Effect of Chlorination on Microbiological Quality of Effluent of a Full-Scale Wastewater Treatment Plant



 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Determination of Wastewater Physicochemical Characteristics
2.2. DNA Extraction, Illumina Sequencing, Diversity Indices Estimation and Statistical Analyses
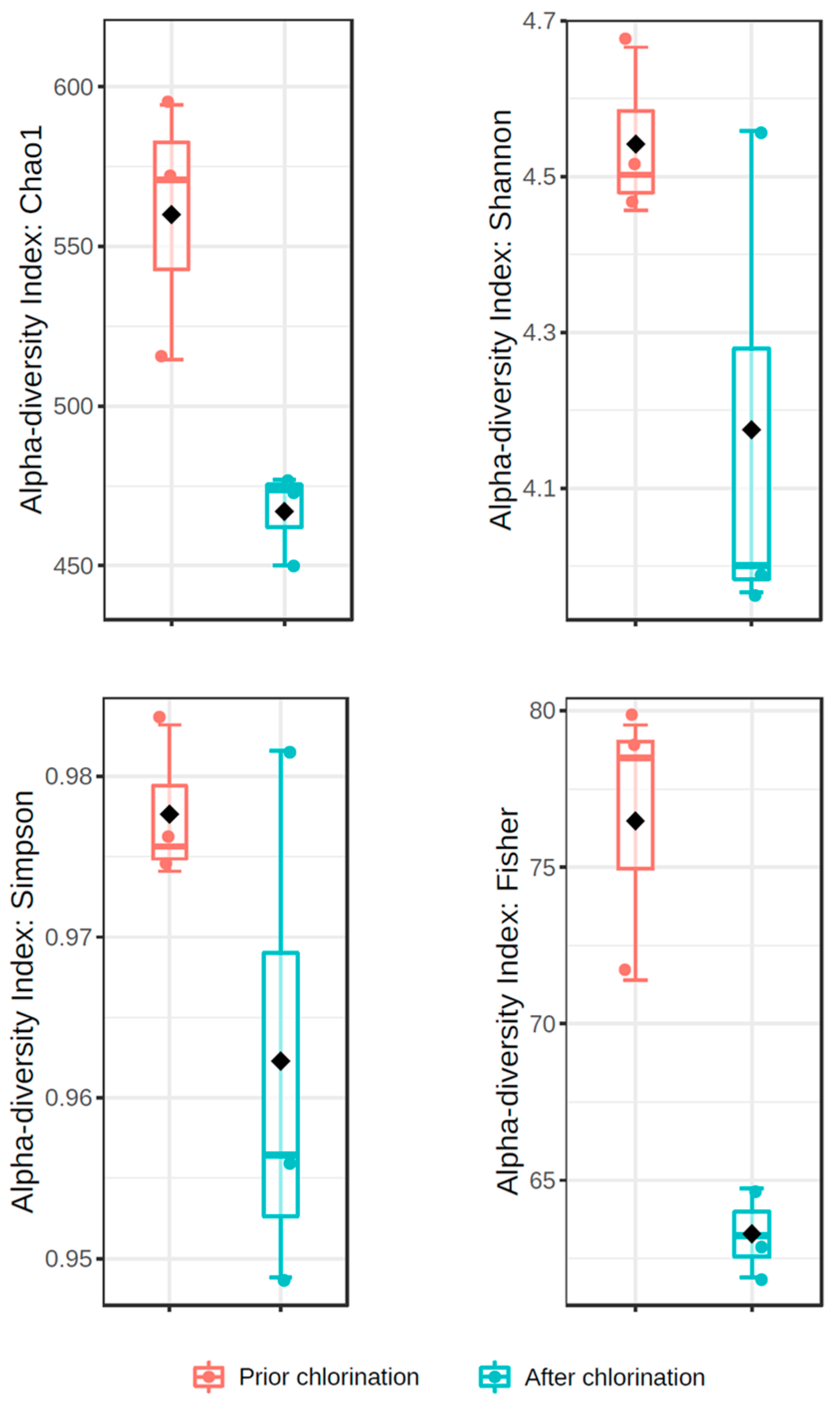
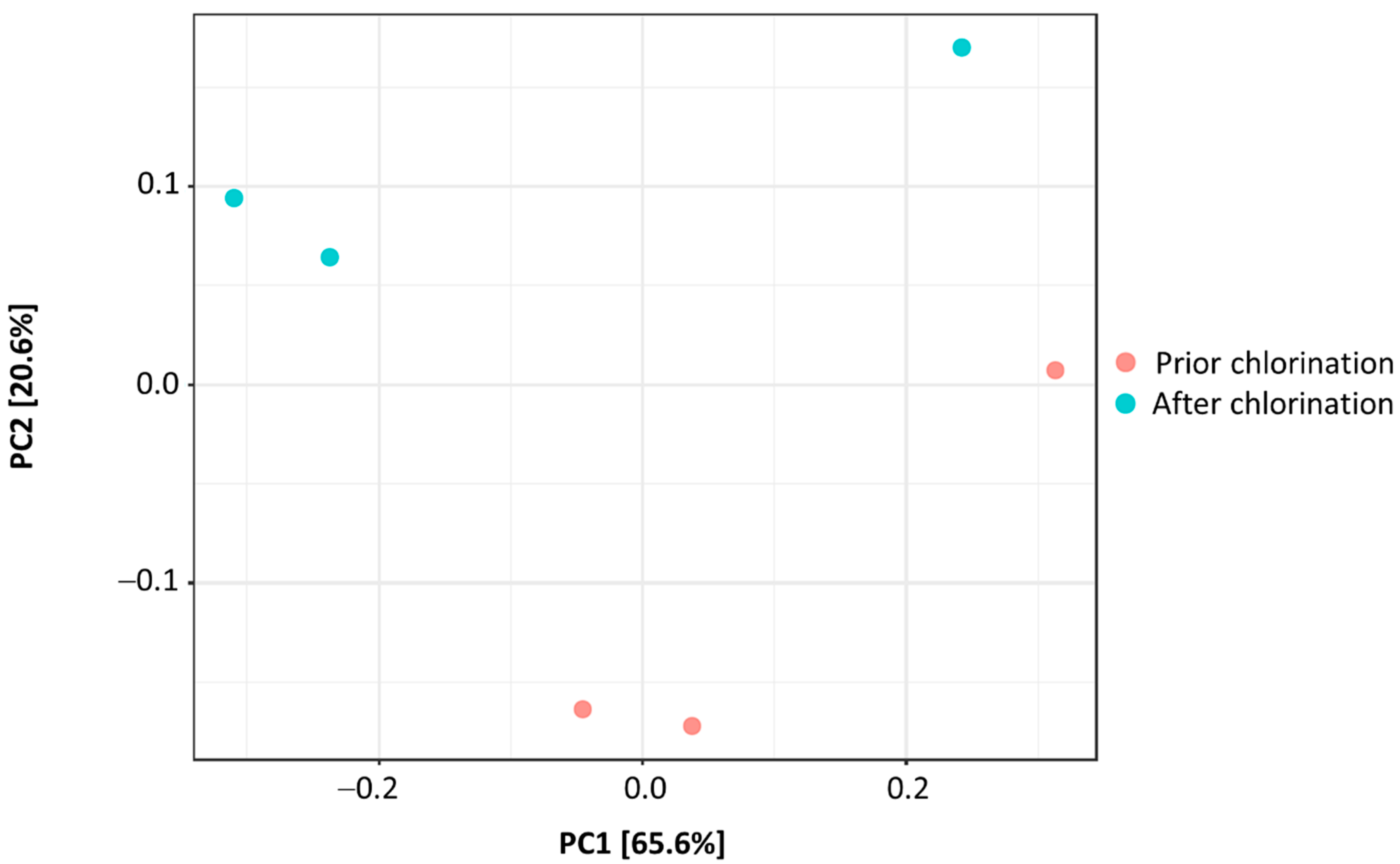
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Guadagnini, R.A.; Dos Santos, L.U.; Franco, R.M.B.; Guimarães, J.R. Inactivation of bacteria and helminth in wastewater treatment plant effluent using oxidation processes. Water Sci. Technol. 2013, 68, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Numberger, D.; Ganzert, L.; Zoccarato, L.; Mühldorfer, K.; Sauer, S.; Grossart, H.P.; Greenwood, A.D. Characterization of bacterial communities in wastewater with enhanced taxonomic resolution by full-length 16S rRNA sequencing. Sci. Rep. 2019, 9, 9673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greay, T.L.; Gofton, A.W.; Zahedi, A.; Paparini, A.; Linge, K.L.; Joll, C.A.; Ryan, U.M. Evaluation of 16S next-generation sequencing of hypervariable region 4 in wastewater samples: An unsuitable approach for bacterial enteric pathogen identification. Sci. Total Environ. 2019, 670, 1111–1124. [Google Scholar] [CrossRef] [PubMed]
- Beattie, R.E.; Skwor, T.; Hristova, K.R. Survivor microbial populations in post-chlorinated wastewater are strongly associated with untreated hospital sewage and include ceftazidime and meropenem resistant populations. Sci. Total Environ. 2020, 740, 140186. [Google Scholar] [CrossRef]
- Wolff, D.; Krah, D.; Dötsch, A.; Ghattas, A.K.; Wick, A.; Ternes, T.A. Insights into the variability of microbial community composition and micropollutant degradation in diverse biological wastewater treatment systems. Water Res. 2018, 143, 313–324. [Google Scholar] [CrossRef]
- Sousa, J.M.; Macedo, G.; Pedrosa, M.; Becerra-Castro, C.; Castro-Silva, S.; Pereira, M.F.R.; Silva, A.M.; Nunes, O.C.; Manaia, C.M. Ozonation and UV254 nm radiation for the removal of microorganisms and antibiotic resistance genes from urban wastewater. J. Hazard. Mater. 2017, 323, 434–441. [Google Scholar] [CrossRef]
