
A Causal Analysis of the Effect of Age and Sex Differences on Brain Atrophy in the Elderly Brain


Abstract
:1. Introduction
2. Methods
2.1. Study Participants
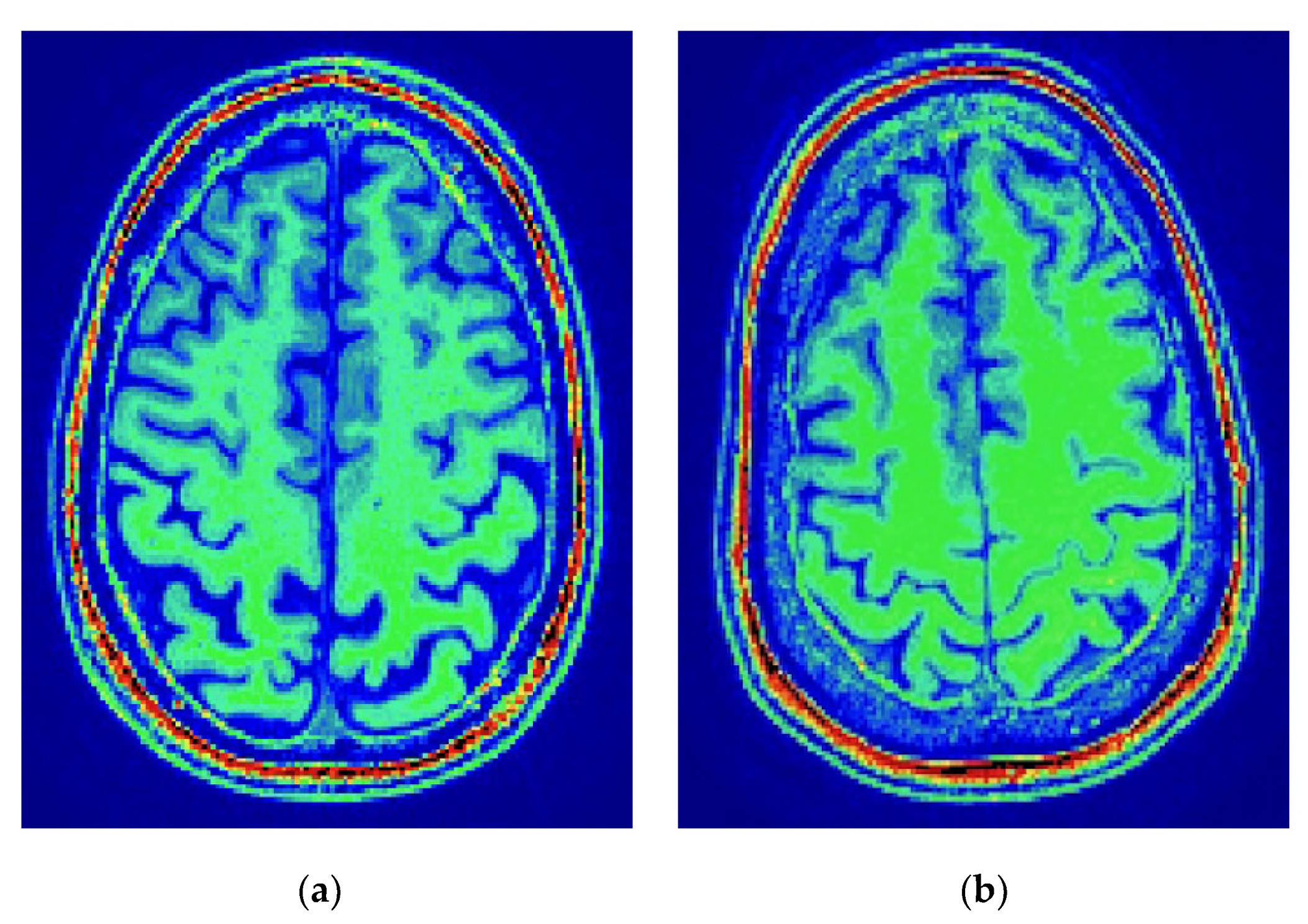
2.2. MRI Data Acquisition and Preprocessing
2.3. Statistical Data Analysis
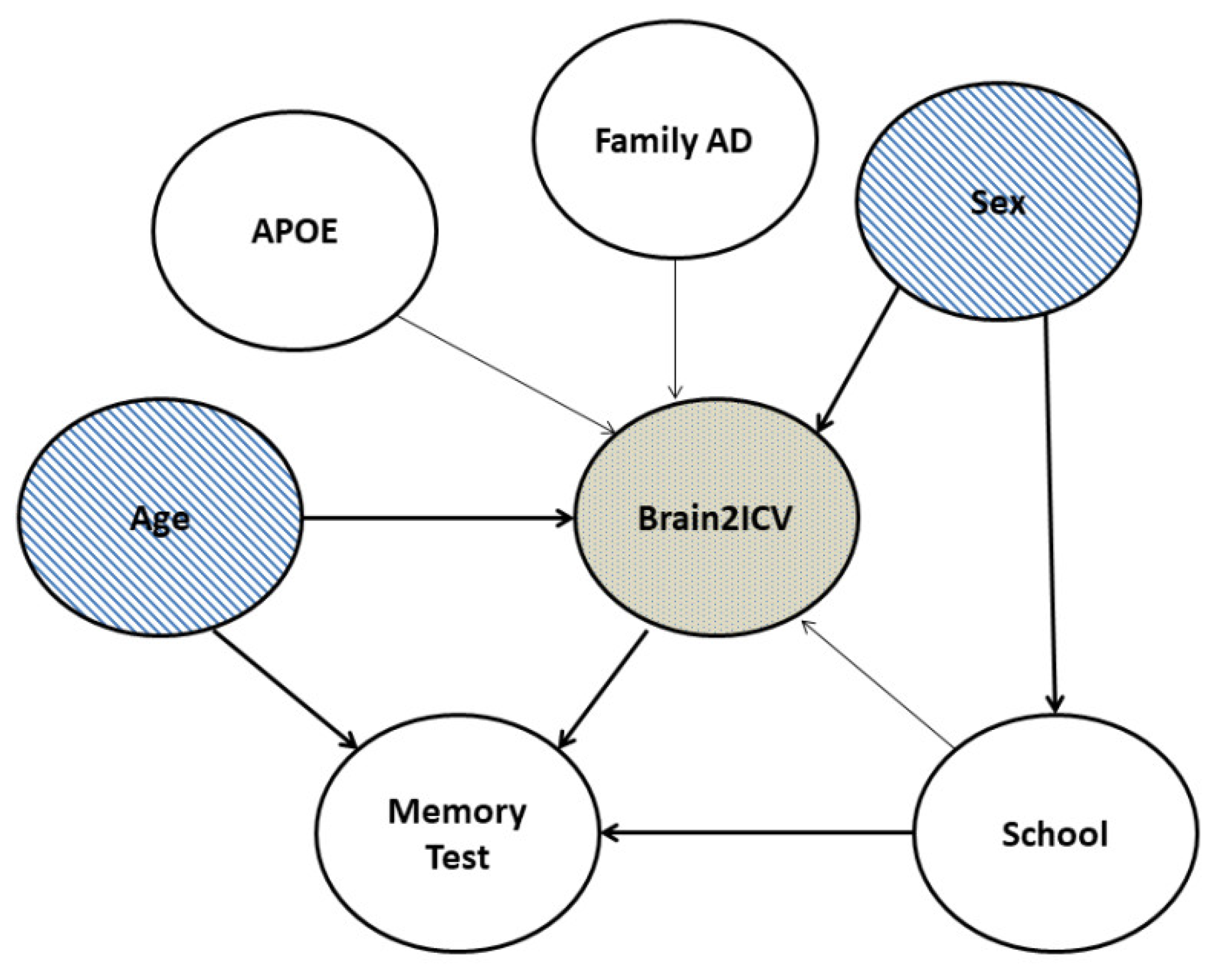
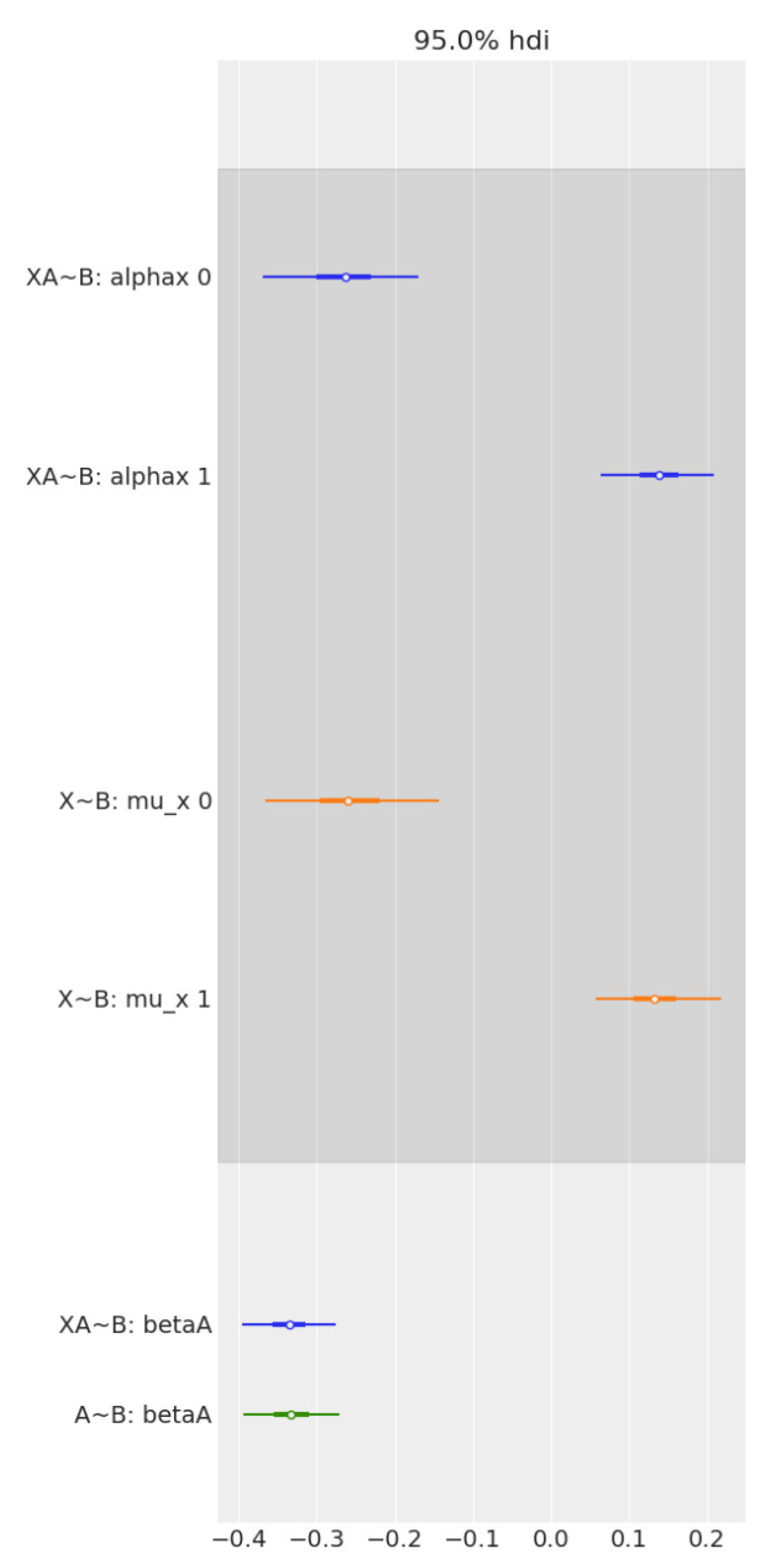
2.4. Causal Data Analysis
3. Results
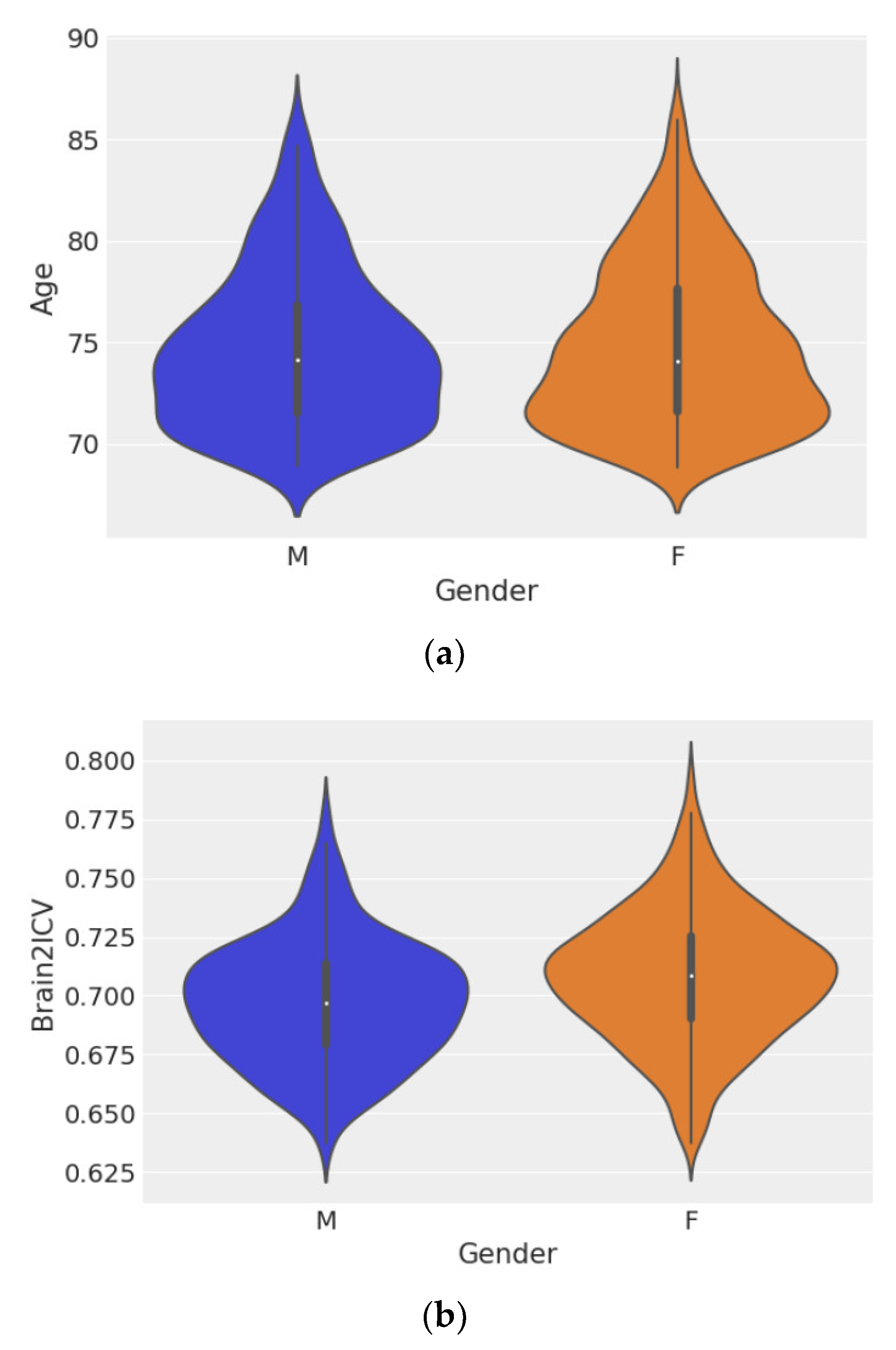
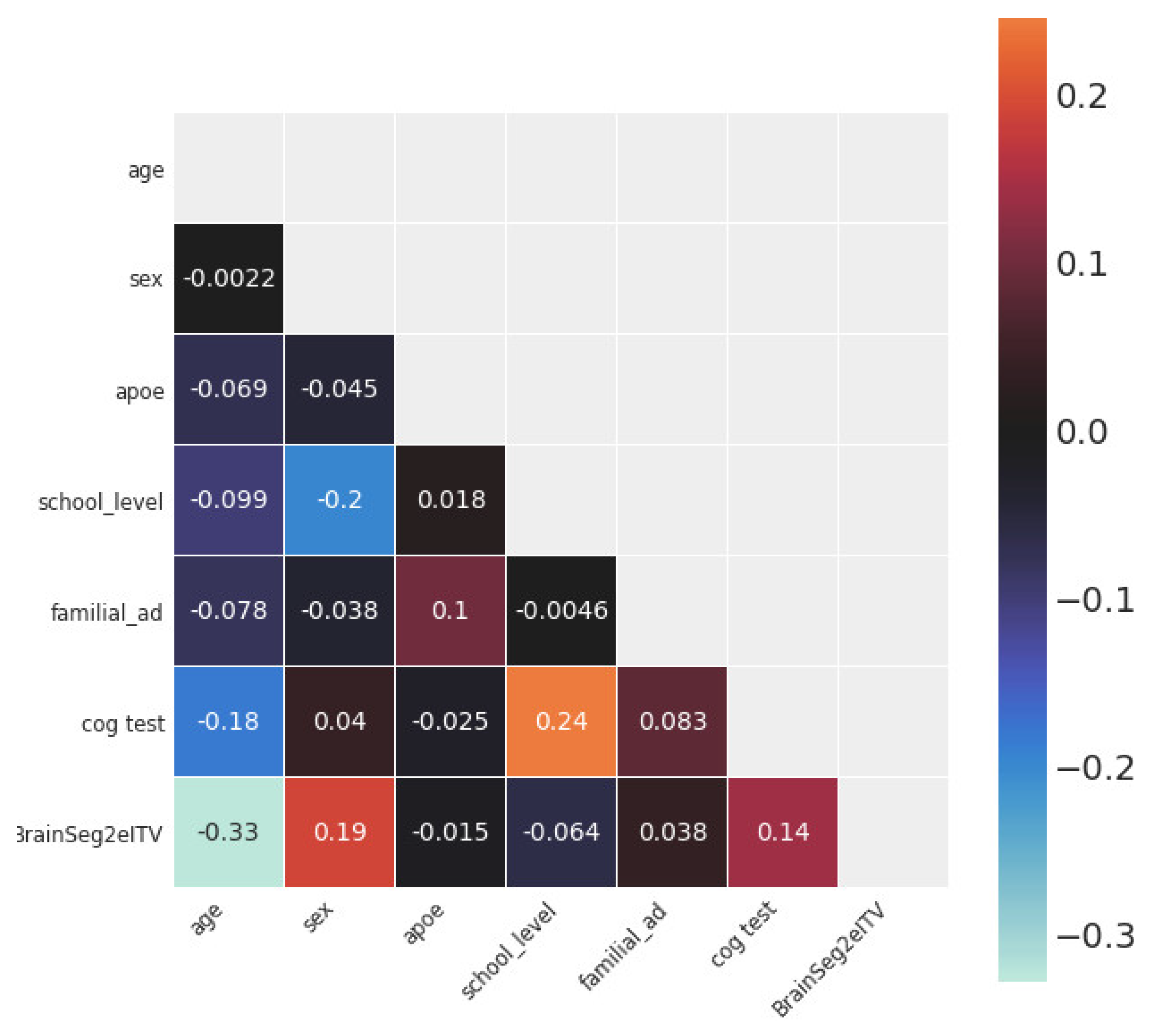
3.1. Statistical and Correlation Analysis
3.2. Causal Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyd, R. Tables of the weights of the human body and internal organs in the sane and insane of both sexes at various ages, arranged from 2614 post-mortem examinations. Phil. Trans. R. Soc. Lond. 1861, 151. [Google Scholar] [CrossRef]
- Marshall, J. Relations between the weight of the brain and its parts, and the stature and mass of the body, in man. J. Anat. Physiol. 1892, 26 Pt 4, 445. [Google Scholar] [PubMed]
- Esiri, M.M. Ageing and the brain. J. Pathol. 2007, 211, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Corsellis, J. Evidence for a secular increase in human brain weight during the past century. Ann. Hum. Biol. 1977, 4, 253–257. [Google Scholar] [CrossRef]
- Ho, K.-C.; Roessmann, U.; Straumfjord, J.; Monroe, G. Analysis of brain weight. i. adult brain weight in relation to sex, race, and age. Arch. Pathol. Lab. Med. 1980, 104, 635–639. [Google Scholar]
