
The Mitochondrial Genome of the Sea Anemone Stichodactyla haddoni Reveals Catalytic Introns, Insertion-Like Element, and Unexpected Phylogeny


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Collection and Nucleic Acid Isolation
2.2. DNA and RNA Sequencing
2.3. Mitogenome Assembly and Annotation
2.4. Mitochondrial Transcriptome
2.5. Phylogenetic Analysis
3. Results
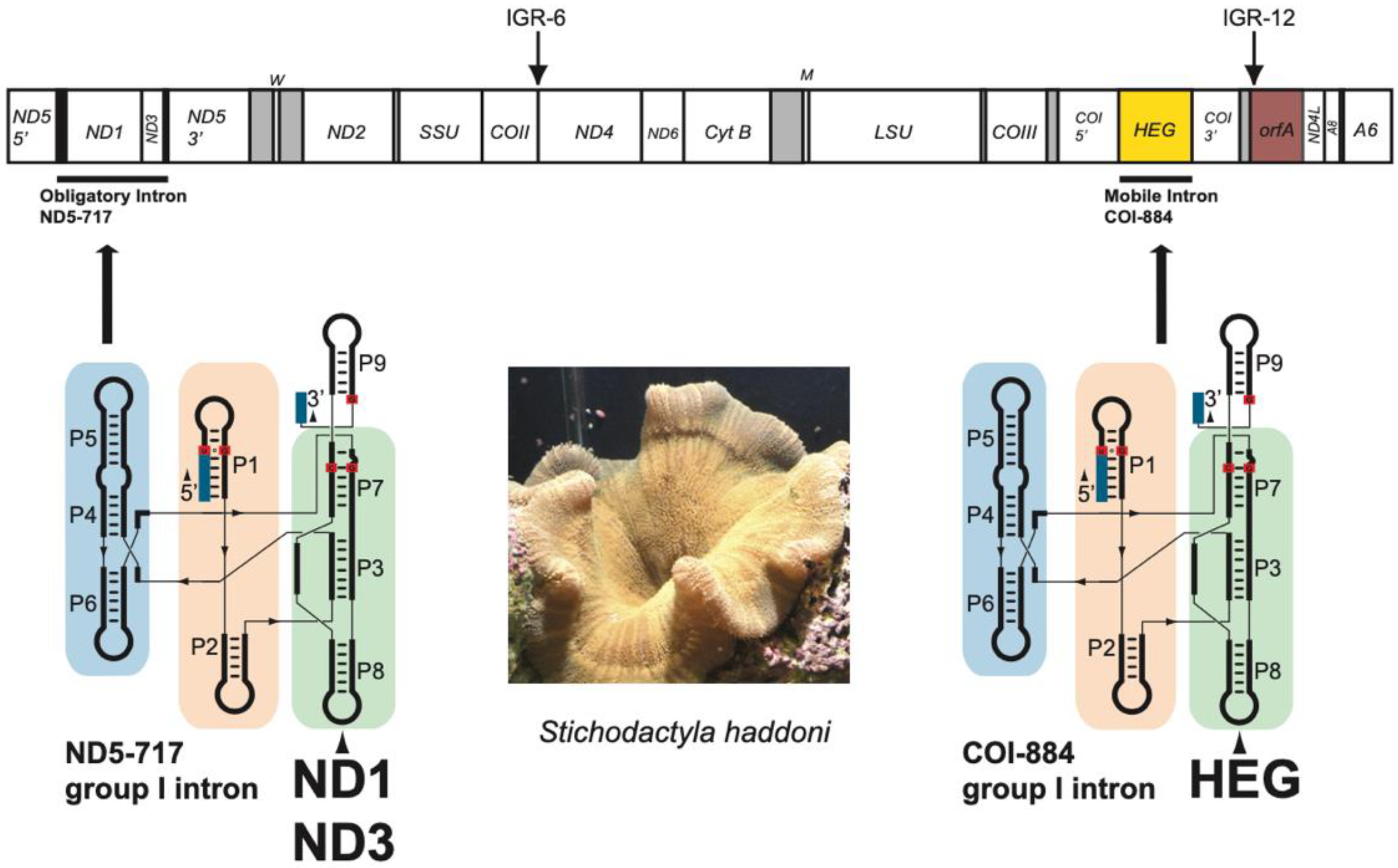
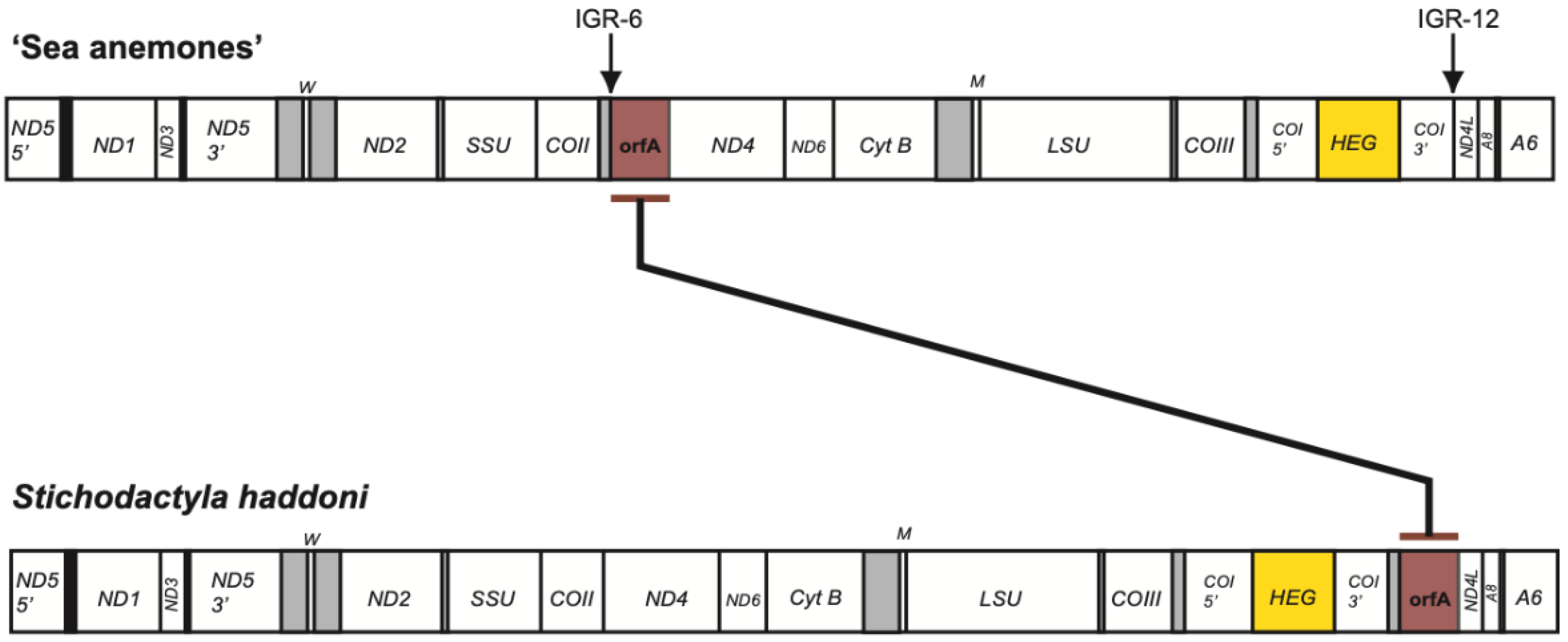
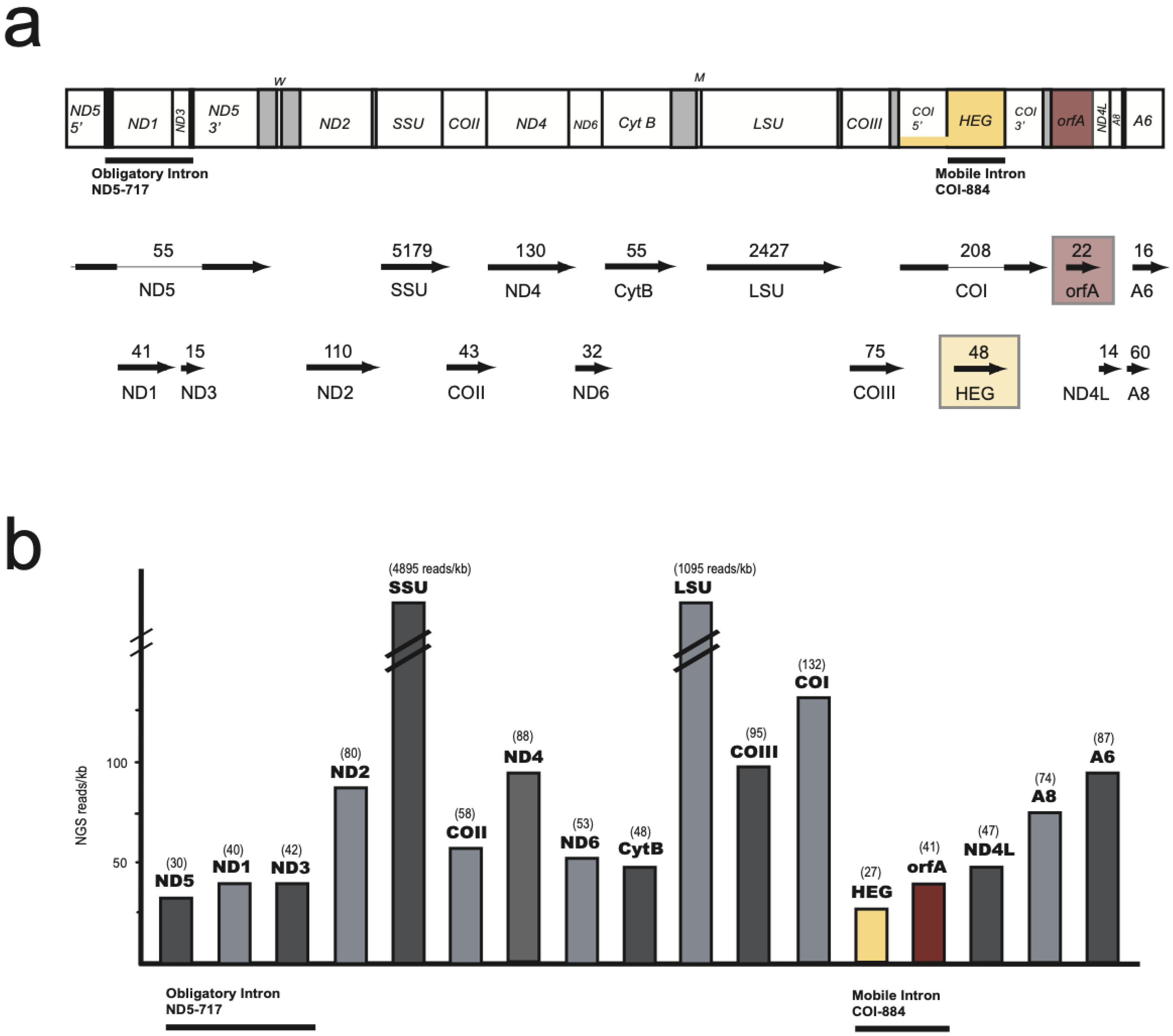
3.1. Characteristic Features of the S. Haddoni Mitogenome
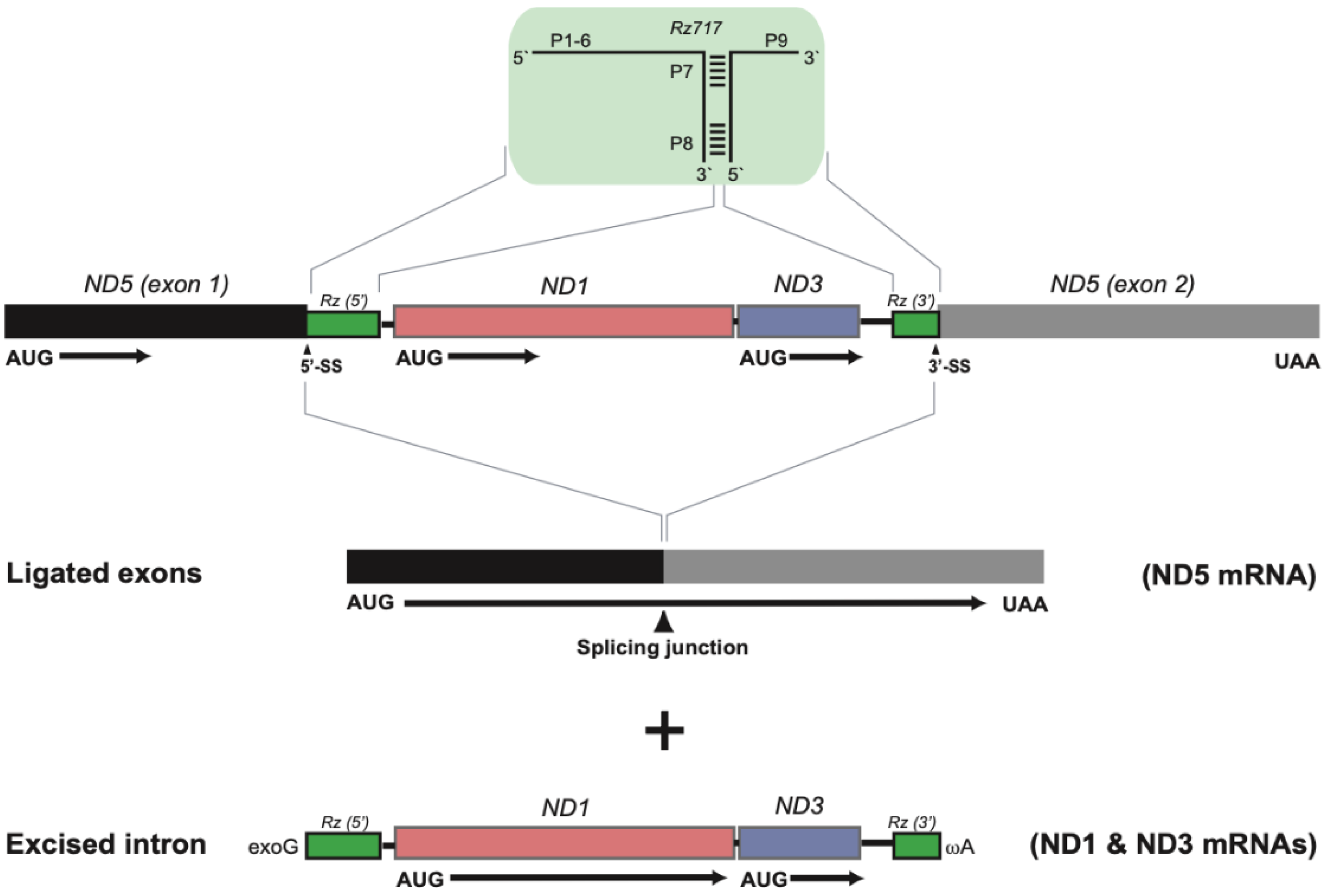
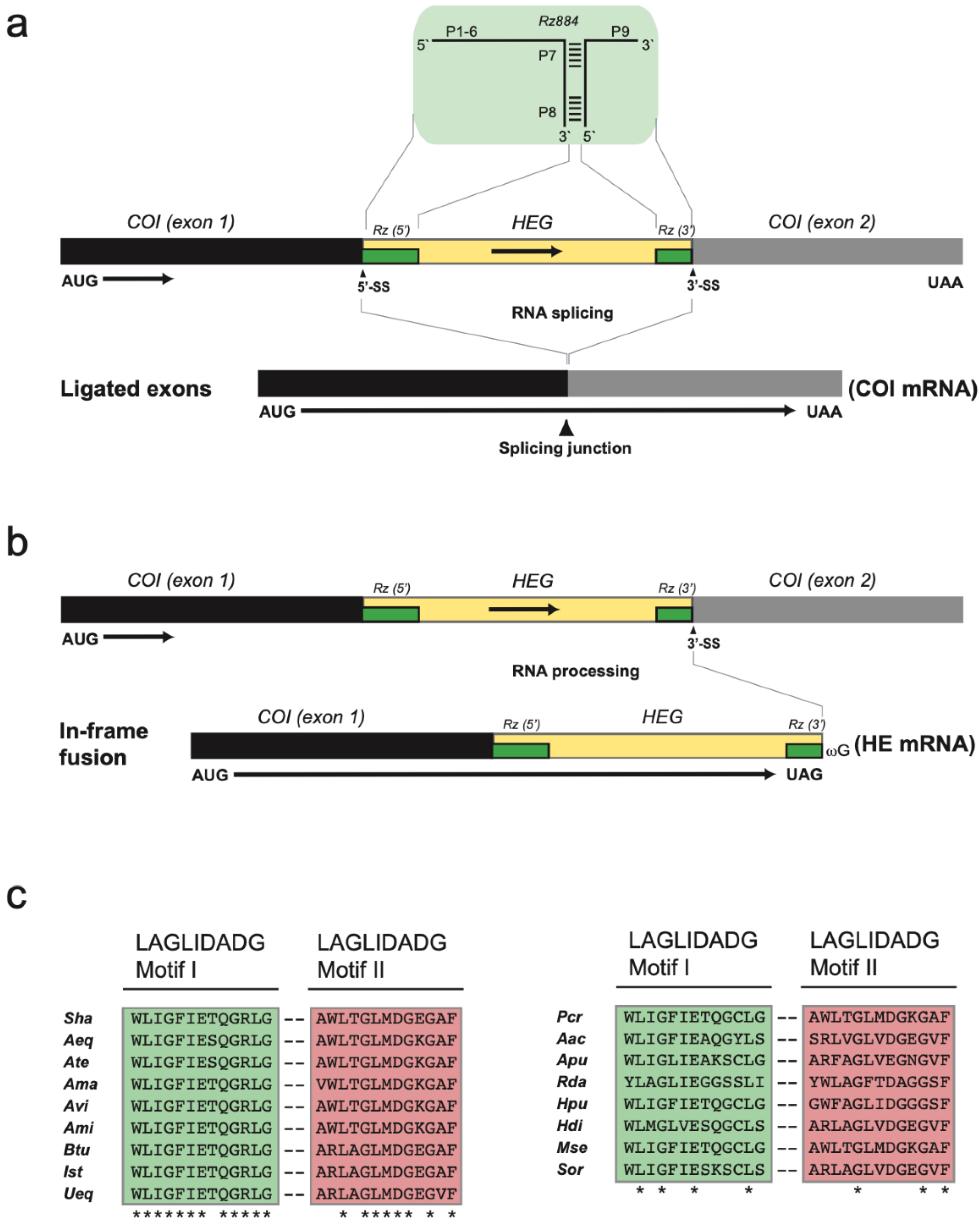
3.2. Two Complex Catalytic Group I Introns
3.3. Nonconventional Protein Coding Genes
3.4. Expression of Mitochondrial Genes
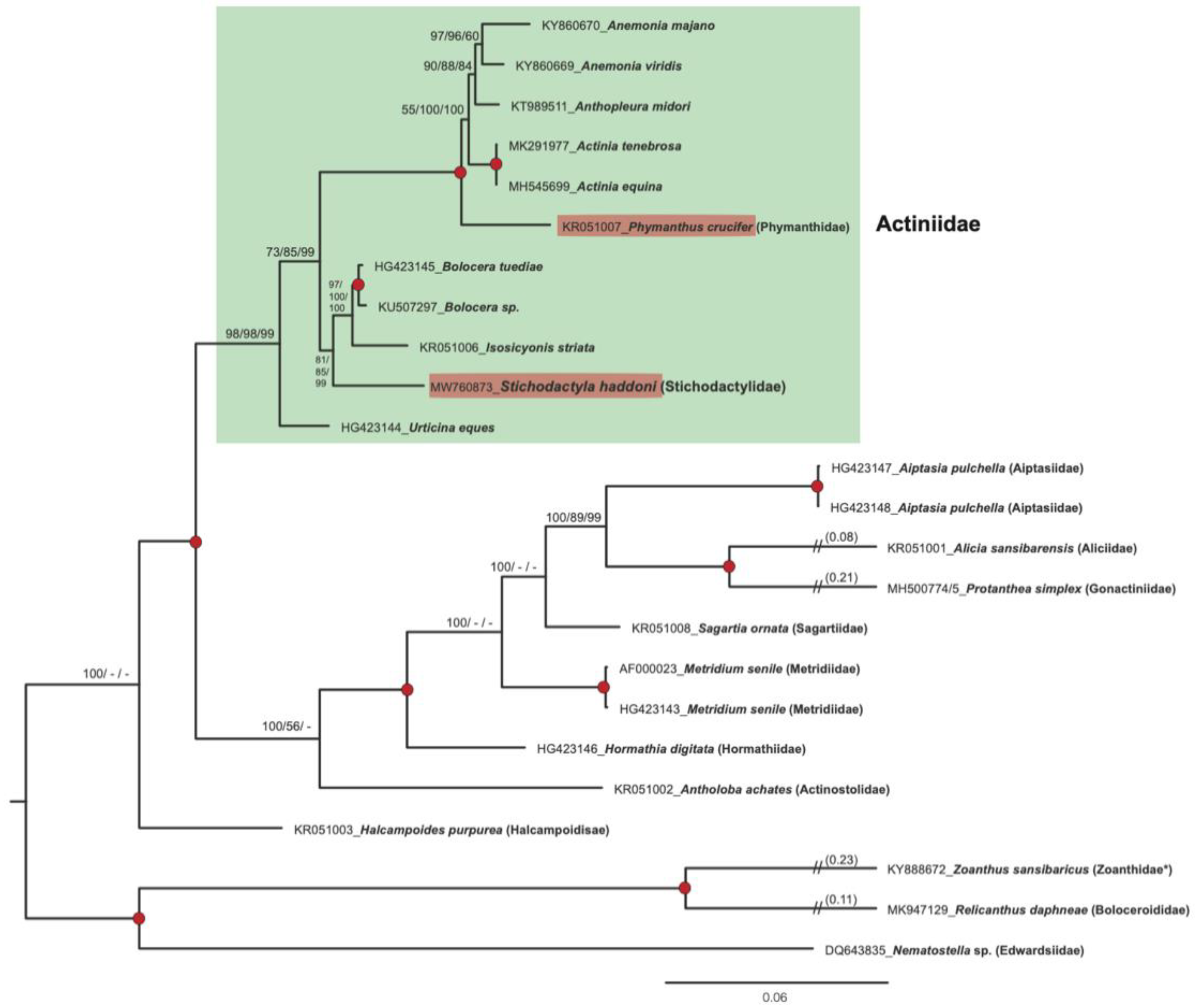
3.5. Mitogenome-Based Phylogeny
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johansen, S.D.; Emblem, Å. Mitochondrial group I introns in hexacorals are regulatory elements. In Advances in the Studies of the Bentic Zone; Soto, L.A., Ed.; IntechOpen: London, UK, 2020; Chapter 7; pp. 1–25. [Google Scholar] [CrossRef] [Green Version]
- Osigus, H.J.; Eitel, M.; Bernt, M.; Donath, A.; Schierwater, B. Mitogenomics at the base of Metazoa. Mol. Phylogenet. Evol. 2013, 69, 339–351. [Google Scholar] [CrossRef]