- Yao, X.; Hu, X.; Liu, Y.; Wang, X.; Hong, X.; Chen, X.; Pillai, S.C.; Dionysiou, D.D.; Wang, D. Simultaneous photocatalytic degradation of ibuprofen and H2 evolution over Au/sheaf-like TiO2 mesocrystals. Chemosphere 2020, 261, 127759. [Google Scholar] [CrossRef]
- Pang, Y.C.; Xi, J.Y.; Xu, Y.; Huo, Z.Y.; Hu, H.Y. Shifts of live bacterial community in secondary effluent by chlorine disinfection revealed by Miseq high-throughput sequencing combined with propidium monoazide treatment. Appl. Microbiol. Biotechnol. 2016, 100, 6435–6446. [Google Scholar] [CrossRef]
- Francy, D.S.; Stelzer, E.A.; Bushon, R.N.; Brady, A.M.G.; Williston, A.G.; Riddell, K.R.; Borchardt, M.A.; Spencer, S.K.; Gellner, T.M. Comparative effectiveness of membrane bioreactors, conventional secondary treatment, and chlorine and UV disinfection to remove microorganisms from municipal wastewaters. Water Res. 2012, 46, 4164–4178. [Google Scholar] [CrossRef]
- Papadimitriou, T.; Kormas, K.; Dionysiou, D.D.; Laspidou, C. Using H2O2 treatments for the degradation of cyanobacteria and microcystins in a shallow hypertrophic reservoir. Environ. Sci. Pollut. Res. 2016, 23, 21523–21535. [Google Scholar] [CrossRef]
- Nasuhoglu, D.; Isazadeh, S.; Westlund, P.; Neamatallah, S.; Yargeau, V. Chemical, microbial and toxicological assessment of wastewater treatment plant effluents during disinfection by ozonation. Chem. Eng. J. 2018, 346, 466–476. [Google Scholar] [CrossRef]
- Pedersen, P.O.; Brodersen, E.; Cecil, D. Disinfection of tertiary wastewater effluent prior to river discharge using peracetic acid; treatment efficiency and results on by-products formed in full scale tests. Water Sci. Technol. 2013, 68, 1852–1856. [Google Scholar] [CrossRef] [PubMed]
- Hassaballah, A.H.; Nyitrai, J.; Hart, C.H.; Dai, N.; Sassoubre, L.M. A pilot-scale study of peracetic acid and ultraviolet light for wastewater disinfection. Environ. Sci. Water Res. Technol. 2019, 5, 1453–1463. [Google Scholar] [CrossRef]