- Dekaban, A.S.; Sadowsky, D. Changes in brain weights during the span of human life: Relation of brain weights to body heights and body weights. Ann. Neurol. 1978, 4, 345–356. [Google Scholar] [CrossRef]
- Fox, N.C.; Schott, J.M. Imaging cerebral atrophy: Normal ageing to alzheimer’s disease. Lancet 2004, 363, 392–394. [Google Scholar] [CrossRef]
- Despotović, I.; Goossens, B.; Philips, W. Mri segmentation of the human brain: Challenges, methods, and applications. Comput. Math. Methods Med. 2015, 2015, 450341. [Google Scholar]
- Wenger, E.; Mårtensson, J.; Noack, H.; Bodammer, N.C.; Kühn, S.; Schaefer, S.; Heinze, H.-J.; Düzel, E.; Bäckman, L.; Lindenberger, U.; et al. Comparing manual and automatic segmentation of hippocampal volumes: Reliability and validity issues in younger and older brains. Hum. Brain Mapp. 2014, 35, 4236–4248. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, J.; Csernansk, J.G.; Davatzikos, C.; Fox, N.C.; Frisoni, G.B.; Thompson, P.M. Computer-assisted imaging to assess brain structure in healthy and diseased brains. Lancet Neurol. 2003, 2, 79–88. [Google Scholar] [CrossRef]
- Caspi, Y.; Brouwer, R.M.; Schnack, H.G.; vande Nieuwenhuijzen, M.E.; Cahn, W.; Kahn, R.S.; Niessen, W.J.; vander Lugt, A.; Pol, H.H. Changes in the intracranial volume from early adulthood to the sixth decade of life: A longitudinal study. NeuroImage 2020, 220, 116842. [Google Scholar] [CrossRef]
- Raz, N.; Rodrigue, K.M. Differential aging of the brain: Patterns, cognitive correlates and modifiers. Neurosci. Biobehav. Rev. 2006, 30, 730–748. [Google Scholar] [CrossRef]
