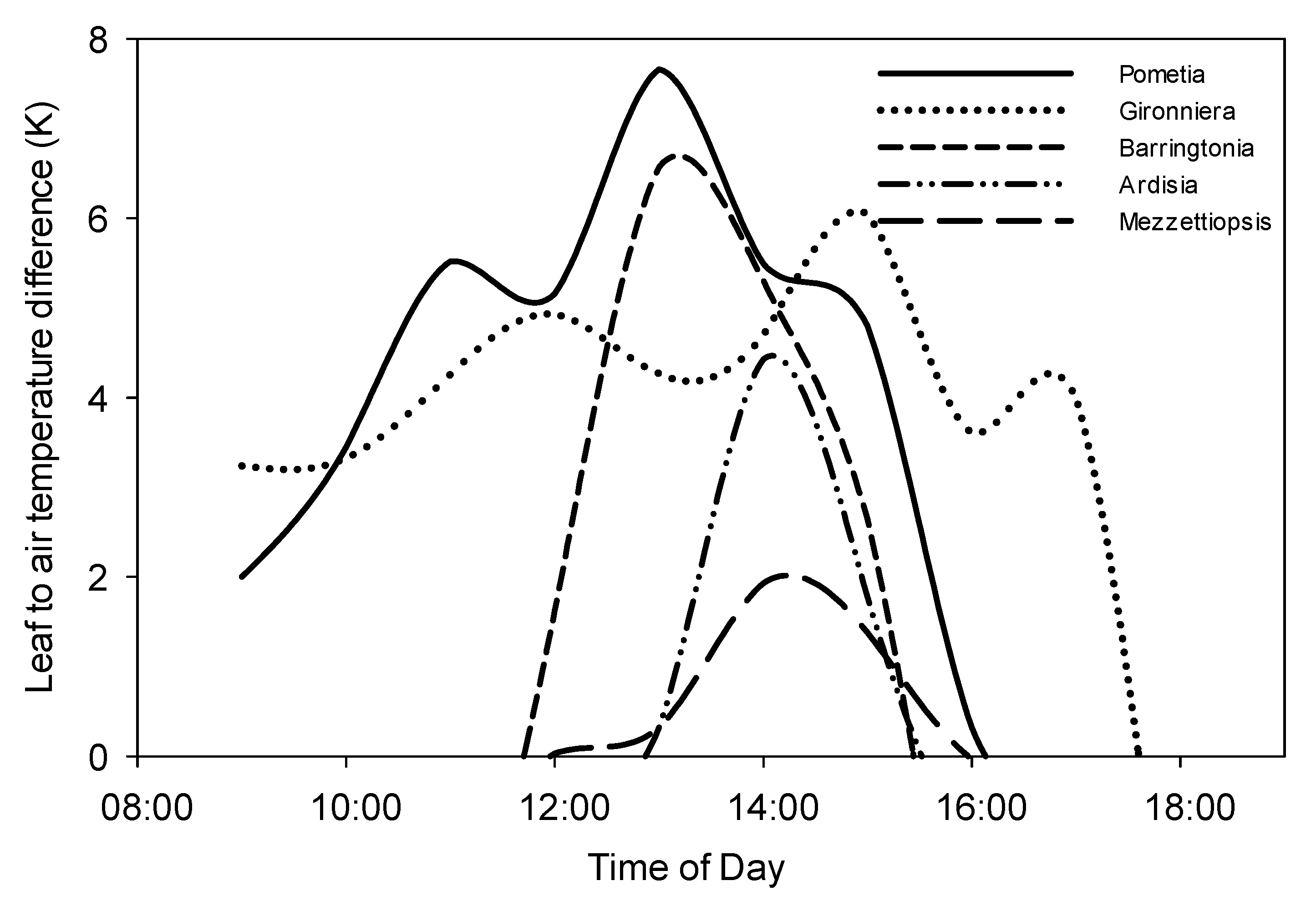
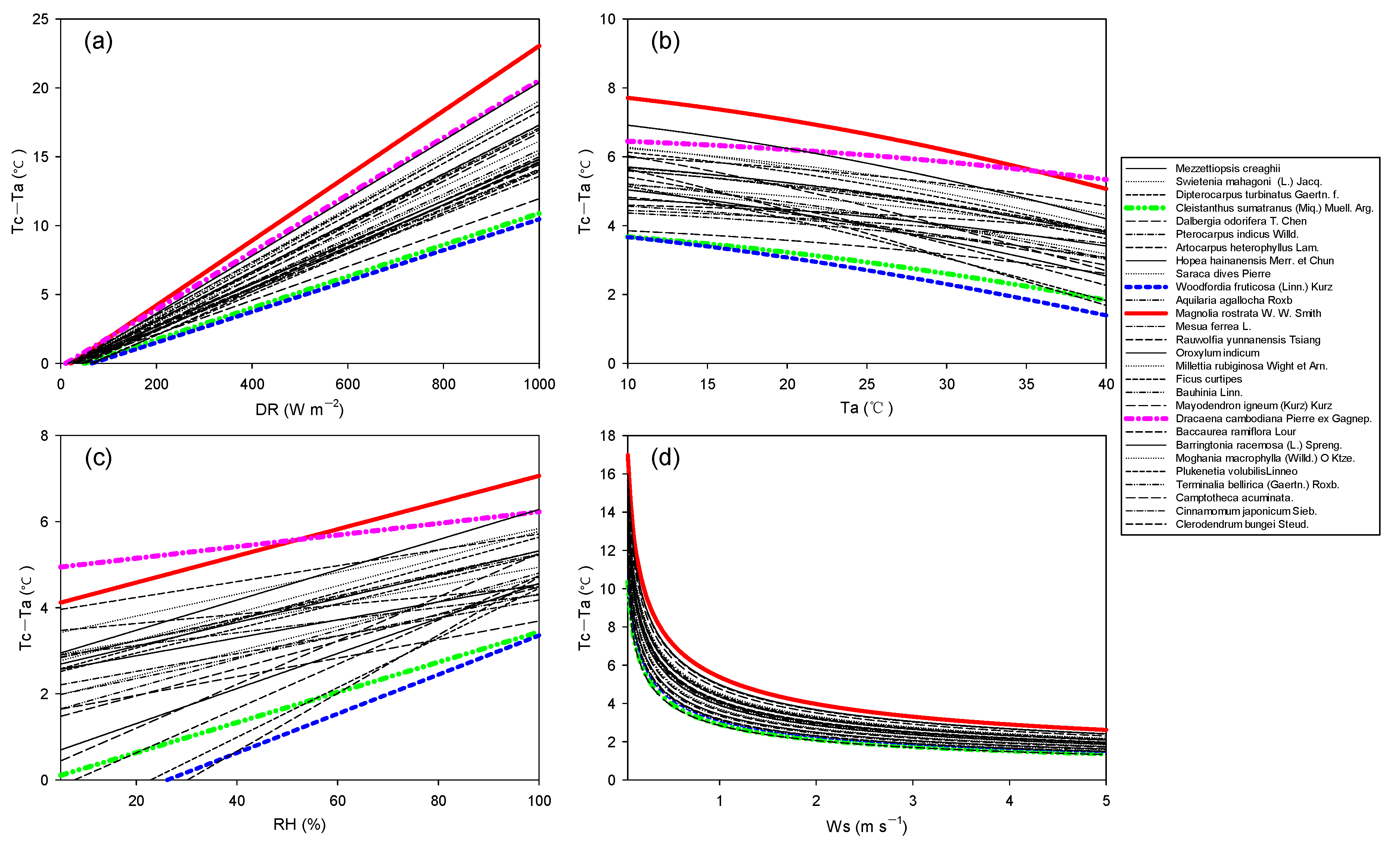
3.3. (Tc–Ta) Variations in a Changing Climate
4. Discussion
5. Conclusions and Implications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Studies in Ecology; Univ of California Press: Berkeley, CA, USA, 1979; Volume 5. [Google Scholar]
- Raich, J.W.; Russell, A.E.; Kitayama, K.; Parton, W.J.; Vitousek, P.M. Temperature Influences Carbon Accumulation in Moist Tropical Forests. Ecology 2006, 87, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.; Zhao, J.; Wang, G.; Chen, M.; Yang, L.; He, C.; Restrepocoupe, N.; Peng, S.; Liu, X.; Ribeiro Da Rocha, H.; et al. Surface conductance for evapotranspiration of tropical forests: Calculations, variations, and controls. Agric. For. Meteorol. 2019, 275, 317–328. [Google Scholar] [CrossRef]
- Lin, H.; Chen, Y.J.; Zhang, H.L.; Fu, P.L.; Fan, Z.X. Stronger cooling effects of transpiration and leaf physical traits of the plants from a hot dry habitat than from a hot wet habitat. Funct. Ecol. 2017, 31, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Govindasamy, V.; Rane, J.; Singh, A.K.; Choudhary, R.L.; Raina, S.K.; George, P.; Aher, L.K.; Singh, N.P. Canopy temperature depression (CTD) and canopy greenness associated with variation in seed yield of soybean genotypes grown in semi-arid environment. S. Afr. J. Bot. 2017, 113, 230–238. [Google Scholar] [CrossRef]
- Rahman, M.A.; Moser, A.; Gold, A.; Rotzer, T.; Pauleit, S. Vertical air temperature gradients under the shade of two contrasting urban tree species during different types of summer days. Sci. Total Environ. 2018, 633, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Page, G.F.; Lienard, J.F.; Pruett, M.J.; Moffett, K.B. Spatiotemporal dynamics of leaf transpiration quantified with time-series thermal imaging. Agric. For. Meteorol. 2018, 256, 304–314. [Google Scholar] [CrossRef]