- Valero, P.; Verbel, M.; Silva-Agredo, J.; Mosteo, R.; Ormad, M.P.; Torres-Palma, R.A. Electrochemical advanced oxidation processes for Staphylococcus aureus disinfection in municipal WWTP effluents. J. Environ. Manag. 2017, 198, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cheng, D.; Li, Y.; Yu, L.; Gin, K.Y.H.; Chen, J.P.; Reinhard, M. Effects of monochloramine and hydrogen peroxide on the bacterial community shifts in biologically treated wastewater. Chemosphere 2017, 189, 399–406. [Google Scholar] [CrossRef]
- Di Cesare, A.; De Carluccio, M.; Eckert, E.M.; Fontaneto, D.; Fiorentino, A.; Corno, G.; Prete, P.; Cucciniello, R.; Proto, A.; Rizzo, L. Combination of flow cytometry and molecular analysis to monitor the effect of UVC/H2O2 vs UVC/H2O2/Cu-IDS processes on pathogens and antibiotic resistant genes in secondary wastewater effluents. Water Res. 2020, 184, 116194. [Google Scholar] [CrossRef]
- Martínez-Hernández, S.; Vázquez-Rodríguez, G.A.; Beltrán-Hernández, R.I.; Prieto-García, F.; Miranda-López, J.M.; Franco-Abuín, C.M.; Álvarez-Hernández, A.; Iturbe, U.; Coronel-Olivares, C. Resistance and inactivation kinetics of bacterial strains isolated from the non-chlorinated and chlorinated effluents of a WWTP. Int. J. Environ. Res. Public Health 2013, 10, 3363–3383. [Google Scholar] [CrossRef]
- Alexander, J.; Karaolia, P.; Fatta-Kassinos, D.; Schwartz, T. Impacts of advanced oxidation processes on microbiomes during wastewater treatment. In Advanced Treatment Technologies for Urban Wastewater Reuse-The Handbook of Environmental Chemistry; Fatta-Kassinos, D., Dionysiou, D., Kümmerer, K., Eds.; Springer: Cham, Switzerland, 2015; Volume 45, pp. 129–144. [Google Scholar]
- Kulkarni, P.; Olson, N.D.; Paulson, J.N.; Pop, M.; Maddox, C.; Claye, E.; Rosenberg Goldstein, R.E.; Sharma, M.; Gibbs, S.G.; Mongodin, E.F.; et al. Conventional wastewater treatment and reuse site practices modify bacterial community structure but do not eliminate some opportunistic pathogens in reclaimed water. Sci. Total Environ. 2018, 639, 1126–1137. [Google Scholar] [CrossRef]