- Holland, D.; Chang, L.; Ernst, T.M.; Curran, M.; Buchthal, S.D.; Alicata, D.; Skranes, J.; Johansen, H.; Hernandez, A.; Yamakawa, R.; et al. Structural growth trajectories and rates of change in the first 3 months of infant brain development. JAMA Neurol. 2014, 71, 1266–1274. [Google Scholar] [CrossRef] [Green Version]
- Hedman, A.M.; van Haren, N.E.; Schnack, H.G.; Kahn, R.S.; HulshoffPol, H.E. Human brain changes across the life span: A review of 56 longitudinal magnetic resonance imaging studies. Hum. Brain Mapp. 2012, 33, 1987–2002. [Google Scholar] [CrossRef]
- Fjell, A.M.; Walhovd, K.B. Structural brain changes in aging: Courses, causes and cognitive consequences. Rev. Neurosci. 2010, 21, 187–222. [Google Scholar] [CrossRef]
- Rast, P.; Kennedy, K.M.; Rodrigue, K.M.; Robinson, P.R.; Gross, A.L.; McLaren, D.G.; Grabowski, T.; Schaie, K.W.; Willis, S.L. Apoeε4 genotype and hypertension modify 8-year cortical thinning: Five occasion evidence from the seattle longitudinal study. Cereb. Cortex 2017, 28, 1934–1945. [Google Scholar] [CrossRef] [Green Version]
- Chételat, G.; Landeau, B.; Eustache, F.; Mézenge, F.; Viader, F.; de La Sayette, V.; Desgranges, B.; Baron, J.-C. Using voxel-based morphometry to map the structural changes associated with rapid conversion in mci: A longitudinal mri study. Neuroimage 2005, 27, 934–946. [Google Scholar] [CrossRef]
- Misra, C.; Fan, Y.; Davatzikos, C. Baseline and longitudinal patterns of brain atrophy in mci patients, and their use in prediction of short-term conversion to ad: Results from adni. Neuroimage 2009, 44, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Rusinek, H.; De Santi, S.; Frid, D.; Tsui, W.