- Flot, J.F.; Tillier, S. The mitochondrial genome of Pocillopora (Cnidaria: Scleractinia) contains two variable regions: The putative D-loop and a novel ORF of unknown function. Gene 2007, 401, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Beagley, C.T.; Wolstenholme, D.R. Characterization and localization of mitochondrial DNA-encoded tRNA and nuclear DNA-encoded tRNAs in the sea anemone Metridium senile. Curr. Genet. 2013, 59, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Emblem, Å.; Okkenhaug, S.; Weiss, E.S.; Denver, D.R.; Karlsen, B.O.; Moum, T.; Johansen, S.D. Sea anemones possess dynamic mitogenome structures. Mol. Phylogenet. Evol. 2014, 75, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.I.; Johansen, S.D. Zoantharian mitochondrial genomes contain unique complex group I introns and highly conserved intergenic regions. Gene 2017, 628, 24–31. [Google Scholar] [CrossRef]
- Chi, S.I.; Dahl, M.; Emblem, Å.; Johansen, S.D. Giant group I intron in a mitochondrial genome is removed by RNA back-splicing. BMC Mol. Biol. 2019, 20, 16. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.I.; Urbarova, I.; Johansen, S.D. Expression of homing endonuclease gene and insertion-like element in sea anemone mitochondrial genomes: Lesson learned from Anemonia viridis. Gene 2018, 652, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Beagley, C.T.; Okada, N.A.; Wolstenholme, D.R. Two mitochondrial group I introns in a metazoan, the sea anemone Metridium senile: One intron contains genes for subunits 1 and 3 of NADH dehydrogenase. Proc. Natl. Acad. Sci. USA 1996, 93, 5619–5623. [Google Scholar] [CrossRef] [Green Version]
- Haugen, P.; Simon, D.M.; Bhattacharya, D. The natural history of group I introns. Trends Genet. 2005, 21, 111–119. [Google Scholar] [CrossRef]
- Nielsen, H.; Johansen, S.D. Group I introns: Moving in new directions. RNA Biol. 2009, 6, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, A.; Lopez, J.V.; Becking, L.E.; Kelly, M.; Pomponi, S.A.; Worheide, G.; Erpenbeck, D.; Cardenas, P. Evolution of group I introns in Porifera: New evidence for intron mobility and implications for DNA barcoding. BMC Evol. Biol. 2017, 17, 82. [Google Scholar] [CrossRef]
- Cech, T.R.; Damberger, S.H.; Gutell, R.R. Representation of the secondary and tertiary structure of group I introns. Nat. Struct. Biol. 1994, 1, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Vicens, Q.; Cech, T.R. Atomic level architecture of group I introns revealed. Trends Biochem. Sci. 2006, 31, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Hedberg, A.; Johansen, S.D. Nuclear group I introns in self-splicing and beyond. Mob. DNA 2013, 4, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cech, T.R. Self-splicing of group I introns. Annu. Rev. Biochem. 