
Mitochondrial DNA Analysis Clarifies Taxonomic Status of the Northernmost Snow Sheep (Ovis nivicola) Population

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bunch, T.D.; Wu, C.; Zhang, Y.-P.; Wang, S. Phylogenetic Analysis of Snow Sheep (Ovis nivicola) and Closely Related Taxa. J. Hered. 2006, 97, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revin, Y.V.; Sopin, L.V.; Zheleznov, N.K. Snow Sheep; Nauka: Novosibirsk, Russia, 1988. (In Russian) [Google Scholar]
- Zheleznov-Chukotsky, N.K. Ecology of Snow Sheep of Northern Asia; Nauka: Moscow, Russia, 1994. (In Russian) [Google Scholar]
- Chernyavsky, F.B. On taxonomy and history of snow sheep (subgenus Pachyceros, Artiodactyla). J. Zool. 2004, 83, 1059–1070. (In Russian) [Google Scholar]
- Danilkin, A.A. Mammals of Russia and Adjacent Regions: Hollow-Horned Ruminants (Bovidae); KMK: Moscow, Russia, 2005. (In Russian) [Google Scholar]
- Zheleznov-Chukotsky, N.K. New subspecies of snow sheep in Russian northern Asia. In 6th World Congress on Mountain Un-gulates and 5th International Symposium on Mouflon; Book of Abstracts; Ministry of the Interior: Nicosia, Cyprus, 2016; p. 95. [Google Scholar]
- Damm, G.R.; Franco, N. CIC Caprinae Atlas of the World; CIC International Council for Game and Wildlife Conservation: Budakeszi, Hungary; Rowland Ward Publications: Johannesburg, South Africa, 2014. [Google Scholar]
- Abramson, N.I. Molecular and conventional phylogenetics. Towards the common ground. Proc. Zool. Inst. Russ. Acad. Sci. 2013, 2, 219–229. (In Russian) [Google Scholar]
- Ropiquet, A.; Hassanin, A. Molecular evidence for the polyphyly of the genus Hemitragus (Mammalia, Bovidae). Mol. Phylogenetics Evol. 2005, 36, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Tokarska, M.; Marshall, T.; Kowalczyk, R.; Wójcik, J.M.; Pertoldi, C.; Kristensen, T.N.; Loeschcke, V.; Gregersen, V.R.; Bendixen, C. Effectiveness of microsatellite and SNP markers for parentage and identity analysis in species with low genetic diversity: The case of European bison. Heredity 2009, 103, 326–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharzinova, V.R.; Dotsev, A.V.; Deniskova, T.E.; Solovieva, A.D.; Fedorov, V.I.; Layshev, K.A.; Romanenko, T.M.; Okhlopkov, I.M.; Wimmers, K.; Reyer, H.; et al. Genetic diversity and population structure of domestic and wild reindeer (Rangifer tarandus L. 1758): A novel approach using BovineHD BeadChip. PLoS ONE 2018, 13, e0207944. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.; Poissant, J.; Kijas, J.; Coltman, D. The International Sheep Genomics Consortium. A genome-wide set of SNPs detects population substructure and long range linkage disequilibrium in wild sheep. Mol. Ecol. Resour. 2010, 11, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Dotsev, A.V.; Deniskova, T.E.; Okhlopkov, I.M.; Mészáros, G.; Sölkner, J.; Reyer, H.; Wimmers, K.; Brem, G.; Zinovieva, N.A. Genome-wide SNP analysis unveils genetic structure and phylogeographic history of snow sheep (Ovis nivicola) populations inhabiting the Verkhoyansk Mountains and Momsky Ridge (northeastern Siberia). Ecol. Evol. 2018, 8, 8000–8010. [Google Scholar] [CrossRef] [PubMed]
- Dotsev, A.V.; Kunz, E.; Shakhin, A.V.; Petrov, S.N.; Kostyunina, O.V.; Okhlopkov, I.M.; Deniskova, T.E.; Barbato, M.; Bagirov, V.A.; Medvedev, D.G.; et al. The first complete mitochondrial genomes of snow sheep (Ovis nivicola) and thinhorn sheep (Ovis dalli) and their phylogenetic implications for the genus Ovis. Mitochondrial DNA Part B 2019, 4, 1332–1333. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, H.R.; Naderi, S.; Chintauan-Marquier, I.C.; Taberlet, P.; Virk, A.T.; Naghash, H.R.; Rioux, D.; Kaboli, M.; Pompanon, F. Evolution and taxonomy of the wild species of the genus Ovis (Mammalia, Artiodactyla, Bovidae). Mol. Phylogenet. Evol. 2010, 54, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Naidu, A.; Fitak, R.R.; Munguia-Vega, A.; Culver, M. Novel primers for complete mitochondrial cytochrome b gene sequencing in mammals. Mol. Ecol. Resour. 2012, 12, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2005, 23, 254–267. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Fu, Y.-X. Statistical Tests of Neutrality of Mutations against Population Growth, Hitchhiking and Background Selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Becker, R.A.; Wilks, A.R.; Brownrigg, R.; Minka, T.P.; Deckmyn, A. Maps: Draw Geographical Maps; R Package Version 3.3.0. 2018. Available online: https://CRAN.R-project.org/package=maps (accessed on 22 December 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2009; p. 213. [Google Scholar] [CrossRef]