-H.; Tarshish, C.Y.; Convit, A.; de Leon, M.J. Regional brain atrophy rate predicts future cognitive decline: 6-year longitudinal mr imaging study of normal aging. Radiology 2003, 229, 691–696. [Google Scholar] [CrossRef]
- Ghione, E.; Bergsland, N.; Dwyer, M.G.; Hagemeier, J.; Jakimovski, D.; Paunkoski, I.; Ramasamy, D.P.; Carl, E.; Hojnacki, D.; Kolb, C.; et al. Aging and brain atrophy in multiple sclerosis. J. Neuroimaging 2019, 29, 527–535. [Google Scholar] [CrossRef]
- Oschwald, J.; Guye, S.; Liem, F.; Rast, P.; Willis, S.; Röcke, C.; Jäncke, L.; Martin, M.; Mérillat, S. Brain structure and cognitive ability in healthy aging: A review on longitudinal correlated change. Rev. Neurosci. 2019, 31, 1–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egloff, L.; Lenz, C.; Studerus, E.; Harrisberger, F.; Smieskova, R.; Schmidt, A.; Huber, C.; Simon, A.; Lang, U.E.; Riecher-Rössler, A.; et al. Sexually dimorphic subcortical brain volumes in emerging psychosis. Schizophr. Res. 2018, 199, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Király, A.; Szabó, N.; Tóth, E.; Csete, G.; Faragó, P.; Kocsis, K.; Must, A.; Vécsei, L.; Kincses, Z.T. Male brain ages faster: The age and gender dependence of subcortical volumes. Brain Imaging Behav. 2016, 10, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Bigler, E.D.; Tate, D.F. Brain volume, intracranial volume, and dementia. Investig. Radiol. 2001, 36, 539–546. [Google Scholar] [CrossRef]