- Schimel, D.S.; Braswell, B.H.; Holland, E.A.; McKeown, R.; Ojima, D.S.; Painter, T.H.; Parton, W.J.; Townsend, A.R. Climatic, edaphic, and biotic controls over storage and turnover of carbon in soils. Glob. Biogeochem. Cycles 1994, 8, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Solomon, A.M.; Trexler, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef]
- Jobbàgy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Van der Molen, M.K.; Dolman, A.J.; Ciais, P.; Eglin, T.; Gobron, N.; Meir, P.; Peters, W.; Phillips, O.L.; Reichstein, M.; Chen, T.; et al. Drought and ecosystem carbon cycling. Agric. For. Meteorol. 2011, 151, 765–773. [Google Scholar] [CrossRef]
- Daniel, S.; Körner, C. Infrared thermometry of alpine landscapes challenges climatic warming projections. Glob. Chang. Biol. 2010, 16, 2602–2613. [Google Scholar]
- Leuzinger, S.; Körner, C. Tree species diversity affects canopy leaf temperatures in a mature temperate forest. Agric. For. Meteorol. 2007, 146, 29–37. [Google Scholar] [CrossRef]
- Leuzinger, S.; Roland, V.; Körner, C. Tree surface temperature in an urban environment. Agric. For. Meteorol. 2010, 150, 56–62. [Google Scholar] [CrossRef]
- Solomon, S.; Qin, D. Climate Change 2007: The Physical Science Basis, Contribution of Working Group I to the Third Assessment Report of the IPCC.; Cambridge University Press: Cambridge, UK, 2007; p. 996. [Google Scholar]
- Hu, Y.H.; Cao, M.; Lin, L.X. Dynamics of tree species composition and community structure of a tropical seasonal rain forest in Xishuangbanna, Southwest China. Acta Ecol. Sin. 2010, 30, 949–957. [Google Scholar]
- Zhu, H. Forest vegetation of Xishuangbanna, south China. For. Stud. China 2006, 8, 1–58. [Google Scholar]
