
The Influence of the Recording Time in Modelling the Swimming Behaviour of the Freshwater Inbenthic Copepod Bryocamptus pygmaeus

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Data Set
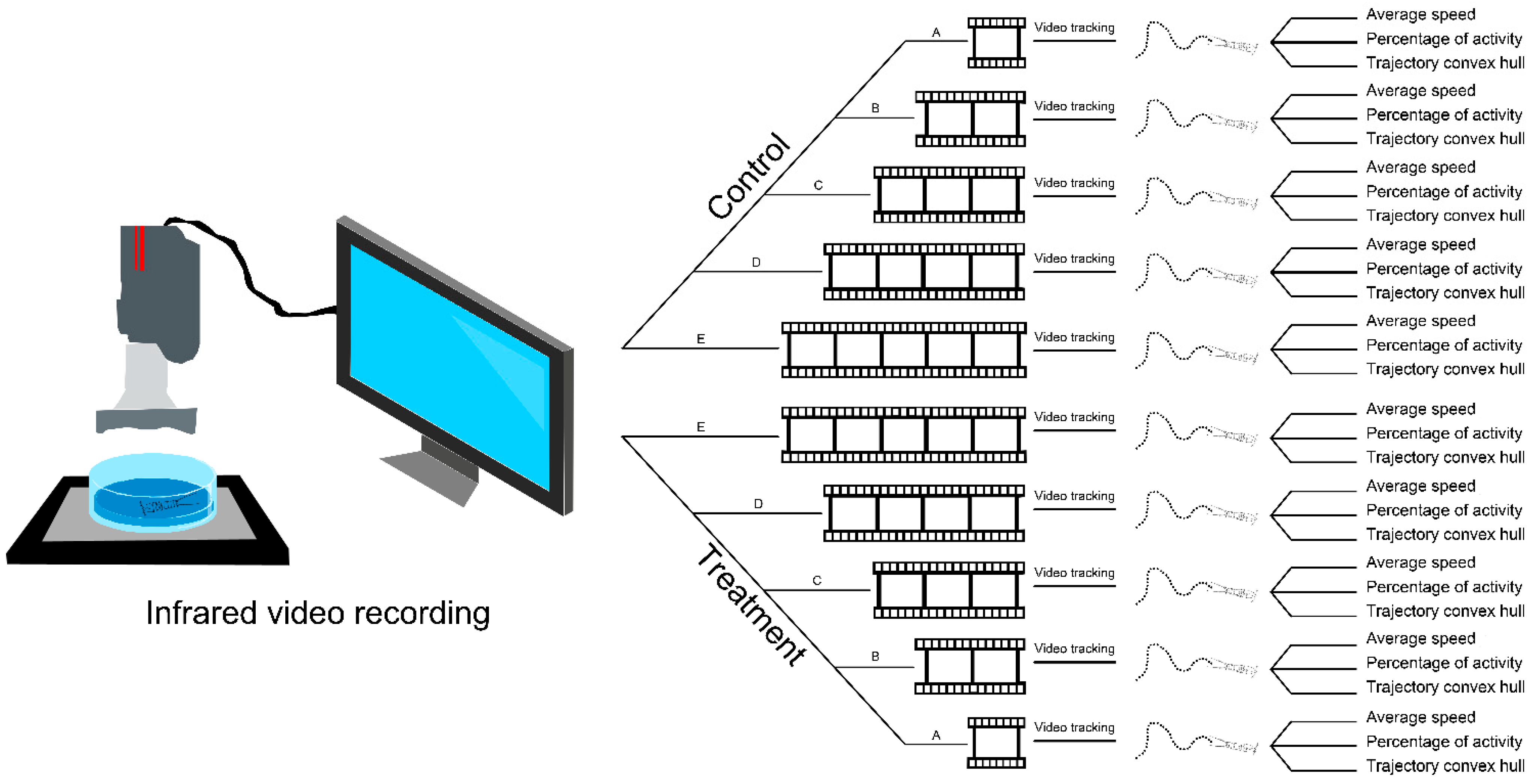
2.2. Behavioural Test and Video Recording
2.3. Video Processing and Trajectory Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Henry, J.; Rodriguez, A.; Wlodkowic, D. Impact of Digital Video Analytics on Accuracy of Chemobehavioural Phenotyping in Aquatic Toxicology. PeerJ 2019, 7, e7367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bownik, A. Daphnia swimming behaviour as a biomarker in toxicity assessment: A review. Sci. Total Environ. 2017, 601–602, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, T.; Di Cicco, M.; Di Censo, D.; Galante, A.; Boscaro, F.; Messana, G.; Galassi, D.M.P. Environmental Risk Assessment of Propranolol in the Groundwater Bodies of Europe. Environ. Pollut. 2019, 255, 113189. [Google Scholar] [CrossRef]
- Di Cicco, M.; Di Lorenzo, T.; Fiasca, B.; Ruggieri, F.; Cimini, A.; Panella, G.; Benedetti, E.; Galassi, D.M.P. Effects of Diclofenac on the Swimming Behavior and Antioxidant Enzyme Activities of the Freshwater Interstitial Crustacean Bryocamptus pygmaeus (Crustacea, Harpacticoida). Sci. Total Environ. 2021, 799, 149461. [Google Scholar] [CrossRef]
- Briffa, S.M.; Sullivan, J.; Siupa, A.; Carnell-Morris, P.; Carboni, M.; Jurkschat, K.; Peters, R.J.B.; Schultz, C.; Seol, K.H.; Kwon, S.-J.; et al. Nanoparticle Tracking Analysis of Gold Nanoparticles in Aqueous Media through an Inter-Laboratory Comparison. J. Vis. Exp. 2020, e61741. [Google Scholar] [CrossRef]
- Shadrin, N.; Litvinchuk, L. Impact of Increased Mineral Particle Concentration on the Behavior, Suspension-Feeding and Reproduction of Acartia clausi (Copepoda). In The Comparative Roles of Suspension-Feeders in Ecosystems; Dame, R.F., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Anufriieva, E.V.; Shadrin, N.V. The swimming behavior of Artemia (Anostraca): New experimental and observational data. Zoology 2014, 117, 415–421. [Google Scholar] [CrossRef]
- Seuront, L. Effect of Salinity on the Swimming Behaviour of the Estuarine Calanoid Copepod Eurytemora affinis. J. Plankton Res. 2006, 28, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Sidler, D.; Michalec, F.-G.; Holzner, M. Behavioral Response of the Freshwater Cyclopoid Copepod Eucyclops serrulatus to Hydropeaking and Thermopeaking in a Laboratory Flume. J. Freshw. Ecol. 2018, 33, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Panadeiro, V.; Rodriguez, A.; Henry, J.; Wlodkowic, D.; Andersson, M. A Review of 28 Free Animal-Tracking Software Applications: Current Features and Limitations. Lab. Anim. 2021, 50, 246–254. [Google Scholar] [CrossRef]