- Fritz, S.A.; Bininda-Emonds, O.R.P.; Purvis, A. Geographical variation in predictors of mammalian extinction risk: Big is bad, but only in the tropics. Ecol. Lett. 2009, 12, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, A.M.; Barraclough, T.G. The evolutionary reality of higher taxa in mammals. Proc. R. Soc. B Boil. Sci. 2014, 281, 20132750. [Google Scholar] [CrossRef] [Green Version]
- Bagirov, V.A.; Okhlopkov, I.M.; Zinovieva, N.A. Yakut Snow Sheep: Genetic Diversity and Ways of Conservation of the Gene Pool; Publishing House of L.K. Ernst Federal Research Center for Animal Husbandry: Dubrovitsy, Russia, 2016. (In Russian) [Google Scholar]






{kind=link}
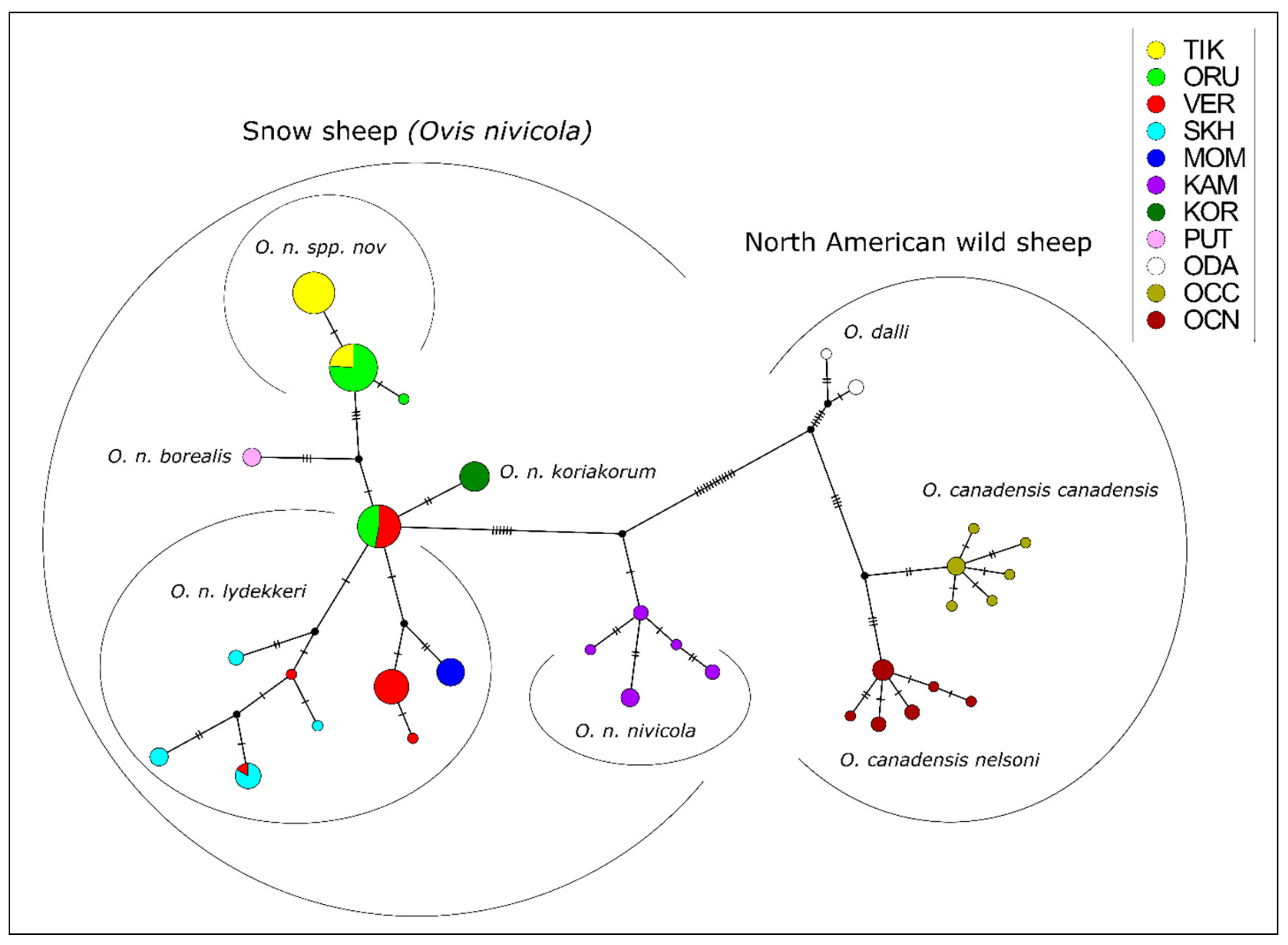{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Species | n | GenBank Accession Number | References |
|---|---|---|---|---|
| 1 | Rocky Mountain bighorn sheep (Ovis canadensis canadensis) | 8 | EU365985, EU366063, EU366064, EU366065, EU366066, EU366067, FJ936176, FJ936177 | Rezaei H.R. et al. [18] |
| 2 | Desert bighorn sheep (Ovis canadensis nelsoni) | 11 | EU366059, EU366060, EU366061, EU366062, FJ936178, FJ936179, FJ936180, FJ936181, FJ936182, FJ936183, | Rezaei H.R. et al. [18] |
| HM222706 | Naidu A. et al. [19] | |||
| 3 | Dall sheep (Ovis dalli) | 3 | MH779627 | Dotsev A. et al. [14] |
| EU365992, FJ936184 | Rezaei H.R. et al. [18] |
| Population | n | S | K (±SD) | H | Hd (±SD) | π (±SD) | Tajima’s D | Fu’s Fs |
|---|---|---|---|---|---|---|---|---|
| TIK | 21 | 1 | 0.381 ± 0.375 | 2 | 0.381 ± 0.101 | 0.00033± 0.00037 | 0.65593 | 0.94374 |
| ORU | 25 | 5 | 1.893 ± 1.117 | 3 | 0.507 ± 0.075 | 0.00166 ± 0.00109 | 1.23135 | 3.39988 |
| VER | 23 | 7 | 1.636 ± 1.002 | 5 | 0.640 ± 0.065 | 0.00144 ± 0.00098 | −0.43378 | 0.34957 |
| SKH | 11 | 8 | 3.018 ± 1.703 | 4 | 0.746 ± 0.098 | 0.00265 ± 0.00169 | 0.43451 | 1.79212 |
| MOM | 7 | 0 | 0 | 1 | 0 | 0 | - | - |
| KAM | 9 | 7 | 2.722 ± 1.592 | 5 | 0.861 ± 0.087 | 0.00239 ± 0.00158 | 0.25402 | −0.16693 |
| KOR | 8 | 0 | 0 | 1 | 0 | 0 | - | - |
| PUT | 3 | 0 | 0 | 1 | 0 | 0 | - | - |
| Population | TIK | ORU | VER | SKH | MOM | KOR | KAM | PUT | OCC | OCN | ODA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TIK | * | 0.002 | 0.005 | 0.008 | 0.005 | 0.006 | 0.012 | 0.006 | 0.025 | 0.022 | 0.025 |
| ORU | 0.438 | * | 0.004 | 0.006 | 0.004 | 0.004 | 0.010 | 0.005 | 0.024 | 0.021 | 0.024 |
| VER | 0.831 | 0.570 | * | 0.004 | 0.003 | 0.003 | 0.009 | 0.005 | 0.023 | 0.021 | 0.024 |