- Nasser, A.M.; Benisti, N.L.; Ofer, N.; Hovers, S.; Nitzan, Y. Comparative reduction of Giardia cysts, F+ coliphages, sulphite reducing clostridia and fecal coliforms by wastewater treatment processes. J. Environ. Sci. Health A 2017, 52, 144–148. [Google Scholar] [CrossRef]
- Lin, Y.; Li, D.; Zeng, S.; He, M. Changes of microbial composition during wastewater reclamation and distribution systems revealed by high-throughput sequencing analyses. Front. Environ. Sci. Eng. 2016, 10, 539–547. [Google Scholar] [CrossRef]
- Li, D.; Zeng, S.; Gu, A.Z.; He, M.; Shi, H. Inactivation, reactivation and regrowth of indigenous bacteria in reclaimed water after chlorine disinfection of a municipal wastewater treatment plant. J. Environ. Sci. 2013, 25, 1319–1325. [Google Scholar] [CrossRef]
- Park, E.; Mancl, K.M.; Tuovinen, O.H.; Bisesi, M.S.; Lee, J. Ensuring safe reuse of residential wastewater: Reduction of microbes and genes using peat biofilter and batch chlorination in an on-site treatment system. J. Appl. Microbiol. 2016, 121, 1777–1788. [Google Scholar] [CrossRef] [PubMed]
- Limayem, A.; Wasson, S.; Mehta, M.; Pokhrel, A.R.; Patil, S.; Nguyen, M.; Chen, J.; Nayak, B. High-throughput detection of bacterial community and its drug-resistance profiling from local reclaimed wastewater plants. Front. Cell Infect. Microbiol. 2019, 9, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater; American Public Health Association (APHA): Washington, DC, USA, 1998. [Google Scholar]
- Remmas, N.; Roukouni, C.; Ntougias, S. Bacterial community structure and prevalence of Pusillimonas-like bacteria in aged landfill leachate. Environ. Sci. Pollut. Res. 2017, 24, 6757–6769. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for Illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016, 081257. [Google Scholar] [CrossRef] [Green Version]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Hammer, Ø.; (Natural History Museum, University of Oslo, Olso, Norway). PAST-Paleontological Statistics; Version 3.25., 2019. Available online: https://folk.uio.no/ohammer/past/index.html (accessed on 18 January 2021).