- Budick, S.A.; O’Malley, D.M. Locomotor Repertoire of the Larval Zebrafish: Swimming, Turning and Prey Capture. J. Exp. Biol. 2000, 203, 2565–2579. [Google Scholar] [CrossRef]
- Burgess, H.A.; Granato, M. Modulation of Locomotor Activity in Larval Zebrafish during Light Adaptation. J. Exp. Biol. 2007, 210, 2526–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwaffo, V.; Porfiri, M. Turning Rate Dynamics of Zebrafish Exposed to Ethanol. Int. J. Bifurc. Chaos 2015, 25, 1540006. [Google Scholar] [CrossRef]
- Dunn, T.W.; Mu, Y.; Narayan, S.; Randlett, O.; Naumann, E.A.; Yang, C.-T.; Schier, A.F.; Freeman, J.; Engert, F.; Ahrens, M.B. Brain-Wide Mapping of Neural Activity Controlling Zebrafish Exploratory Locomotion. eLife 2016, 5, e12741. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A Simple Setup to Perform 3D Locomotion Tracking in Zebrafish by Using a Single Camera. Inventions 2018, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, J.-B.; He, K.-J.; Wang, F.; Liu, C.-F. Advances of Zebrafish in Neurodegenerative Disease: From Models to Drug Discovery. Front. Pharmacol. 2021, 12, 713963. [Google Scholar] [CrossRef]
- Barrozo, E.R.; Fowler, D.A.; Beckman, M.L. Exposure to D2-like Dopamine Receptor Agonists Inhibits Swimming in Daphnia magna. Pharmacol. Biochem. Behav. 2015, 137, 101–109. [Google Scholar] [CrossRef]
- Duong, T.-B.; Odhiambo, B.K.; Oldham, D.; Hoffman, M.; Frankel, T.E. Acute Exposure to an Organochlorine Pesticide Alters Locomotor Behavior and Cardiac Function in the Freshwater Invertebrate Daphnia magna. Mar. Freshw. Behav. Physiol. 2021, 54, 51–64. [Google Scholar] [CrossRef]
- Bownik, A.; Jasieczek, M.; Kosztowny, E. Ketoprofen Affects Swimming Behavior and Impairs Physiological Endpoints of Daphnia magna. Sci. Total Environ. 2020, 725, 138312. [Google Scholar] [CrossRef]
- Thorp, J.H.; Rogers, D.C.; Covich, A.P. Freshwater Invertebrates, 4th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Schratzberger, M.; Ingels, J. Meiofauna Matters: The Roles of Meiofauna in Benthic Ecosystems. J. Exp. Mar. Biol. Ecol. 2018, 502, 12–25. [Google Scholar] [CrossRef]
- Williams, R.; Conway, D.V.P.; Hunt, H.G. The Role of Copepods in the Planktonic Ecosystems of Mixed and Stratified Waters of the European Shelf Seas. Hydrobiologia 1994, 292, 521–530. [Google Scholar] [CrossRef]
- Mauchline, J. The Biology of Calanoid Copepods; Advances in Marine Biology; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Turner, J.T. The Importance of Small Planktonic Copepods and Their Roles in Pelagic Marine Food Webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Brun, P.; Payne, M.R.; Kiørboe, T. Trait Biogeography of Marine Copepods—An Analysis across Scales. Ecol. Lett. 2016, 19, 1403–1413. [Google Scholar] [CrossRef] [Green Version]