1990, 59, 543–568. [Google Scholar] [CrossRef]
- Stoddard, B.L. Homing endonuclease structure and function. Q. Rev. Biophys. 2005, 38, 49–95. [Google Scholar] [CrossRef]
- Hafez, M.; Hausner, G. Homing endonucleases: DNA scissors on a mission. Genomics 2012, 55, 553–569. [Google Scholar] [CrossRef] [Green Version]
- Emblem, Å.; Karlsen, B.O.; Evertsen, J.; Johansen, S.D. Mitogenome rearrangement in the cold-water scleractinian coral Lophelia pertusa (Cnidaria, Anthozoa) involves a long-term evolving group I intron. Mol. Phylogenet. Evol. 2011, 61, 495–503. [Google Scholar] [CrossRef]
- Goddard, M.R.; Leigh, J.; Roger, A.J.; Pemberton, A.J. Invasion and persistence of a selfish gene in the Cnidaria. PLoS ONE 2006, 1, e3. [Google Scholar] [CrossRef] [Green Version]
- Celis, J.S.; Edgell, D.R.; Stelbrink, B.; Wibberg, D.; Hauffe, T.; Blom, J.; Kalinowski, J.; Wilke, T. Evolutionary and biogeographical implications of degraded LAGLIDADG endonuclease functionality and group I intron occurrence in stony corals (Scleractinia) and mushroom corals (Corallimorpharia). PLoS ONE 2017, 12, e0173734. [Google Scholar] [CrossRef] [Green Version]