- Bardiya, N.; Bae, J.H. Dissimilatory perchlorate reduction: A review. Microbiol. Res. 2011, 166, 237–254. [Google Scholar] [CrossRef]
- DuBois, J.L.; Ojha, S. Production of dioxygen in the dark: Dismutases of oxyanions. Met. Ions Life Sci. 2015, 15, 45–87. [Google Scholar] [CrossRef]
- Zhu, Z.; Shan, L.; Hu, F.; Li, Z.; Zhong, D.; Yuan, Y.; Zhang, J. Biofilm formation potential and chlorine resistance of typical bacteria isolated from drinking water distribution systems. RSC Adv. 2020, 10, 31295–31304. [Google Scholar] [CrossRef]
- Mao, G.; Song, Y.; Bartlam, M.; Wang, Y. Long-term effects of residual chlorine on Pseudomonas aeruginosa in simulated drinking water fed with low AOC medium. Front. Microbiol. 2018, 9, 879. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Shi, P.; Hu, Q.; Li, B.; Zhang, T.; Zhang, X.X. Bacterial community shift drives antibiotic resistance promotion during drinking water chlorination. Environ. Sci. Technol. 2015, 49, 12271–12279. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- An, W.; Guo, F.; Song, Y.; Gao, N.; Bai, S.; Dai, J.; Wei, H.; Zhang, L.; Yu, D.; Xia, M.; et al. Comparative genomics analyses on EPS biosynthesis genes required for floc formation of Zoogloea resiniphila and other activated sludge bacteria. Water Res. 2016, 102, 494–504. [Google Scholar] [CrossRef]
- Durán, A.E.; Muniesa, M.; Mocé-Llivina, L.; Campos, C.; Jofre, J.; Lucena, F. Usefulness of different groups of bacteriophages as model micro-organisms for evaluating chlorination. J. Appl. Microbiol. 2003, 95, 29–37. [Google Scholar] [CrossRef]
- Towhid, S.T. Microbial interaction as a determinant of the quality of supply drinking water: A conceptual analysis. Front. Public Health 2018, 6, 184. [Google Scholar] [CrossRef]
- Li, C.; Dong, F.; Feng, L.; Zhao, J.; Zhang, T.; Cizmas, L.; Sharma, V.K. Bacterial community structure and microorganism inactivation following water treatment with ferrate(VI) or chlorine. Environ. Chem. Lett. 2017, 15, 525–530. [Google Scholar] [CrossRef]
- Wang, O.; Coates, J.D. Biotechnological applications of microbial (per)chlorate reduction. Microorganisms 2017, 5, 76. [Google Scholar] [CrossRef] [Green Version]
- Mehta-Kolte, M.G.; Loutey, D.; Wang, O.; Youngblut, M.D.; Hubbard, C.G.; Wetmore, K.M.; Conrad, M.E.; Coates, J.D. Mechanism of H2S oxidation by the dissimilatory perchlorate-reducing microorganism Azospira suillum PS. mBio 2017, 8, e02023-16. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Low, A.; Zemb, O.; Koenig, J.; Michaelsen, A.; Manefield, M. Complete chloroform dechlorination by organochlorine respiration and fermentation. Environ. Microbiol. 2012, 14, 883–894. [Google Scholar] [CrossRef]
- Hug, L.A.; Maphosa, F.; Leys, D.; Löffler, F.E.; Smidt, H.; Edwards, E.A.; Adrian, L. Overview of organohalide-respiring bacteria and a proposal for a classification system for reductive dehalogenases. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jugder, B.E.; Ertan, H.; Bohl, S.; Lee, M.; Marquis, C.P.; Manefield, M. Organohalide respiring bacteria and reductive dehalogenases: Key tools in organohalide bioremediation. Front. Microbiol. 2016, 7, 249. [Google Scholar] [CrossRef] [PubMed]