- Uttieri, M. Trends in Copepod Studies—Distribution, Biology and Ecology; Nova Science Publishers Inc.: New York, NY, USA, 2018; p. 326. [Google Scholar]
- Thompson, R.M.; Dunne, J.A.; Woodward, G. Freshwater Food Webs: Towards a More Fundamental Understanding of Biodiversity and Community Dynamics. Freshw. Biol. 2012, 57, 1329–1341. [Google Scholar] [CrossRef]
- Bellamy, A.R.; Bauer, J.E. Nutritional Support of Inland Aquatic Food Webs by Aged Carbon and Organic Matter. Limnol. Oceanogr. Lett. 2017, 2, 131–149. [Google Scholar] [CrossRef] [Green Version]
- Galassi, D.M.P.; Huys, R.; Reid, J.W. Diversity, Ecology and Evolution of Groundwater Copepods. Freshw. Biol. 2009, 54, 691–708. [Google Scholar] [CrossRef] [Green Version]
- Iannella, M.; Fiasca, B.; Di Lorenzo, T.; Biondi, M.; Di Cicco, M.; Galassi, D.M.P. Jumping into the Grids: Mapping Biodiversity Hotspots in Groundwater Habitat Types across Europe. Ecography 2020, 43, 1825–1841. [Google Scholar] [CrossRef]
- Iannella, M.; Fiasca, B.; Di Lorenzo, T.; Biondi, M.; Di Cicco, M.; Galassi, D.M.P. Spatial Distribution of Stygobitic Crustacean Harpacticoids at the Boundaries of Groundwater Habitat Types in Europe. Sci. Rep. 2020, 10, 19043. [Google Scholar] [CrossRef]
- Dole-Olivier, M.-J.; Galassi, D.M.P.; Marmonier, P.; Creuzé des Châtelliers, M. The Biology and Ecology of Lotic Microcrustaceans. Freshw. Biol. 2000, 44, 63–91. [Google Scholar] [CrossRef]
- Uttieri, M.; Paffenhöfer, G.-A.; Mazzocchi, M.G. Prey Capture in Clausocalanus furcatus (Copepoda: Calanoida). The Role of Swimming Behaviour. Mar. Biol. 2008, 153, 925–935. [Google Scholar] [CrossRef]
- Van Duren, L.A.; Videler, J.J. Swimming Behaviour of Developmental Stages of the Calanoid Copepod Temora longicornis at Different Food Concentrations. Mar. Ecol. Prog. Ser. 1995, 126, 153–161. [Google Scholar] [CrossRef]
- Kiørboe, T. Optimal Swimming Strategies in Mate-Searching Pelagic Copepods. Oecologia 2008, 155, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.; Nishida, S. Swimming Behaviour and Photoresponses of the Iridescent Copepods, Sapphirina gastrica and Sapphirina opalina (Copepoda: Poecilostomatoida). J. Mar. Biol. Assoc. UK 2004, 84, 727–731. [Google Scholar] [CrossRef]
- Uttieri, M.; Nihongi, A.; Mazzocchi, M.G.; Strickler, J.R.; Zambianchi, E. Pre-Copulatory Swimming Behaviour of Leptodiaptomus ashlandi (Copepoda: Calanoida): A Fractal Approach. J. Plankton Res. 2007, 29, 7–26. [Google Scholar] [CrossRef] [Green Version]