- Greenberg, D.L.; Messer, D.F.; Payne, M.E.; MacFall, J.R.; Provenzale, J.M.; Steffens, D.C.; Krishnan, R.R. Aging, gender, and the elderly adult brain: An examination of analytical strategies. Neurobiol. Aging 2008, 29, 290–302. [Google Scholar] [CrossRef] [Green Version]
- Pintzka, C.W.S.; Hansen, T.I.; Evensmoen, H.R.; Håberg, A.K. Marked effects of intracranial volume correction methods on sex differences in neuroanatomical structures: A hunt mri study. Front. Neurosci. 2015, 9, 238. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.A.S.; Rodrigues, T.P.; Zatz, M.; Lebrao, M.L.; Duarte, Y.A.; Naslavsky, M.S.; Nascimento, F.B.; Amaro, E. Quantitative evaluation of brain volume among elderly individuals in são paulo, brazil: A population-based study. Radiol. Bras. 2019, 52, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Nordenskjöld, R.; Malmberg, F.; Larsson, E.-M.; Simmons, A.; Ahlström, H.; Johansson, L.; Kullberg, J. Intracranial volume nor- malization methods: Considerations when investigating gender differences in regional brain volume. Psychiatry Res. Neuroimaging 2015, 231, 227–235. [Google Scholar] [CrossRef]
- Gómez-Ramírez, J.; Fernández-Blázquez, M.A.; González-Rosa, J.J. Prediction of chronological age in healthy elderly subjects with machine learning from mri brain segmentation and cortical parcellation. Brain Sci. 2022, 12, 579. [Google Scholar] [CrossRef]