- Krzmarzick, M.J.; Crary, B.B.; Harding, J.J.; Oyerinde, O.O.; Leri, A.C.; Myneni, S.C.B.; Novak, P.J. Natural niche for organohalide-respiring Chloroflexi. Appl. Environ. Microbiol. 2012, 78, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.; Tian, Z.; Yang, H.; Yang, M.; Zhang, Y. Detection of viable bacteria during sludge ozonation by the combination of ATP assay with PMA-miseq sequencing. Water 2017, 9, 166. [Google Scholar] [CrossRef]
- Eichler, S.; Christen, R.; Höltje, C.; Westphal, P.; Bötel, J.; Brettar, I.; Mehling, A.; Höfle, M.G. Composition and dynamics of bacterial communities of a drinking water supply system as assessed by RNA- and DNA-based 16S rRNA gene fingerprinting. Appl. Environ. Microbiol. 2006, 72, 1858–1872. [Google Scholar] [CrossRef] [Green Version]
- Li, B.B.; Peng, Z.Y.; Zhi, L.L.; Li, H.B.; Zheng, K.K.; Li, J. Distribution and diversity of filamentous bacteria in wastewater treatment plants exhibiting foaming of Taihu Lake Basin, China. Environ. Pollut. 2020, 267, 115644. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Z.; Song, J.; Cai, L.; Yu, Y.; Fang, H. Foam shares antibiotic resistomes and bacterial pathogens with activated sludge in wastewater treatment plants. J. Hazard. Mater. 2021, 408, 124855. [Google Scholar] [CrossRef]
- Caravelli, A.; Giannuzzi, L.; Zaritzky, N. Effect of chlorine on filamentous microorganisms present in activated sludge as evaluated by respirometry and INT-dehydrogenase activity. Water Res. 2004, 38, 2395–2405. [Google Scholar] [CrossRef]
- Liu, J.; Ren, H.; Ye, X.; Wang, W.; Liu, Y.; Lou, L.; Cheng, D.; He, X.; Zhou, X.; Qiu, S.; et al. Bacterial community radial-spatial distribution in biofilms along pipe wall in chlorinated drinking water distribution system of East China. Appl. Microbiol. Biotechnol. 2017, 101, 749–759. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef] [Green Version]
- Catherine, Q.; Susanna, W.; Isidora, E.S.; Mark, H.; Aurélie, V.; Jean-François, H. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef] [PubMed]
- Chia, M.A.; Jankowiak, J.G.; Kramer, B.J.; Goleski, J.A.; Huang, I.S.; Zimba, P.V.; do Carmo Bittencourt-Oliveira, M.; Gobler, C.J. Succession and toxicity of Microcystis and Anabaena (Dolichospermum) blooms are controlled by nutrient-dependent allelopathic interactions. Harmful Algae 2018, 74, 67–77. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Influent | Effluent | Removal (%) |
|---|---|---|---|
| BOD5 (mg/L) | 363 ± 5 | 17.3 ± 0.5 | 95.2 ± 0.1 |
| COD (mg/L) | 626 ± 17 | 67.1 ± 2.4 | 89.1 ± 0.4 |
| TSS (mg/L) | 311 ± 7 | 18.9 ± 0.7 | 93.8 ± 0.2 |
| NO3−-N (mg/L) | - | 9.4 ± 0.5 | - |
| Genus | Prior to Chlorination | After Chlorination | Significance |
|---|---|---|---|
| Anaerolinea | 0.02 ± 0.01 | 0.01 ± 0.01 | p < 0.01 |
| Bellilinea | 1.42 ± 0.70 | 0.80 ± 0.34 | n.s. |
| Caldilinea | 2.19 ± 0.96 | 1.07 ± 0.52 | n.s. |