| SKH | 0.850 | 0.661 | 0.560 | * | 0.006 | 0.005 | 0.011 | 0.007 | 0.025 | 0.025 | 0.028 |
| MOM | 0.950 | 0.689 | 0.651 | 0.741 | * | 0.004 | 0.010 | 0.006 | 0.024 | 0.022 | 0.025 |
| KOR | 0.959 | 0.718 | 0.665 | 0.714 | 1.000 | * | 0.009 | 0.005 | 0.023 | 0.021 | 0.024 |
| KAM | 0.919 | 0.813 | 0.804 | 0.771 | 0.868 | 0.865 | * | 0.011 | 0.020 | 0.020 | 0.021 |
| PUT | 0.950 | 0.704 | 0.741 | 0.708 | 1.000 | 1.000 | 0.834 | * | 0.026 | 0.025 | 0.028 |
| OCC | 0.972 | 0.924 | 0.933 | 0.913 | 0.967 | 0.968 | 0.904 | 0.958 | * | 0.012 | 0.013 |
| OCN | 0.972 | 0.934 | 0.941 | 0.926 | 0.966 | 0.967 | 0.912 | 0.959 | 0.769 | * | 0.006 |
| ODA | 0.981 | 0.929 | 0.935 | 0.899 | 0.981 | 0.982 | 0.884 | 0.966 | 0.876 | 0.886 | * |
| Source of Variation | d.f. | SS | VC | V% |
|---|---|---|---|---|
| Among populations | 3 | 105.035 | 2.36458 | 78.8 |
| Within populations | 58 | 36.9 | 0.63621 | 21.2 |
| Total | 61 | 141.935 | 3.00080 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dotsev, A.V.; Kunz, E.; Kharzinova, V.R.; Okhlopkov, I.M.; Lv, F.-H.; Li, M.-H.; Rodionov, A.N.; Shakhin, A.V.; Sipko, T.P.; Medvedev, D.G.; et al. Mitochondrial DNA Analysis Clarifies Taxonomic Status of the Northernmost Snow Sheep (Ovis nivicola) Population. Life 2021, 11, 252. https://doi.org/10.3390/life11030252
Dotsev AV, Kunz E, Kharzinova VR, Okhlopkov IM, Lv F-H, Li M-H, Rodionov AN, Shakhin AV, Sipko TP, Medvedev DG, et al. Mitochondrial DNA Analysis Clarifies Taxonomic Status of the Northernmost Snow Sheep (Ovis nivicola) Population. Life. 2021; 11(3):252. https://doi.org/10.3390/life11030252
Chicago/Turabian StyleDotsev, Arsen V., Elisabeth Kunz, Veronika R. Kharzinova, Innokentiy M. Okhlopkov, Feng-Hua Lv, Meng-Hua Li, Andrey N. Rodionov, Alexey V. Shakhin, Taras P. Sipko, Dmitry G. Medvedev, and et al. 2021. "Mitochondrial DNA Analysis Clarifies Taxonomic Status of the Northernmost Snow Sheep (Ovis nivicola) Population" Life 11, no. 3: 252. https://doi.org/10.3390/life11030252
APA StyleDotsev, A. V., Kunz, E., Kharzinova, V. R., Okhlopkov, I. M., Lv, F.-H., Li, M.-H., Rodionov, A. N., Shakhin, A. V., Sipko, T. P., Medvedev, D. G., Gladyr, E. A., Bagirov, V. A., Brem, G., Medugorac, I., & Zinovieva, N. A. (2021). Mitochondrial DNA Analysis Clarifies Taxonomic Status of the Northernmost Snow Sheep (Ovis nivicola) Population. Life, 11(3), 252. https://doi.org/10.3390/life11030252