- Maes, W.H.; Steppe, K. Estimating evapotranspiration and drought stress with ground-based thermal remote sensing in agriculture: A review. J. Exp. Bot. 2012, 63, 4671–4712. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, L.J.; Campbell, C.; Zaragozacastells, J.; Hurry, V.; Atkin, O.K. Impact of growth temperature on scaling relationships linking photosynthetic metabolism to leaf functional traits. Funct. Ecol. 2010, 24, 1181–1191. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy temperature as a crop water stress index. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Keenan, T.F.; Niinemets, U. Global leaf trait estimates biased due to plasticity in the shade. Nat. Plants 2017, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Slot, M.; Reysanchez, C.; Winter, K.; Kitajima, K. Trait-based scaling of temperature-dependent foliar respiration in a species-rich tropical forest canopy. Funct. Ecol. 2014, 28, 1074–1086. [Google Scholar] [CrossRef]
- Zhang, T.; Niinemets, U.; Sheffield, J.; Lichstein, J.W. Shifts in tree functional composition amplify the response of forest biomass to climate. Nature 2018, 556, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Slot, M.; Winter, K. In situ temperature response of photosynthesis of 42 tree and liana species in the canopy of two Panamanian lowland tropical forests with contrasting rainfall regimes. New Phytol. 2017, 214, 1103–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Tc–Ta (K) | T-Range (K) | Tmin (°C) | Tmax (°C) | n |
|---|---|---|---|---|---|
| Pometia tomentosa | 6.4 a | 14.5 ± 1.51 | 24.3 ± 0.37 | 38.8 ± 1.14 | 9 |
| Barringtonia pendula | 6.1 b | 11.7 ± 2.04 | 23.8 ± 0.61 | 35.5 ± 1.43 | 9 |
| Gironniera subaequalis | 4.7 c | 8.7 ± 1.26 | 25.6 ± 0.25 | 34.3 ± 1.01 | 9 |
| Ardisia tenera | 4.4 d | 13.8 ± 1.05 | 19.4 ± 0.18 | 33.2 ± 0.87 | 9 |