- Di Sabatino, A.; Coscieme, L.; Miccoli, F.P.; Cristiano, G. Benthic Invertebrate Assemblages and Leaf-Litter Breakdown along the Eucrenal–Hypocrenal Ecotone of a Rheocrene Spring in Central Italy: Are There Spatial and Seasonal Differences? Ecohydrology 2021, 14, e2289. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Cifoni, M.; Baratti, M.; Pieraccini, G.; Di Marzio, W.D.; Galassi, D.M.P. Four Scenarios of Environmental Risk of Diclofenac in European Groundwater Ecosystems. Environ. Pollut. 2021, 287, 117315. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, T.; Galassi, D.M. Effect of Temperature Rising on the Stygobitic Crustacean Species Diacyclops belgicus: Does Global Warming Affect Groundwater Populations? Water 2017, 9, 951. [Google Scholar] [CrossRef] [Green Version]
- R Developmental Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- McLean, D.J.; Skowron Volponi, M.A. Trajr: An R Package for Characterisation of Animal Trajectories. Ethology 2018, 124, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M. A New Method for Non-Parametric Multivariate Analysis of Variance. Aust. J. Ecol. 2001, 26, 2–46. [Google Scholar]
- Carroll, R.J.; Schneider, H. A Note on Levene’s Tests for Equality of Variances. Stat. Probab. Lett. 1985, 3, 191–194. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef]
- Melvin, S.D.; Wilson, S.P. The Utility of Behavioral Studies for Aquatic Toxicology Testing: A Meta-Analysis. Chemosphere 2013, 93, 2217–2223. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.T.; Ågerstrand, M.; Brooks, B.W.; Allen, J.; Bertram, M.G.; Brodin, T.; Dang, Z.; Duquesne, S.; Sahm, R.; Hoffmann, F.; et al. The Role of Behavioral Ecotoxicology in Environmental Protection. Environ. Sci. Technol. 2021, 55, 5620–5628. [Google Scholar] [CrossRef] [PubMed]
- Seuront, L. When Complexity Rimes with Sanity: Loss of Fractal and Multifractal Behavioural Complexity as an Indicator of Sublethal Contaminations in Zooplankton. In Marine Productivity: Perturbations and Resilience of Socio-Ecosystems; Springer: Cham, Switzerland, 2015; pp. 129–137. [Google Scholar]
- Melvin, S.D.; Petit, M.A.; Duvignacq, M.C.; Sumpter, J.P. Towards Improved Behavioural Testing in Aquatic Toxicology: Acclimation and Observation Times Are Important Factors When Designing Behavioural Tests with Fish. Chemosphere 2017, 180, 430–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amsler, M.O.; Amsler, C.D.; Rittschof, D.; Becerro, M.A.; Mcclintock, J.B. The Use of Computer-Assisted Motion Analysis for Quantitative Studies of the Behaviour of Barnacle (Balanus amphitrite) Larvae. Mar. Freshw. Behav. Physiol. 2006, 39, 259–268. [Google Scholar] [CrossRef]