- Ham, S.; Lee, S.-J.V. Advances in transcriptome analysis of human brain aging. Exp. Mol. Med. 2020, 52, 1787–1797. [Google Scholar] [CrossRef]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Sayed, N.; Huang, Y.; Nguyen, K.; Krejciova-Rajaniemi, Z.; Grawe, A.P.; Gao, T.; Tibshirani, R.; Hastie, T.; Alpert, A.; Cui, L.; et al. An inflammatory aging clock (iage) based on deep learning tracks multimorbidity, immunosenescence, frailty and cardiovascular aging. Nat. Aging 2021, 1, 598–615. [Google Scholar] [CrossRef]
- Pearl, J. Causal diagrams for empirical research. Biometrika 1995, 82, 669–688. [Google Scholar] [CrossRef]
- Fernández-Blázquez, M.A.; Noriega-Ruiz, B.; Ávila Villanueva, M.; Valentí-Soler, M.; Frades-Payo, B.; Ser, T.D.; Gómez-Ramírez, J. Impact of individual and neighborhood dimensions of socioeconomic status on the prevalence of mild cognitive impairment over seven-year follow-up. Aging Ment. Health 2021, 25, 814–823. [Google Scholar] [CrossRef]
- Gómez-Ramírez, J.; Villanueva, M.Á.; Payo, B.F.; del Ser Quijano, T.; Soler, M.V.; Sevilla, M.A.Z.; Fernández-Blázquez, M.Á. Exploratory data analysis in the vallecas project: A six years longitudinal study in healthy brain aging. bioRxiv 2019, 674853. [Google Scholar] [CrossRef]
- Sanz-Blasco, R.; Ruiz-Sánchez de León, J.M.; Ávila-Villanueva, M.; Valentí-Soler, M.; Gómez-Ramírez, J.; Fernández-Blázquez, M.A. Transition from mild cognitive impairment to normal cognition: Determining the predictors of reversion with multi-state markov models. Alzheimer’s Dement. 2022, 18, 1177–1185. [Google Scholar] [CrossRef]
- Estimated Total Intracranial Volume aka ICV. Freesurfer. 2020. Available online: http://www.freesurfer.net/fswiki/eTIV (accessed on 6 April 2021).
