Enhanced Cadmium Accumulation and Tolerance in Transgenic Hairy Roots of Solanum nigrum L. Expressing Iron-Regulated Transporter Gene IRT1

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Media
2.2. Development of Wild-Type and Transgenic Hairy Roots of Solanum nigrum L.
2.3. Determination of IRT1 Gene Expression
2.4. Evaluation of Wild-Type and Transgenic Hairy Roots under Cd Stress
2.5. Determination of Biomass Weight and Cd Content
2.6. Observation of ROS Levels and Cell Viability
2.7. Analysis of Antioxidant Enzyme Activities
2.8. Statistical Analysis
3. Results
3.1. Development of Solanum nigrum L. Hairy Roots Expressing Heterologous IRT1
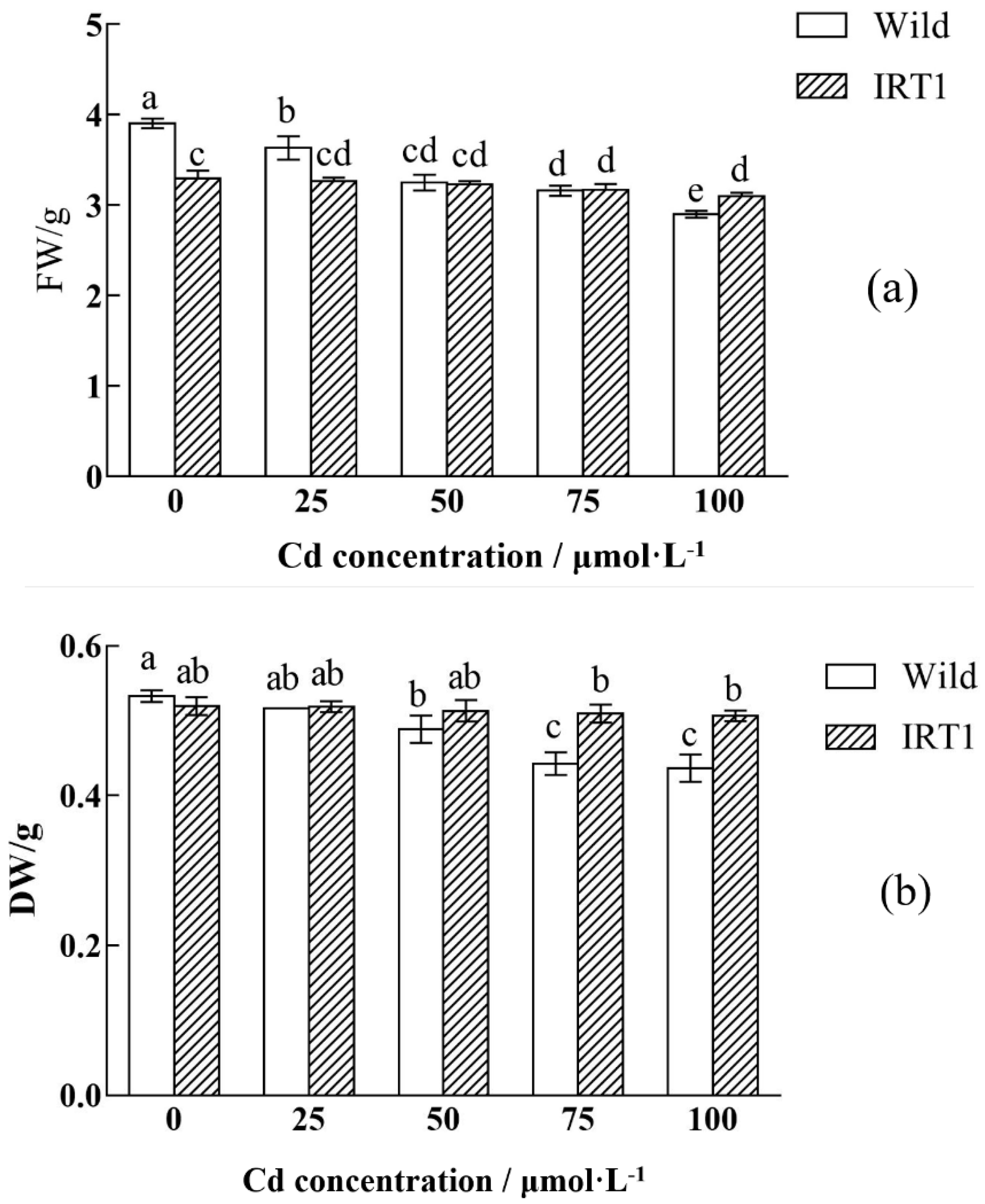
3.2. Growth of the Wild-Type and Transgenic Hairy Roots under Cd Stress
3.3. Cd Accumulation in the Wild-Type and Transgenic Hairy Roots under Cd Stress
3.4. ROS Release and Cell Damage in the Wild-Type and Transgenic Hairy Roots under Cd Stress
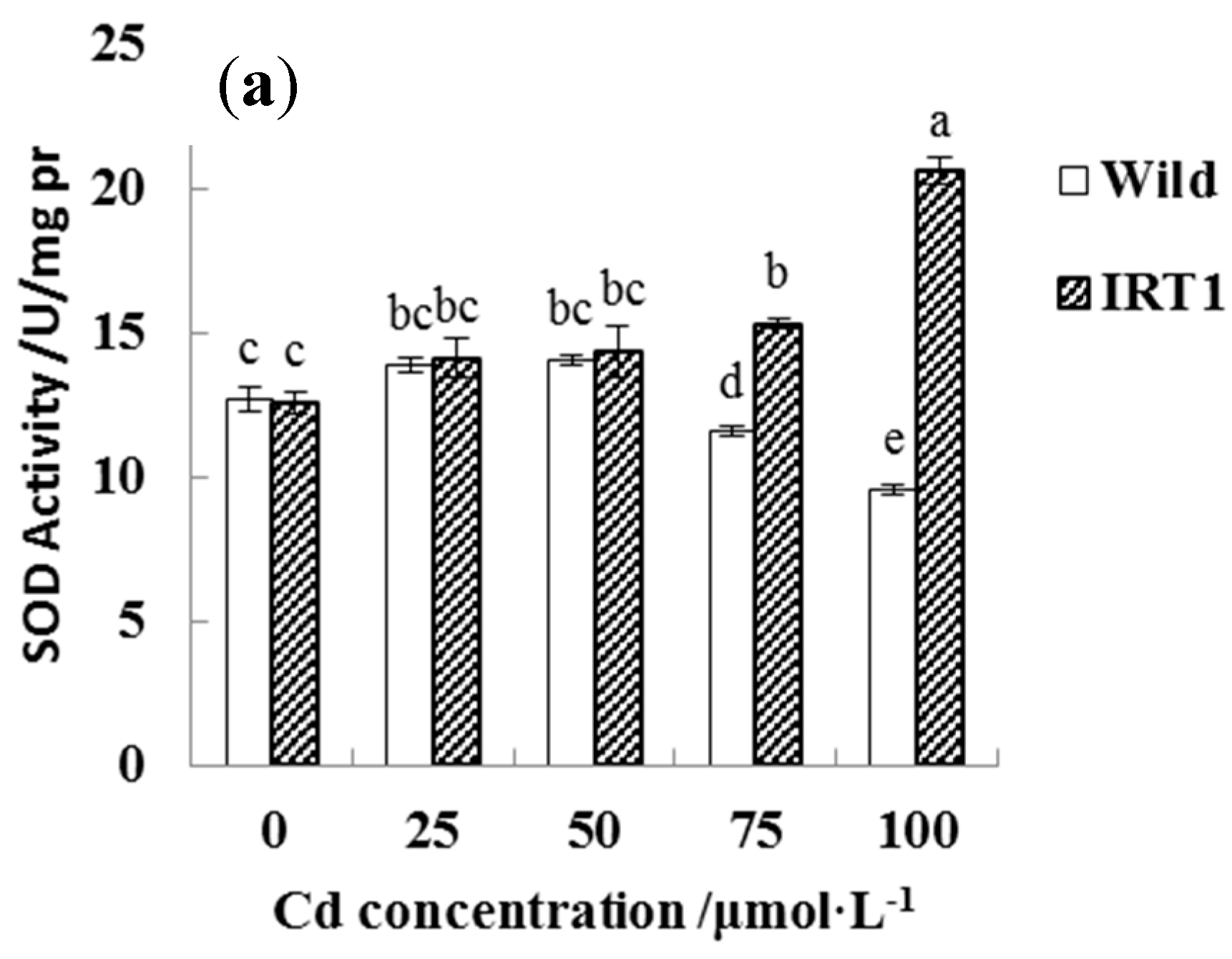
3.5. Antioxidant Enzyme Activities in the the Wild-Type and Transgenic Hairy Roots under Cd Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, C.; Ji, X.H.; Luo, X.G. Phytoremediation of heavy metal pollution: A bibliometric and scientometric analysis from 1989 to 2018. Int. J. Environ. Res. Public Health 2019, 16, 4755. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wang, X.C.; Yang, L.; Chu, Y.Y. Research on progress in combined remediation technologies of heavy metal polluted sediment. Int. J. Environ. Res. Public Health 2019, 16, 5098. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Zhao, S.S.; Rogers, K.M.; Xia, Y.A.; Zhang, B.; Suo, R.; Zhao, Y. A case of milk traceability in small-scale districts-Inner Mongolia of China by nutritional and geographical parameters. Food Chem. 2020, 316, 126332. [Google Scholar] [CrossRef]
- Caruso, M.C.; Braghieri, A.; Capece, A.; Napolitano, F.; Romano, P.; Galgano, F.; Altieri, G.; Genovese, F. Recent updates on the use of agro-food waste for biogas production. Appl. Sci. 2019, 9, 1217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Liu, Y.; Yang, H.Y.; Yan, Q.J.; Yang, S.Q.; Jiang, Z.Q.; Li, S.Z. Biochemical properties and application of a novel beta-1,3-1,4-glucanase from Paenibacillus barengoltzii. Food Chem. 2017, 234, 68–75. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, H.; Jiang, H.; Wang, H.; Chen, K.; Duan, J.; Feng, S.; Wu, G. Regulation of cadmium tolerance and accumulation by miR156 in Arabidopsis. Chemospfere 2020, 242, 125168. [Google Scholar] [CrossRef]