- Gerhardt, A. Aquatic Behavioral Ecotoxicology—Prospects and Limitations. Hum. Ecol. Risk Assess. Int. J. 2007, 13, 481–491. [Google Scholar] [CrossRef]
- Venkateswara Rao, J.; Kavitha, P.; Jakka, N.M.; Sridhar, V.; Usman, P.K. Toxicity of Organophosphates on Morphology and Locomotor Behavior in Brine Shrimp, Artemia salina. Arch. Environ. Contam. Toxicol. 2007, 53, 227–232. [Google Scholar] [CrossRef]
- Venkateswara Rao, J.; Begum, G.; Pallela, R.; Usman, P.K.; Rao, R.N. Changes in Behavior and Brain Acetylcholinesterase Activity in Mosquito Fish, Gambusia affinis in Response to the Sub-Lethal Exposure to Chlorpyrifos. Int. J. Environ. Res. Public Health 2005, 2, 478–483. [Google Scholar]
- Bradley, C.J.; Strickler, J.R.; Buskey, E.J.; Lenz, P.H. Swimming and Escape Behavior in Two Species of Calanoid Copepods from Nauplius to Adult. J. Plankton Res. 2013, 35, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Dur, G.; Souissi, S.; Schmitt, F.; Michalec, F.-G.; Mahjoub, M.-S.; Hwang, J.-S. Effects of Animal Density, Volume, and the Use of 2D/3D Recording on Behavioral Studies of Copepods. Hydrobiologia 2011, 666, 197–214. [Google Scholar] [CrossRef]
- McAllen, R.; Taylor, A. The Effect of Salinity Change on the Oxygen Consumption and Swimming Activity of the High-Shore Rockpool Copepod Tigriopus brevicornis. J. Exp. Mar. Biol. Ecol. 2001, 263, 227–240. [Google Scholar] [CrossRef]
- Worton, B.J. A Convex Hull-Based Estimator of Home-Range Size. Biometrics 1995, 51, 1206–1215. [Google Scholar] [CrossRef]
- Baiamonte, M.; Parker, M.O.; Vinson, G.P.; Brennan, C.H. Sustained Effects of Developmental Exposure to Ethanol on Zebrafish Anxiety-Like Behaviour. PLoS ONE 2016, 11, e0148425. [Google Scholar] [CrossRef]
- Schnörr, S.J.; Steenbergen, P.J.; Richardson, M.K.; Champagne, D.L. Measuring Thigmotaxis in Larval Zebrafish. Behav. Brain Res. 2012, 228, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Kohler, S.A.; Parker, M.O.; Ford, A.T. Shape and Size of the Arenas Affect Amphipod Behaviours: Implications for Ecotoxicology. PeerJ 2018, 6, e5271. [Google Scholar] [CrossRef]
- Loretto, D.; Vieira, M.V. The Effects of Reproductive and Climatic Seasons on Movements in the Black-Eared Opossum (Didelphis aurita Wied-Neuwied, 1826). J. Mammal. 2005, 86, 287–293. [Google Scholar] [CrossRef]
- Almeida, P.J.; Vieira, M.V.; Kajin, M.; Forero-Medina, G.; Cerqueira, R. Indices of Movement Behaviour: Conceptual Background, Effects of Scale and Location Errors. Zool. Curitiba 2010, 27, 674–680. [Google Scholar] [CrossRef]