- Daly, M.; Brugler, M.R.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; France, S.C.; McFadden, C.S.; Opresko, D.M.; Rodriguez, E.; et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnnaeus. Zootaxa 2007, 1668, 127–182. [Google Scholar] [CrossRef] [Green Version]
- Attaran-Fariman, G.; Javid, P.; Shakouri, A. Morphology and phylogeny of the sea anemone Stichodactyla haddoni (Cnidaria: Anthozoa: Actiniaria) from Chabahar Bay, Iran. Turk. J. Zool. 2015, 39, 998–1003. [Google Scholar] [CrossRef]
- Daly, M.; Chaudhuri, A.; Gusmao, L.; Rodriguez, E. Phylogenetic relationships among sea anemones (Cnidaria: Anthozoa: Actiniaria). Mol. Phylogenet. Evol. 2008, 48, 292–301. [Google Scholar] [CrossRef]
- Chevreux, B.; Wetter, T.; Suhai, S. Genome sequence assembly using trace signals and additional sequence information. German Conf. Bioinform. 1999, 99, 45–56. [Google Scholar]
- Hahn, C.; Bachmann, L.; Chevreux, B. Reconstructing mitochondrial genomes directly from genomic next-generation sequencing reads—A baiting and iterative mapping approach. Nucleic Acids Res. 2013, 41, e129. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Rivera-de-Torre, E.; Martinez-del-Pozo, A.; Garb, J.E. Stichodactyla helianthus’ de novo transcriptome assembly: Discovery of a new actinoporin isoform. Toxicon 2018, 150, 105–114. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Dubin, A.; Chi, S.I.; Emblem, Å.; Moum, T.; Johansen, S.D. Deep-water sea anemone with a two-chromosome mitochondrial genome. Gene 2019, 692, 195–200. [Google Scholar] [CrossRef]
- Nielsen, H.; Fiskaa, T.; Birgisdottir, A.B.; Haugen, P.; Einvik, C.; Johansen, S.D. The ability to form full-length intron RNA circles is a general property of nuclear group I introns. RNA 2003, 9, 1464–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambowitz, A.M.; Belfort, M. Introns as mobile genetic elements. Annu. Rev. Biochem. 1993, 62, 587–622. [Google Scholar] [CrossRef]
- Guo, W.W.; Moran, J.V.; Hoffman, P.W.; Henke, R.M.; Butow, R.A.; Perlman, P.S. The mobile group I intron 3a of the yeast mitochondrial COCI gene codes a 35-kDa processed protein that is an endonuclease but not a maturase. J. Biol. Chem. 1995, 270, 15563–15570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.F.; Kitahara, M.V.; Tachikawa, H.; Fukami, H.; Miller, D.J.; Chen, C.A. Novel organization of the mitochondrial genome in the deep-sea coral, Madrepora oculate (Hexacorallia, Scleractinia, Oculinidae) and its taxonomic implications. Mol. Phylogenet. Evol. 2012, 65, 323–328. [Google Scholar] [CrossRef]