- Zhang, B.; Sun, Q.; Liu, H.J.; Li, S.Z.; Jiang, Z.Q. Characterization of actinidin from Chinese kiwifruit cultivars and its applications in meat tenderization and production of angiotensin I-converting enzyme (ACE) inhibitory peptides. LWT Food Sci. Technol. 2017, 78, 1–7. [Google Scholar] [CrossRef]
- Rehman, M.Z.; Rizwan, M.; Ali, S. Remediation of heavy metal contaminated soils by using Solanum nigrum: A review. Ecotoxicol. Environ. Saf. 2017, 143, 236–248. [Google Scholar] [CrossRef]
- Schueck, M.; Greger, M. Screening the capacity of 34 wetland plant species to remove heavy metals from water. Int. J. Environ. Res. Public Health 2020, 17, 4623. [Google Scholar] [CrossRef]
- Sun, Y.P.; Lu, Q.Y.; Cao, Q.S.; Wang, M.H.; Cheng, X.Y.; Yan, Q. Comparative transcriptome analysis of the molecular mechanism of the hairy roots of Brassica campestris L. in response to cadmium stress. Int. J. Mol. Sci. 2020, 21, 180. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.H.; Lee, C.-M.; Nam, M.J. Cytoprotective effects of taxifolin against cadmium-induced apoptosis in human keratinocytes. Hum. Exp. Toxicol. 2019, 38, 992–1003. [Google Scholar] [CrossRef]
- Jablonska, E.; Socha, K.; Reszka, E.; Wieczorek, E.; Skokowski, J.; Kalinowski, L.; Fendler, W.; Seroczynska, B.; Wozniak, M.; Borawska, M.H.; et al. Cadmium, arsenic, selenium and iron- Implications for tumor progression in breast cancer. Environ. Toxicol. Pharm. 2017, 53, 151–157. [Google Scholar] [CrossRef]
- Xia, L.; Chen, S.; Dahms, H.-U.; Ying, X.; Peng, X.J.E. Cadmium induced oxidative damage and apoptosis in the hepatopancreas of Meretrix meretrix. Ecotoxicology 2016, 25, 959–969. [Google Scholar] [CrossRef]
- Connolly, E.L.; Fett, J.P.; Guerinot, M.L. Expression of the IRT1 metal transporter is controlled by metals at the levels of transcript and protein accumulation. Plant Cell 2002, 14, 1347–1357. [Google Scholar] [CrossRef] [Green Version]
- Das, N.; Bhattacharya, S.; Maiti, M.K. Enhanced cadmium accumulation and tolerance in transgenic tobacco overexpressing rice metal tolerance protein gene OsMTP1 is promising for phytoremediation. Plant Physiol. Biochem. 2016, 105, 297–309. [Google Scholar] [CrossRef]
- Yao, J.; Sun, J.G.; Chen, Y.K.; Shi, L.Q.; Wang, Y. The molecular mechanism underlying cadmium resistance in NHX1 transgenicLemna turoniferawas studied by comparative transcriptome analysis. Plant Cell Tissue Organ Cult. 2020, 143, 189–200. [Google Scholar] [CrossRef]