- Buckner, R.L.; Head, D.; Parker, J.; Fotenos, A.F.; Marcus, D.; Morris, J.C.; Snyder, A.Z. A unified approach for morphometric and functional data analysis in young, old, and demented adults using automated atlas-based head size normalization: Reliability and validation against manual measurement of total intracranial volume. Neuroimage 2004, 23, 724–738. [Google Scholar] [CrossRef]
- Fischl, B. Freesurfer. Neuroimage 2012, 62, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Evans, A.C.; Collins, D.L.; Mills, S.; Brown, E.D.; Kelly, R.L.; Peters, T.M. 3D statistical neuroanatomical models from 305 mri volumes. In Proceedings of the 1993 IEEE Conference Record Nuclear Science Symposium and Medical Imaging Conference, San Francisco, CA, USA, 31 October 1993–6 November 1993; pp. 1813–1817. [Google Scholar]
- Klasson, N.; Olsson, E.; Eckerström, C.; Malmgren, H.; Wallin, A. Estimated intracranial volume from freesurfer is biased by total brain volume. Eur. Radiol. Exp. 2018, 2, 24. [Google Scholar] [CrossRef]
- Heinen, R.; Bouvy, W.H.; Mendrik, A.M.; Viergever, M.A.; Biessels, G.J.; DeBresser, J. Robustness of automated methods for brain volume measurements across different mri field strengths. PLoS ONE 2016, 11, e0165719. [Google Scholar] [CrossRef] [Green Version]
- Whitwell, J.L.; Crum, W.R.; Watt, H.C.; Fox, N.C. Normalization of cerebral volumes by use of intracranial volume: Implications for longitudinal quantitative mr imaging. Am. J. Neuroradiol. 2001, 22, 1483–1489. [Google Scholar] [PubMed]
- Davidson-Pilon, C. Bayesian Methods for Hackers: Probabilistic Programming and Bayesian Inference; Addison-Wesley Professional: Boston, MA, USA, 2015. [Google Scholar]
- Pearl, J.; Mackenzie, D. The Book of Why: The New Science of Cause and Effect; Basic Books: New York, NY, USA, 2018. [Google Scholar]
- Pearl, J. Causality: Models, Reasoning and Inference; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Peters, J.; Janzing, D.; Schölkopf, B. Elements of Causal Inference: Foundations and Learning Algorithms; The MIT Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Hernán, M.A.; Hernández-Díaz, S.; Werler, M.M.; Mitchell, A.A. Causal knowledge as a prerequisite for confounding evaluation: An application to birth defects epidemiology. Am. J. Epidemiol. 2002, 155, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Piccininni, M.; Konigorski, S.; Rohmann, J.L.; Kurth, T. Directed acyclic graphs and causal thinking in clinical risk prediction modeling. Bmc Med. Res. Methodol. 2020, 20, 179. [Google Scholar] [CrossRef] [PubMed]
- Erten-Lyons, D.; Dodge, H.H.; Woltjer, R.; Silbert, L.C.; Howieson, D.B.; Kramer, P.; Kaye, J.A. Neuropathologic basis of age-associated brain atrophy. JAMA Neurol. 2013, 70, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Fjell, A.M.; Walhovd, K.B.; Fennema-Notestine, C.; McEvoy, L.K.; Hagler, D.J.; Holland, D.; Brewer, J.B.; Dale, A.M. One-year brain atrophy evident in healthy aging. J. Neurosci. 2009, 29, 15223–15231. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Ramirez, J.; Quilis-Sancho, J.; Fernandez-Blazquez, M.A. A comparative analysis of mri automated segmentation of subcortical brain volumes in a large dataset of elderly subjects. Neuroinformatics 2021, 20, 63–70. [Google Scholar] [CrossRef]
- Seabold, S.; Perktold, J. Statsmodels: Econometric and statistical modeling with python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010. [Google Scholar]
- Martin, O. Bayesian Analysis with Python: Introduction to Statistical Modeling and Probabilistic Programming Using PyMC3 and ArviZ; Packt Publishing Ltd.: Birmingham, UK, 2018. [Google Scholar]