| Mezzettiopsis creaghii | 1.9 e | 10.9 ± 1.07 | 16.8 ± 0.26 | 27.7 ± 0.81 | 9 |
| No. | Species Name | Tc–Ta °C | Leaf Area cm2 | gsmax mmol m−2 s−1 | n |
|---|---|---|---|---|---|
| 1 | Mezzettiopsis creaghii | 0.99 ± 0.86 | 32.83 ± 6.47 | 173.29 ± 49.90 | 5 |
| 2 | Swietenia mahagoni (L.) Jacq. | 2.93 ± 2.00 | 44.32 ± 5.10 | 162.93 ± 27.76 | 5 |
| 3 | Dipterocarpus turbinatus Gaertn. f. | 1.80 ± 0.51 | 106.38 ± 7.44 | 222.04 ± 17.58 | 4 |
| 4 | Cleistanthus sumatranus (Miq.) Muell. Arg. | 0.81 ± 0.11 | 15.98 ± 5.93 | 401.26 ± 29.47 | 5 |
| 5 | Dalbergia odorifera T. Chen | 0.57 ± 0.48 | 14.51 ± 3.83 | 221.41 ± 57.08 | 5 |
| 6 | Pterocarpus indicus Willd. | 0.79 ± 0.15 | 70.93 ± 30.95 | 188.18 ± 65.16 | 5 |
| 7 | Artocarpus heterophyllus Lam. | 1.43 ± 1.92 | 97.80 ± 11.60 | 448.82 ± 44.31 | 5 |
| 8 | Hopea hainanensis Merr. et Chun | 0.83 ± 0.28 | 74.03 ± 12.96 | 480 ± 31.26 | 5 |
| 9 | Saraca dives Pierre | 1.27 ± 0.11 | 100.88 ± 30.05 | 166.59 ± 33.11 | 5 |
| 10 | Woodfordia fruticose (Linn.) Kurz | 1.15 ± 1.73 | 20.10 ± 3.70 | 548.88 ± 48.58 | 5 |
| 11 | Aquilaria agallocha Roxb | 0.31 ± 0.08 | 29.64 ± 4.58 | 217.26 ± 21.22 | 5 |
| 12 | Magnolia rostrata W. W. Smith | 0.59 ± 0.51 | 253.10 ± 116.03 | 170.23 ± 49.83 | 5 |
| 13 | Mesua ferrea L. | 0.63 ± 0.11 | 22.28 ± 7.27 | 127.65 ± 26.22 | 5 |
| 14 | RauwoIfia yunnanensis Tsiang | 0.23 ± 0.22 | 23.86 ± 4.01 | 88.26 ± 43.74 | 5 |
| 15 | Oroxylum indicum | 0.30 ± 0.72 | 59.16 ± 23.86 | 355.58 ± 139.85 | 5 |
| 16 | Millettia rubiginosa Wight et Arn. | 1.68 ± 1.06 | 46.97 ± 16.90 | 235.17 ± 102.95 | 5 |
| 17 | Ficus curtipes | 0.26 ± 0.38 | 70.38 ± 17.56 | 209.97 ± 78.20 | 5 |
| 18 | Bauhinia Linn. | 0.16 ± 0.17 | 65.65 ± 9.33 | 183.91 ± 53.83 | 5 |
| 19 | Mayodendron igneum (Kurz) Kurz | 0.32 ± 0.22 | 41.82 ± 8.00 | 274.25 ± 77.56 | 6 |