- Wei, S.H.; Zeng, X.F.; Wang, S.S.; Zhu, J.G.; Ji, D.D.; Li, Y.M.; Jiao, H.J. Hyperaccumulative property of Solanum nigrum L. to Cd explored from cell membrane permeability, subcellular distribution, and chemical form. J. Soils Sediments 2014, 14, 558–566. [Google Scholar] [CrossRef]
- Eide, D.; Broderius, M.; Fett, J.; Guerinot, M.L. A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 5624–5628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.H.; Zhou, Q.X.; Wang, X.; Zhang, K.S.; Guo, G.L.; Qi, L.N.; Ma, Y. A newly-discovered Cd-hyperaccumulator Solatium nigrum L. Chin. Sci. Bull. 2005, 50, 33–38. [Google Scholar] [CrossRef]
- Sun, R.L.; Zhou, Q.X.; Sun, F.H.; Jin, C.X. Antioxidative defense and proline/phytochelatin accumulation in a newly discovered Cd-hyperaccumulator, Solanum nigrum L. Environ. Exp. Bot. 2007, 60, 468–476. [Google Scholar] [CrossRef]
- Ibanez, S.; Talano, M.; Ontanon, O.; Suman, J.; Medina, M.I.; Macek, T.; Agostini, E. Transgenic plants and hairy roots: Exploiting the potential of plant species to remediate contaminants. New Biotechnol. 2016, 33, 625–635. [Google Scholar] [CrossRef]
- Shi, H.P.; Zhu, Y.F.; Wang, Y.L.; Tsang, P.K. Effect of cadmium on cytogenetic toxicity in hairy roots of Wedelia trilobata L. and their alleviation by exogenous CaCl2. Environ. Sci. Pollut. Res. Int. 2014, 21, 1436–1443. [Google Scholar] [CrossRef]
- Rengasamy, B. Cadmium tolerance and antioxidative defenses in hairy roots of the cadmium hyperaccumulator, Thlaspi caerulescens. Biotechnol. Bioeng. 2003, 83, 158–167. [Google Scholar] [CrossRef]
- Kim, Y.K.; Kim, J.K.; Kim, Y.B.; Lee, S.; Kim, S.U.; Park, S.U. Enhanced accumulation of phytosterol and triterpene in hairy root cultures of platycodon grandiflorum by overexpression of Panax ginseng 3-hydroxy-3-methylglutaryl-coenzyme a reductase. J. Agric. Food Chem. 2013, 61, 1928–1934. [Google Scholar] [CrossRef]
- Wang, X.T.; Zhi, J.K.; Liu, X.R.; Zhang, H.; Liu, H.B.; Xu, J.C. Transgenic tobacco plants expressing a P1B-ATPase gene from Populus tomentosa Carr. (PtoHMA5) demonstrate improved cadmium transport. Int. J. Biol. Macromol. 2018, 113, 655–661. [Google Scholar] [CrossRef]
- Radin, I.; Mansilla, N.; Rödel, G.; Steinebrunner, I. The Arabidopsis COX11 homolog is essential for cytochrome oxidase activity. Front. Plant Sci. 2015, 6, 1091. [Google Scholar] [CrossRef] [Green Version]