- Sumpter, J.P.; Donnachie, R.L.; Johnson, A.C. The Apparently Very Variable Potency of the Anti-Depressant Fluoxetine. Aquat. Environ. 2014, 151, 57–60. [Google Scholar] [CrossRef]
- Faimali, M.; Gambardella, C.; Costa, E.; Piazza, V.; Morgana, S.; Estévez-Calvar, N.; Garaventa, F. Old Model Organisms and New Behavioral End-Points: Swimming Alteration as an Ecotoxicological Response. Blue Growth Mar. Environ. Saf. 2017, 128, 36–45. [Google Scholar]
- Goetze, E.; Kiørboe, T. Heterospecific Mating and Species Recognition in the Planktonic Marine Copepods Temora stylifera and T. longicornis. Mar. Ecol. Prog. Ser. 2008, 370, 185–198. [Google Scholar] [CrossRef]
- Kane, A.S.; Salierno, J.D.; Brewer, S.K. Fish Models in Behavioral Toxicology: Automated Techniques, Updates and Perspectives. Methods Aquat. Toxicol. 2005, 2, 559–590. [Google Scholar]
- Ayari, A.; Jelassi, R.; Chedliya, G.; Nasri-Ammar, K. Effect of Age, Sex and Mutual Interaction on the Locomotor Behavior of Orchestia gammarellus in the Supralittoral Zone of Ghar El Melh Lagoon (Bizerte, Tunisia). Biol. Rhythm Res. 2015, 46, 1–18. [Google Scholar] [CrossRef]
- Cherry, T.-R.; Kohler, S.A.; Ford, A.T. Sex Specific Differences Recorded in the Behavior of an Amphipod: Implications for Behavioral Toxicology. Front. Mar. Sci. 2020, 7, 370. [Google Scholar] [CrossRef]
- Williams, K.L.; Navins, K.C.; Lewis, S.E. Behavioral Responses to Predation Risk in Brooding Female Amphipods (Gammarus pseudolimnaeus). J. Freshw. Ecol. 2016, 31, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.R.; Olechnowicz, S.; Krätschmer, P.; Jepson, J.; Edwards, C.; Edwards, J. Small Animal Video Tracking for Activity and Path Analysis Using a Novel Open-Source Multi-Platform Application (AnimApp). Sci. Rep. 2019, 9, 12343. [Google Scholar] [CrossRef] [PubMed]
- Walsh, R.N.; Cummins, R.A. The Open-Field Test: A Critical Review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef]
- Kohler, S.A.; Parker, M.O.; Ford, A.T. High-Throughput Screening of Psychotropic Compounds: Impacts on Swimming Behaviours in Artemia franciscana. Toxics 2021, 9, 64. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
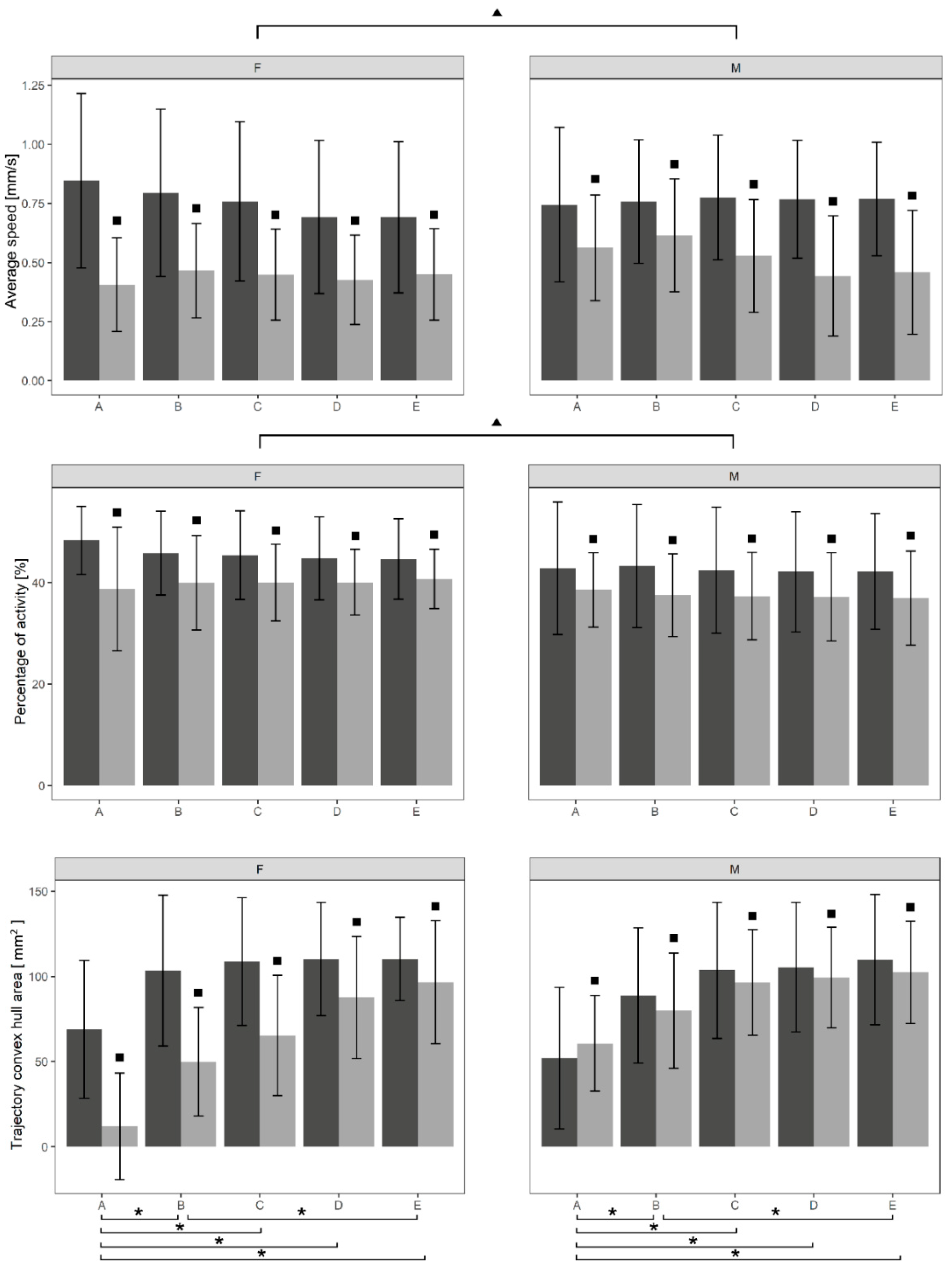
| Time | Condition | Sex | AS (mm s−1) | PA (%) | CH (mm2) |
|---|---|---|---|---|---|
| A | CON | F | 0.85 ± 0.37 | 48.27 ± 6.71 | 68.78 ± 40.49 |
| A | CON | M | 0.74 ± 0.33 | 42.82 ± 13.06 | 52.03 ± 41.57 |
| A | TRE | F | 0.41 ± 0.20 | 38.69 ± 12.17 | 11.91 ± 31.25 |
| A | TRE | M | 0.56 ± 0.22 | 38.53 ± 7.32 | 60.64 ± 28.14 |
| B | CON | F | 0.79 ± 0.35 | 45.75 ± 8.27 | 103.40 ± 44.25 |
| B | CON | M | 0.76 ± 0.26 | 43.25 ± 12.08 | 88.83 ± 39.82 |
| B | TRE | F | 0.47 ± 0.20 | 39.91 ± 9.28 | 49.93 ± 31.84 |