| 20 | Dracaena cambodiana Pierre ex Gagnep. | 0.93 ± 0.16 | 95.00 ± 21.43 | 80.78 ± 33.25 | 5 |
| 21 | Baccaurea ramiflora Lour | 0.67 ± 0.19 | 76.53 ± 8.38 | 121.56 ± 21.29 | 5 |
| 22 | Barringtonia racemosa (L.) Spreng. | 0.08 ± 0.24 | 178.54 ± 51.97 | 219.04 ± 79.81 | 5 |
| 23 | Moghania macrophylla (Willd.) O Ktze. | 1.57 ± 0.46 | 115.02 ± 25.24 | 212.76 ± 70.90 | 5 |
| 24 | Plukenetia volubilisLinneo | 2.61 ± 1.69 | 94.54 ± 19.97 | 550.80 ± 93.84 | 5 |
| 25 | Terminalia bellirica (Gaertn.) Roxb. | 0.38 ± 0.66 | 57.87 ± 11.24 | 269.73 ± 46.20 | 5 |
| 26 | Camptotheca acuminata. | 0.76 ± 0.23 | 144.90 ± 34.76 | 398.22 ± 93.55 | 5 |
| 27 | Cinnamomum japonicum Sieb. | 0.07 ± 0.42 | 23.21 ± 4.00 | 187.48 ± 28.19 | 5 |
| 28 | Clerodendrum bungei Steud. | 1.20 ± 0.12 | 171.80 ± 47.19 | 638.84 ± 43.46 | 5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Q.; Sun, C.; Deng, Y.; Bai, H.; Zhang, Y.; Yu, H.; Zhang, J.; Sha, L.; Zhou, W.; Liu, Y. Tree Surface Temperature in a Primary Tropical Rain Forest. Atmosphere 2020, 11, 798. https://doi.org/10.3390/atmos11080798
Song Q, Sun C, Deng Y, Bai H, Zhang Y, Yu H, Zhang J, Sha L, Zhou W, Liu Y. Tree Surface Temperature in a Primary Tropical Rain Forest. Atmosphere. 2020; 11(8):798. https://doi.org/10.3390/atmos11080798
Chicago/Turabian StyleSong, Qinghai, Chenna Sun, Yun Deng, He Bai, Yiping Zhang, Hui Yu, Jing Zhang, Liqing Sha, Wenjun Zhou, and Yuntong Liu. 2020. "Tree Surface Temperature in a Primary Tropical Rain Forest" Atmosphere 11, no. 8: 798. https://doi.org/10.3390/atmos11080798
APA StyleSong, Q., Sun, C., Deng, Y., Bai, H., Zhang, Y., Yu, H., Zhang, J., Sha, L., Zhou, W., & Liu, Y. (2020). Tree Surface Temperature in a Primary Tropical Rain Forest. Atmosphere, 11(8), 798. https://doi.org/10.3390/atmos11080798