- Ke, S.; Cheng, X.Y.; Zhang, N.; Hu, H.G.; Yan, Q.; Hou, L.L.; Sun, X.; Chen, Z.N. Cadmium contamination of rice from various polluted areas of China and its potential risks to human health. Environ. Monit. Assess. 2015, 187, 408. [Google Scholar] [CrossRef]
- Kristiansen, K.A.; Jensen, P.E.; Møller, I.M.; Schulz, A. Monitoring reactive oxygen species formation and localisation in living cells by use of the fluorescent probe CM-H2DCFDA and confocal laser microscopy. Physiol. Plant. 2009, 136, 369–383. [Google Scholar] [CrossRef]
- Ma, J.F.; Hiradate, S.; Nomoto, K.; Iwashita, T.; Matsumoto, H. Internal detoxification mechanism of Al in Hydrangea (Identification of Al Form in the Leaves). Plant Physiol. 1997, 113, 1033–1039. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.Y.; Wang, C.; Wang, Y.C.; Guo, Y.C.; Zhao, Y.L.; Yang, C.P.; Gao, C.Q. Overexpression of ThVHAc1 and its potential upstream regulator, ThWRKY7, improved plant tolerance of Cadmium stress. Sci. Rep. 2016, 6, 18752. [Google Scholar] [CrossRef] [Green Version]
- Sivaguru, M.; Liu, J.P.; Kochian, L.V. Targeted expression of SbMATE in the root distal transition zone is responsible for sorghum aluminum resistance. Plant J. 2013, 76, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Borges, K.L.R.; Salvato, F.; Loziuk, P.L.; Muddiman, D.C.; Azevedo, R.A. Quantitative proteomic analysis of tomato genotypes with differential cadmium tolerance. Environ. Sci. Pollut. Res. Int. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dixit, P.; Mukherjee, P.K.; Ramachandran, V.; Eapen, S. Glutathione transferase from Trichoderma virens enhances cadmium tolerance without enhancing its accumulation in transgenic Nicotiana tabacum. PLoS ONE 2011, 6, e16360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Guo, J.J.; He, C.T.; Shen, C.; Huang, Y.Y.; Chen, J.X.; Guo, J.H.; Yuan, J.G.; Yang, Z.Y. Comparative transcriptome analysis between low- and high-cadmium-accumulating genotypes of pakchoi (Brassica chinensis L.) in response to cadmium stress. Environ. Sci. Technol. 2016, 50, 6485–6494. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Sun, J.; Du, L.; Liu, X. Comparative transcriptome analysis of cadmium responses in Solanum nigrum and Solanum torvum. New Phytol. 2012, 196, 110–124. [Google Scholar] [CrossRef]
- Tan, S.; Han, R.; Li, P.; Yang, G.; Li, S.; Zhang, P.; Wang, W.B.; Zhao, W.Z.; Yin, L.P. Over-expression of the MxIRT1 gene increases iron and zinc content in rice seeds. Transgenic Res. 2015, 24, 109–122. [Google Scholar] [CrossRef]