| Dehalobacter | 0.01 ± 0.01 | 0.01 ± 0.01 | n.s. |
| Dehalobacterium | At detection limit | At detection limit | n.s. |
| Dehalococcoides | 0.37 ± 0.03 | 0.24 ± 0.06 | n.s. |
| Dehalogenimonas | n.d. | At detection limit | n.s. |
| Dechloromonas | 3.17 ± 0.21 | 1.96 ± 0.36 | p < 0.05 |
| Levilinea | 0.02 ± 0.01 | 0.01 ± 0.01 | p < 0.05 |
| Longilinea | 1.25 ± 0.26 | 1.19 ± 0.35 | n.s. |
| Genus | Prior to Chlorination | After Chlorination | Significance |
|---|---|---|---|
| Candidatus Nitrotoga | 0.03 ± 0.02 | 0.01 ± 0.01 | n.s. |
| Nitrobacter | At detection limit | At detection limit | n.s. |
| Nitrosomonas | 0.23 ± 0.20 | 0.04 ± 0.03 | n.s. |
| Nitrospira | 0.10 ± 0.03 | 0.18 ± 0.05 | n.s. |
| Nitrosovibrio | 0.27 ± 0.03 | n.d. | p < 0.01 |
| Genus | Prior to Chlorination | After Chlorination | Significance |
|---|---|---|---|
| Bacterial Taxa Influencing Enhanced Phosphorus Removal | |||
| Candidatus Accumulibacter | 3.31 ± 0.29 | 2.37 ± 0.63 | n.s. |
| Candidatus Competibacter | 1.02 ± 0.06 | 1.02 ± 0.17 | n.s. |
| Annamox bacteria | |||
| Candidatus Anammoximicrobium | At detection limit | 0.01 ± 0.01 | n.s. |
| Candidatus Kuenenia | At detection limit | 0.01 ± 0.01 | n.s. |
| Genus | Prior to Chlorination | After Chlorination | Significance |
|---|---|---|---|
| Beggiatoa | 0.06 ± 0.03 | 0.03 ± 0.02 | n.s. |
| Candidatus Microthrix | 0.47 ± 0.21 | 0.08 ± 0.04 | n.s. |
| Gordonia | 0.47 ± 0.05 | 0.21 ± 0.04 | p < 0.05 |
| Haliscomenobacter | 0.77 ± 0.10 | 0.43 ± 0.15 | n.s. |
| Nocardioides | 0.96 ± 0.10 | 0.19 ± 0.04 | p < 0.01 |
| Nostocoida Type II | At detection limit | n.d. | n.s. |
| Thiothrix | 0.06 ± 0.01 | 0.02 ± 0.01 | n.s. |
| Zoogloea | 0.42 ± 0.02 | 0.64 ± 0.08 | n.s. |
| Genus | Prior Chlorination | After Chlorination | Significance |
|---|---|---|---|
| Brucella | 0.03 ± 0.02 | At detection limit | n.s. |
| Campylobacter | 0.02 ± 0.01 | 0.04 ± 0.02 | n.s. |
| Enterococcus | 0.01 ± 0.01 | 0.01 ± 0.01 | n.s. |
| Klebsiella | n.d. | At detection limit | n.s. |
| Legionella | 0.01 ± 0.01 | 0.05 ± 0.04 | n.s. |
| Mycobacterium | 0.15 ± 0.05 | n.d. | p < 0.05 |
| Rickettsia | 0.03 ± 0.01 | 0.40 ± 0.37 | n.s. |
| Shigella | At detection limit | n.d. | n.s. |
| Spirochaeta | 0.04 ± 0.01 | 0.07 ± 0.04 | n.s. |
| Treponema | 0.09 ± 0.02 | 0.05 ± 0.00 | n.s. |
| Vibrio | At detection limit | 0.45 ± 0.45 | n.s. |
| Genus | Prior to Chlorination | After Chlorination | Significance |
|---|---|---|---|
| Anabaena | n.d. | 0.04 ± 0.01 | p < 0.01 |
| Arthrospira | n.d. | At detection limit | n.s. |
| Cyanobacterium | At detection limit | 0.01 ± 0.01 | n.s. |
| Cyanothece | n.d. | 0.01 ± 0.01 | p < 0.01 |
| Elstera | At detection limit | At detection limit | n.s. |
| Gloeobacter | 0.08 ± 0.04 | 0.08 ± 0.03 | n.s. |
| Leptolyngbya | 0.24 ± 0.16 | 0.04 ± 0.02 | n.s. |
| Microcystis | 0.03 ± 0.01 | 0.02 ± 0.00 | n.s. |
| Oscillatoria | At detection limit | At detection limit | n.s. |
| Phormidium | At detection limit | At detection limit | n.s. |
| Prochlorococcus | 0.03 ± 0.01 | 0.03 ± 0.01 | n.s. |