- Lin, M.F.; Kitahara, M.V.; Luo, H.; Tracey, D.; Geller, J.; Fukami, H.; Miller, D.J.; Chen, C.A. Mitochondrial genome rearrangements in the Scleractinia/ corallimorpharian complex: Implications for coral phylogeny. Genome Biol. Evol. 2014, 6, 1086–1095. [Google Scholar] [CrossRef]
- Daly, M.; Gusmao, L.C.; Reft, A.J.; Rodriguez, E. Phylogenetic signal in mitochondrial and nuclear markers in sea anemones (Cnidaria, Actiniaria). Integr. Comp. Biol. 2010, 50, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, E.; Barbeitos, M.S.; Brugler, M.R.; Crowley, L.M.; Grajales, A.; Gusmao, L.; Haussermann, V.; Reft, A.; Daly, M. Hidden among sea anemones: The first comprehensive phylogenetic reconstruction of the order Actiniaria (Cnidaria, Anthozoa, Hexacorallia) reveals a novel group of hexacorals. PLoS ONE 2014, 9, e96998. [Google Scholar] [CrossRef]
- Titus, B.M.; Benedict, C.; Laroche, R.; Gusmao, L.C.; Van Deusen, V.; Chiodo, T.; Meyer, C.P.; Berumen, M.L.; Bartholomew, A.; Yanagi, K.; et al. Phylogenetic relationships among the clownfish-hosting sea anemones. Mol. Phylogenet. Evol. 2019, 139, 106526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, Q. Complete mitochondrial genome of the sea anemone, Anthopleura midori (Actiniaria: Actiniidae). Mitochondrial DNA 2017, 28, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Foox, J.; Brugler, M.; Siddall, M.E.; Rodriguez, E. Multiplexed pyrosequencing of nine sea anemone (Cnidaria: Anthozoa: Hexacorallia: Actiniaria) mitochondrial genomes. Mitochondrial DNA 2016, 27, 2826–2832. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johansen, S.D.; Chi, S.I.; Dubin, A.; Jørgensen, T.E. The Mitochondrial Genome of the Sea Anemone Stichodactyla haddoni Reveals Catalytic Introns, Insertion-Like Element, and Unexpected Phylogeny. Life 2021, 11, 402. https://doi.org/10.3390/life11050402
Johansen SD, Chi SI, Dubin A, Jørgensen TE. The Mitochondrial Genome of the Sea Anemone Stichodactyla haddoni Reveals Catalytic Introns, Insertion-Like Element, and Unexpected Phylogeny. Life. 2021; 11(5):402. https://doi.org/10.3390/life11050402
Chicago/Turabian StyleJohansen, Steinar Daae, Sylvia I. Chi, Arseny Dubin, and Tor Erik Jørgensen. 2021. "The Mitochondrial Genome of the Sea Anemone Stichodactyla haddoni Reveals Catalytic Introns, Insertion-Like Element, and Unexpected Phylogeny" Life 11, no. 5: 402. https://doi.org/10.3390/life11050402
APA StyleJohansen, S. D., Chi, S. I., Dubin, A., & Jørgensen, T. E. (2021). The Mitochondrial Genome of the Sea Anemone Stichodactyla haddoni Reveals Catalytic Introns, Insertion-Like Element, and Unexpected Phylogeny. Life, 11(5), 402. https://doi.org/10.3390/life11050402