| B | TRE | M | 0.61 ± 0.24 | 37.47 ± 8.17 | 79.88 ± 33.83 |
| C | CON | F | 0.76 ± 0.35 | 45.35 ± 8.74 | 108.66 ± 37.46 |
| C | CON | M | 0.78 ± 0.26 | 42.39 ± 12.41 | 103.57 ± 39.89 |
| C | TRE | F | 0.45 ± 0.19 | 39.94 ± 7.57 | 65.23 ± 35.37 |
| C | TRE | M | 0.53 ± 0.24 | 37.30 ± 8.63 | 96.41 ± 30.90 |
| D | CON | F | 0.69 ± 0.32 | 44.75 ± 8.18 | 110.10 ± 33.30 |
| D | CON | M | 0.77 ± 0.25 | 42.09 ± 11.81 | 105.36 ± 38.13 |
| D | TRE | F | 0.43 ± 0.19 | 40.00 ± 6.47 | 87.57 ± 35.92 |
| D | TRE | M | 0.44 ± 0.25 | 37.16 ± 8.72 | 99.34 ± 29.51 |
| E | CON | F | 0.69 ± 0.32 | 44.61 ± 7.90 | 110.25 ± 24.35 |
| E | CON | M | 0.77 ± 0.24 | 42.15 ± 11.39 | 109.74 ± 38.25 |
| E | TRE | F | 0.45 ± 0.19 | 40.67 ± 5.84 | 96.55 ± 36.09 |
| E | TRE | M | 0.46 ± 0.26 | 36.90 ± 9.28 | 102.46 ± 29.98 |
| Degrees of Freedom | Sums of Squares | Mean Square | Pseudo F | p-Value | ||
|---|---|---|---|---|---|---|
| Average speed (mm s−1) | Time | 4 | 0.0322 | 0.0080606 | 0.591 | 0.655 |
| Residuals | 395 | 5.1142 | 0.0136379 | |||
| Total | 399 | 5.1464 | ||||
| Condition | 1 | 0.0375 | 0.037485 | 3.1192 | 0.091 | |
| Residuals | 398 | 4.5426 | 0.12018 | |||
| Total | 399 | 4.5801 | ||||
| Sex | 1 | 0.0572 | 0.057226 | 4.1408 | 0.041 | |
| Residuals | 398 | 5.2240 | 0.013820 | |||
| Total | 399 | 5.2813 | ||||
| Percentage of activity (%) | Time | 4 | 0.01339 | 0.0033479 | 0.5853 | 0.656 |
| Residuals | 395 | 2.14502 | 0.0057200 | |||
| Total | 399 | 2.15841 | ||||
| Condition | 1 | 0.00033 | 0.0003292 | 0.0564 | 0.821 | |
| Residuals | 398 | 2.20490 | 0.0058331 | |||
| Total | 399 | 2.20523 | ||||
| Sex | 1 | 0.06317 | 0.063167 | 11.448 | 0.002 | |
| Residuals | 398 | 2.08568 | 0.005518 | |||
| Total | 399 | 2.14885 | ||||
| Convex hull area (mm2) | Time | 4 | 84.89 | 21.2217 | 6.1177 | <0.001 |
| Residuals | 395 | 1300.85 | 3.4689 | |||
| Total | 399 | 1385.74 | ||||
| Condition | 1 | 5.74 | 5.7431 | 1.2862 | 0.235 | |
| Residuals | 398 | 1687.82 | 4.4651 | |||
| Total | 399 | 1693.56 | ||||
| Sex | 1 | 17.22 | 17.2221 | 4.2178 | 0.038 | |
| Residuals | 398 | 1543.46 | 4.0832 | |||
| Total | 399 | 1560.68 |
| Degrees of Freedom | Sums of Squares | Mean Square | Pseudo F | p-Value | ||
|---|---|---|---|---|---|---|
| Average speed (mm s−1) | Time | 4 | 0.0077 | 0.00191 | 0.064 | 0.989 |
| Cond | 1 | 2.5777 | 2.57772 | 86.033 | <0.001 | |
| Sex | 1 | 0.1998 | 0.19975 | 6.667 | 0.013 | |
| Time × Cond | 4 | 0.0301 | 0.00753 | 0.251 | 0.889 | |
| Time × Sex | 4 | 0.0011 | 0.00029 | 0.010 | 0.999 | |
| Cond × Sex | 4 | 0.0399 | 0.03993 | 1.333 | 0.258 | |
| Time × Cond × Sex | 4 | 0.0353 | 0.00883 | 0.295 | 0.894 | |
| Residuals | 380 | 10.7863 | 0.02996 | |||