| Pseudanabaena | At detection limit | At detection limit | n.s. |
| Vampirovibrio | 0.01 ± 0.01 | At detection limit | n.s. |
| Disinfection Method | Susceptible Taxa | Resistant Taxa | Reference |
|---|---|---|---|
| Monochloramine | Arcobacter, Nitrospira, Sphingobium | Chryseobacterium, Cloacibacterium, Clostridium, Mycobacterium, Pseudomonas, Sphingomonas, Streptococcus, Undibacterium | [15] |
| H2O2 | Denitratisoma, Thauera | Anaerolinea, Filimonas | [15] |
| UV 1 | Acetoanaerobium, Acidovorax, Acinetobacter, Aeromonas, Anaerosinus, Aquabacterium, Arcobacter, Comamonas, Enterococcus, Faecalibacterium, Paracoccus, Proteocatella, Streptococcus, Subdoligranulum, Trichococcus, Uruburuella, Veillonella | Ca. Accumulibacter, Ca. Competibacter, Ca. Nitrotoga, Chryseobacterium, Cupriavidus, Dechloromonas, Geothrix, Nitrosomonas, Nitrospira, Rhodoferax, Simplicispira, Thauera, Zoogloea | [2] |
| UV-C/H2O2/IDS-Cu 2 | N.R. | Acinetobacter, Pantoea, Pseudomonas | [16] |
| Chlorination 4 | Acinetobacter, Arcobacter, Azonexus, Azospira, Bifidobacterium, Chitinimonas, Comamonas, Dechloromonas, Enterococcus, Lactobacillales, Laribacter, Neisseria, Nitrosomonas, Propionivibrio, Rheinheimera, Salmonella, Shewanella, Thauera, Tolumonas, Vitreoscilla, Zoogloea | Aquabacterium, Chryseobacterium, Clostridium, Flavobacterium, Gemmata, Hydrogenophaga, Legionella, Mycobacterium, Pseudomonas, Rubrivivax, Turicibacter, Veillonella | [3,8] |
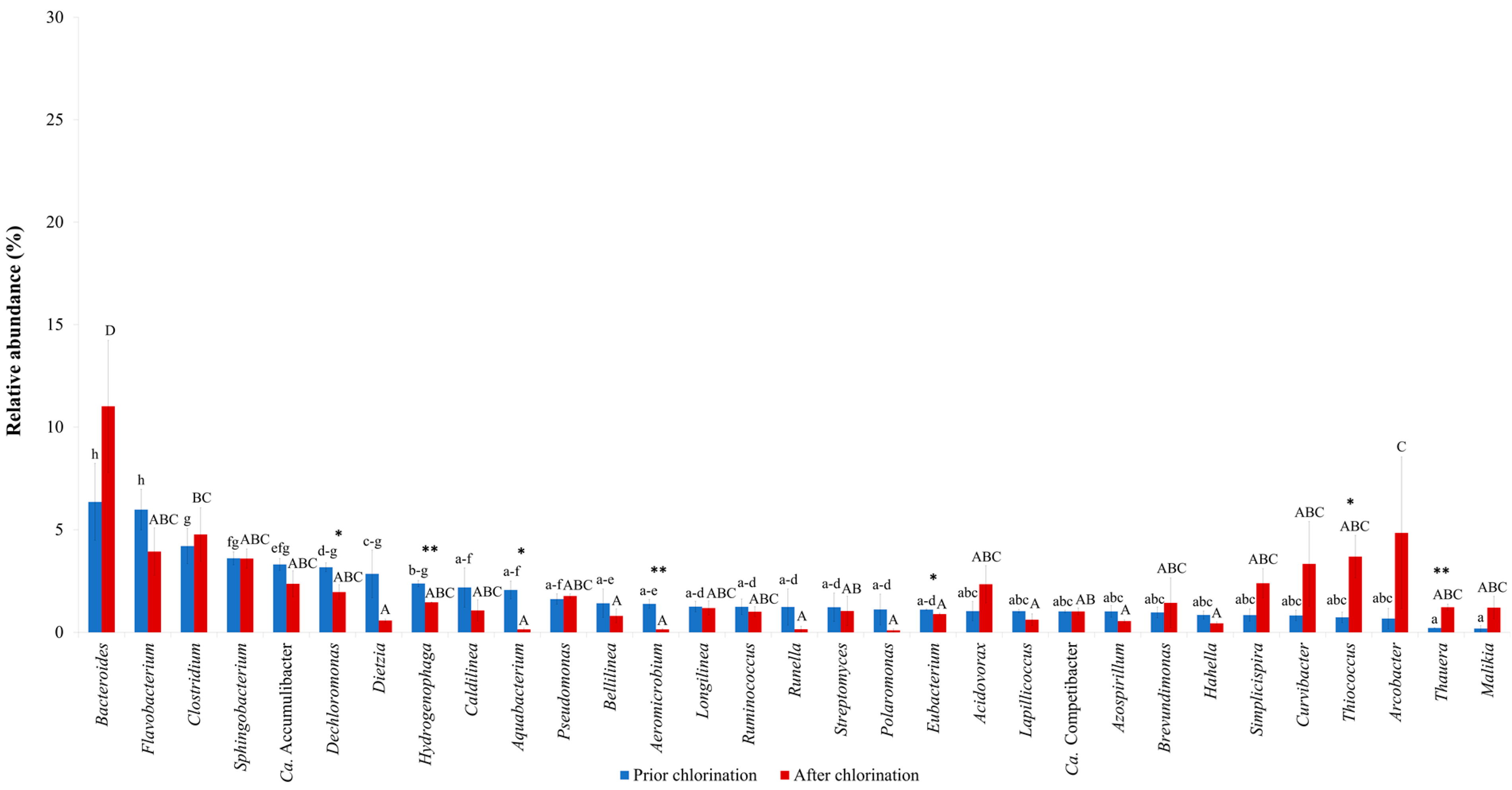
| Chlorination | Aeromicrobium, Aquabacterium, Dechloromonas, Eubacterium, Hydrogenophaga3 | Acidovorax, Arcobacter, Azospirillum, Bacteroides, Bellilinea, Brevundimonas, Caldilinea, Ca. Accumulibacter, Ca. Competibacter, Clostridium, Curvibacter, Dietzia, Flavobacterium, Hahella, Lapillicoccus, Longilinea, Malikia, Polaromonas, Pseudomonas, Ruminococcus, Runella, Simplicispira, Sphingobacterium, Streptomyces, Thauera, Thiococcus3 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zerva, I.; Remmas, N.; Kagalou, I.; Melidis, P.; Ariantsi, M.; Sylaios, G.; Ntougias, S. Effect of Chlorination on Microbiological Quality of Effluent of a Full-Scale Wastewater Treatment Plant. Life 2021, 11, 68. https://doi.org/10.3390/life11010068
Zerva I, Remmas N, Kagalou I, Melidis P, Ariantsi M, Sylaios G, Ntougias S. Effect of Chlorination on Microbiological Quality of Effluent of a Full-Scale Wastewater Treatment Plant. Life. 2021; 11(1):68. https://doi.org/10.3390/life11010068
Chicago/Turabian StyleZerva, Ioanna, Nikolaos Remmas, Ifigeneia Kagalou, Paraschos Melidis, Marina Ariantsi, Georgios Sylaios, and Spyridon Ntougias. 2021. "Effect of Chlorination on Microbiological Quality of Effluent of a Full-Scale Wastewater Treatment Plant" Life 11, no. 1: 68. https://doi.org/10.3390/life11010068
APA StyleZerva, I., Remmas, N., Kagalou, I., Melidis, P., Ariantsi, M., Sylaios, G., & Ntougias, S. (2021). Effect of Chlorination on Microbiological Quality of Effluent of a Full-Scale Wastewater Treatment Plant. Life, 11(1), 68. https://doi.org/10.3390/life11010068