- Barberon, M.; Zelazny, E.; Robert, S.; Conéjéro, G.; Curie, C.; Friml, J.; Vert, G. Monoubiquitin-dependent endocytosis of the IRON-REGULATED TRANSPORTER 1 (IRT1) transporter controls iron uptake in plants. Proc. Natl. Acad. Sci. USA 2011, 108, E450–E458. [Google Scholar] [CrossRef] [Green Version]
- Peng, F.; Wang, C.; Zhu, J.S.; Zeng, J.; Kang, H.Y.; Fan, X.; Sha, L.N.; Zhang, H.Q.; Zhou, Y.H.; Wang, Y. Expression of TpNRAMP5, a metal transporter from Polish wheat (Triticum polonicum L.), enhances the accumulation of Cd, Co and Mn in transgenic Arabidopsis plants. Planta 2018, 247, 1395–1406. [Google Scholar] [CrossRef]
- Fan, W.; Guo, Q.; Liu, C.Y.; Liu, X.Q.; Zhang, M.; Long, D.P.; Xiang, Z.H.; Zhao, A.C. Two mulberry phytochelatin synthase genes confer zinc/cadmium tolerance and accumulation in transgenic Arabidopsis and tobacco. Gene 2018, 645, 95–104. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CdCl2 Concentrations (μM) | Cd Content in Transgenic Hairy Roots (μg/g) | Cd Content in Wild-Type Hairy Roots (μg/g) |
|---|---|---|
| 0 | 0 | 0 |
| 50 | 365.4 ± 2.6 c | 367.7 ± 21.3 c |
| 100 | 886.8 ± 1.0 a | 745.0 ± 10.5 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, P.; Wang, M.; Zhang, T.; Liu, X.; Jiang, H.; Sun, Y.; Cheng, X.; Yan, Q. Enhanced Cadmium Accumulation and Tolerance in Transgenic Hairy Roots of Solanum nigrum L. Expressing Iron-Regulated Transporter Gene IRT1. Life 2020, 10, 324. https://doi.org/10.3390/life10120324
Ye P, Wang M, Zhang T, Liu X, Jiang H, Sun Y, Cheng X, Yan Q. Enhanced Cadmium Accumulation and Tolerance in Transgenic Hairy Roots of Solanum nigrum L. Expressing Iron-Regulated Transporter Gene IRT1. Life. 2020; 10(12):324. https://doi.org/10.3390/life10120324
Chicago/Turabian StyleYe, Peng, Menghua Wang, Teng Zhang, Xiaoyu Liu, He Jiang, Yaping Sun, Xiyu Cheng, and Qiong Yan. 2020. "Enhanced Cadmium Accumulation and Tolerance in Transgenic Hairy Roots of Solanum nigrum L. Expressing Iron-Regulated Transporter Gene IRT1" Life 10, no. 12: 324. https://doi.org/10.3390/life10120324
APA StyleYe, P., Wang, M., Zhang, T., Liu, X., Jiang, H., Sun, Y., Cheng, X., & Yan, Q. (2020). Enhanced Cadmium Accumulation and Tolerance in Transgenic Hairy Roots of Solanum nigrum L. Expressing Iron-Regulated Transporter Gene IRT1. Life, 10(12), 324. https://doi.org/10.3390/life10120324