| Total | 399 | 13.678 | ||||
| Percentage of activity (%) | Time | 4 | 0.0031 | 0.000781 | 0.0754 | 0.980 |
| Cond | 1 | 0.2752 | 0.275215 | 26.5828 | <0.001 | |
| Sex | 1 | 0.812 | 0.81153 | 7.8385 | 0.006 | |
| Time × Cond | 4 | 0.122 | 0.003059 | 0.2955 | 0.887 | |
| Time × Sex | 4 | 0.0006 | 0.000154 | 0.0149 | 1.000 | |
| Cond × Sex | 4 | 0.036 | 0.003611 | 0.3488 | 0.554 | |
| Time × Cond × Sex | 4 | 0.180 | 0.004498 | 0.4345 | 0.786 | |
| Residuals | 380 | 3.7271 | 0.010275 | |||
| Total | 399 | 4.1211 | ||||
| Convex hull area (mm2) | Time | 4 | 405.76 | 101.441 | 15.6249 | <0.001 |
| Cond | 1 | 111.92 | 111.920 | 17.2391 | <0.001 | |
| Sex | 1 | 17.76 | 17.760 | 2.7355 | 0.091 | |
| Time × Cond | 4 | 17.08 | 4.269 | 0.6576 | 0.645 | |
| Time × Sex | 4 | 13.61 | 3.403 | 0.5242 | 0.724 | |
| Cond × Sex | 4 | 51.35 | 51.346 | 7.9089 | 0.009 | |
| Time × Cond × Sex | 4 | 9.47 | 2.368 | 0.3674 | 0.836 | |
| Residuals | 380 | 2337.21 | 6.492 | |||
| Total | 399 | 2964.16 |
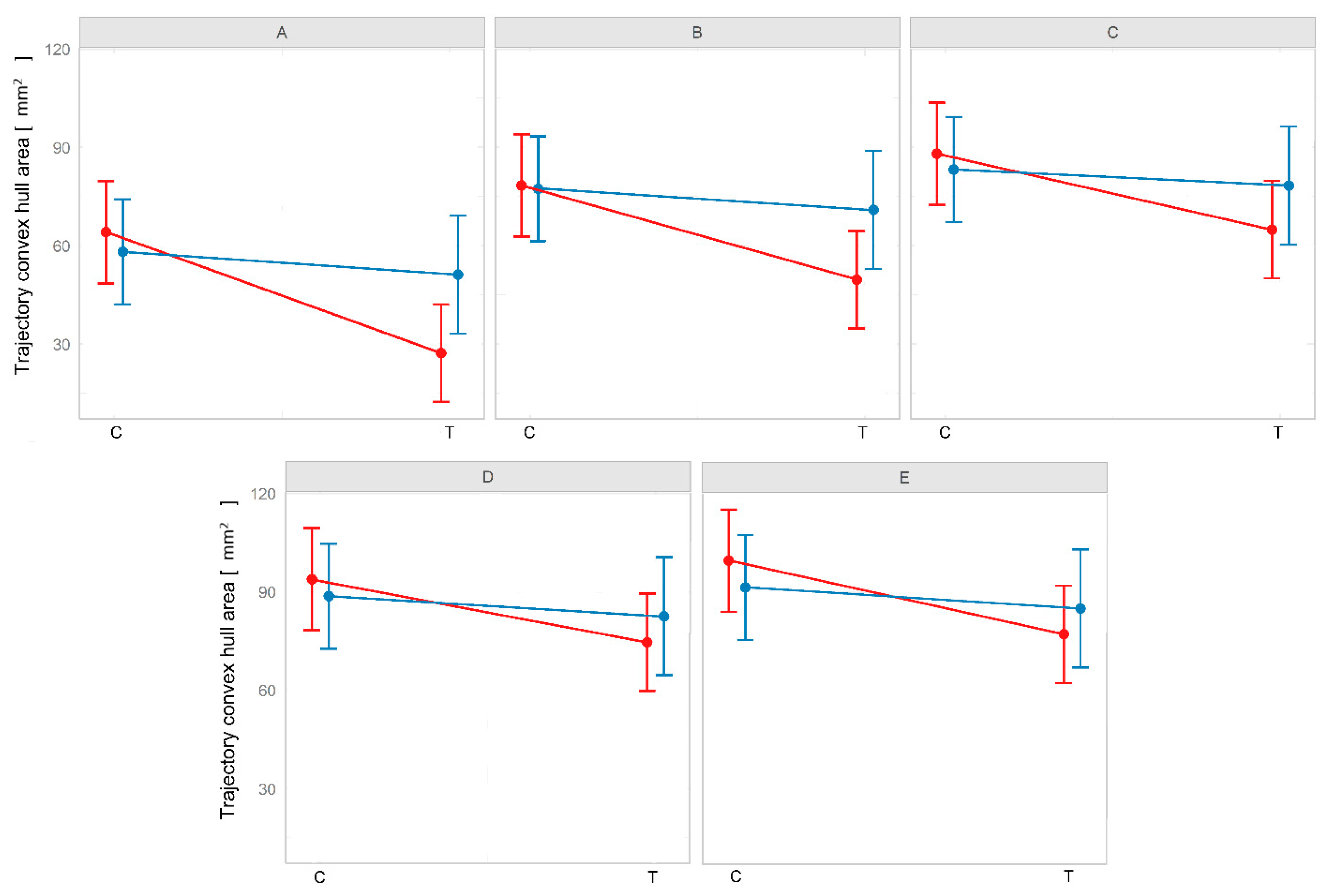
| Comparison | Permutational t | p-Value |
|---|---|---|
| A–B | −2.992 | 0.002773 |
| A–C | −4.453 | 8.48 × 10−6 |
| A–D | −5.278 | 1.308 × 10−9 |
| A–E | −5.818 | 5.953 × 10−9 |
| B–C | −1.635 | 0.1021 |
| B–D | −2.647 | 0.008119 |
| B–E | −3.307 | 0.0009434 |
| C–D | −1.075 | 0.2822 |
| C–E | −1.775 | 0.07582 |
| D–E | −0.6952 | 0.487 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Cicco, M.; Uttieri, M.; Di Lorenzo, T.; Fiasca, B.; Vaccarelli, I.; Tabilio Di Camillo, A.; Galassi, D.M.P. The Influence of the Recording Time in Modelling the Swimming Behaviour of the Freshwater Inbenthic Copepod Bryocamptus pygmaeus. Water 2022, 14, 1996. https://doi.org/10.3390/w14131996
Di Cicco M, Uttieri M, Di Lorenzo T, Fiasca B, Vaccarelli I, Tabilio Di Camillo A, Galassi DMP. The Influence of the Recording Time in Modelling the Swimming Behaviour of the Freshwater Inbenthic Copepod Bryocamptus pygmaeus. Water. 2022; 14(13):1996. https://doi.org/10.3390/w14131996
Chicago/Turabian StyleDi Cicco, Mattia, Marco Uttieri, Tiziana Di Lorenzo, Barbara Fiasca, Ilaria Vaccarelli, Agostina Tabilio Di Camillo, and Diana Maria Paola Galassi. 2022. "The Influence of the Recording Time in Modelling the Swimming Behaviour of the Freshwater Inbenthic Copepod Bryocamptus pygmaeus" Water 14, no. 13: 1996. https://doi.org/10.3390/w14131996
APA StyleDi Cicco, M., Uttieri, M., Di Lorenzo, T., Fiasca, B., Vaccarelli, I., Tabilio Di Camillo, A., & Galassi, D. M. P. (2022). The Influence of the Recording Time in Modelling the Swimming Behaviour of the Freshwater Inbenthic Copepod Bryocamptus pygmaeus. Water, 14(13), 1996. https://doi.org/10.3390/w14131996