- Gamerman, D.; Lopes, H.F. Markov Chain Monte Carlo: Stochastic Simulation for Bayesian Inference; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Patil, A.; Huard, D.; Fonnesbeck, C.J. Pymc: Bayesian stochastic modelling in python. J. Stat. Softw. 2010, 35, 1. [Google Scholar] [CrossRef] [Green Version]
- American Statistical Association. Statement on statistical significance and p-values. Am. Stat. 2016, 70, 129–133. [Google Scholar]
- Benjamin, D.J.; Berger, J.O.; Johannesson, M.; Nosek, B.A.; Wagenmakers, E.-J.; Berk, R.; Bollen, K.A.; Brembs, B.; Brown, L.; Camerer, C.; et al. Redefine statistical significance. Nat. Hum. Behav. 2018, 2, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Wasserstein, R.L.; Schirm, A.L.; Lazar, N.A. Moving to a world beyond p < 0.05. Am. Stat. 2019, 73 (Suppl. 1), 1–19. [Google Scholar]
- Gomez-Ramirez, J.; Sanz, R. On the limitations of standard statistical modeling in biological systems: A full bayesian approach for biology. Progress Biophys. Mol. Biol. 2013, 113, 80–91. [Google Scholar] [CrossRef]
- Jaynes, E.T. Probability Theory: The Logic of Science; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Pearson, K. The grammar of science. Nature 1892, 46, 199–200. [Google Scholar] [CrossRef]
- Royle, N.; Hernández, M.V.; Maniega, S.M.; Arabisala, B.; Bastin, M.; Deary, I.; Wardlaw, J. Influence of thickening of the inner skull table on intracranial volume measurement in older people. Magn. Reson. Imaging 2013, 31, 918–922. [Google Scholar] [CrossRef]
- Aribisala, B.S.; Royle, N.A.; Valdés Hernández, M.C.; Murray, C.; Penke, L.; Gow, A.; Maniega, S.M.; Starr, J.M.; Bastin, M.; Deary, I.; et al. Potential effect of skull thickening on the associations between cognition and brain atrophy in ageing. Age Ageing 2014, 43, 712–716. [Google Scholar] [CrossRef]
- Goyal, M.S.; Blazey, T.M.; Su, Y.; Couture, L.E.; Durbin, T.J.; Bateman, R.J.; Benzinger, T.L.-S.; Morris, J.C.; Raichle, M.E.; Vlassenko, A.G. Persistent metabolic youth in the aging female brain. Proc. Natl. Acad. Sci. USA 2019, 116, 3251–3255. [Google Scholar] [CrossRef] [Green Version]
- Salvatier, J.; Wiecki, T.V.; Fonnesbeck, C. Probabilistic programming in python using pymc3. PeerJ Comput. Sci. 2016, 2, e55. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Mean | SD |
|---|---|---|
| Age | 74.72 | 3.86 |
| Cognitive Score | 9.41 | 2.66 |
| Brain2ICV | 0.70 | 0.03 |
| Total | % | |
| Sex | ||
| Male | 303 | 34.04 |
| Female | 587 | 65.96 |
| APOE | ||
| Noncarriers | 726 | 81.57 |
| Heterozygous e4 | 157 | 17.64 |
| Homozygous e4 | 7 | 0.79 |
| Level of education | ||
| No formal education | 170 | 19.10 |
| Primary education | 265 | 29.76 |
| High school | 224 | 25.17 |
| University | 231 | 25.96 |
| Family history of AD | ||
| No | 670 | 75.28 |
| Yes | 220 | 24.72 |
| F | PR(>F) | |
|---|---|---|
| Age | 119.694 | ** 3.242 × 10−26 |
| Sex | 32.746 | ** 1.438 × 10−8 |
| APOE | 1.099 | 2.948 × 10−1 |
| Family history of AD | 1.022 | 3.124 × 10−1 |
| Level of education | 3.530 | 6.058 × 10−2 |
| Mean | sd | HDI 3% | HDI 97% | |
|---|---|---|---|---|
| µ1 | 0.697 | 0.002 | 0.694 | 0.70 |
| µ2 | 0.708 | 0.001 | 0.706 | 0.71 |
| σ | 0.027 | 0.001 | 0.026 | 0.028 |
| µ1 − µ2 | −0.011 | 0.002 | −0.014 | −0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Ramírez, J.; Fernández-Blázquez, M.A.; González-Rosa, J.J. A Causal Analysis of the Effect of Age and Sex Differences on Brain Atrophy in the Elderly Brain. Life 2022, 12, 1586. https://doi.org/10.3390/life12101586
Gómez-Ramírez J, Fernández-Blázquez MA, González-Rosa JJ. A Causal Analysis of the Effect of Age and Sex Differences on Brain Atrophy in the Elderly Brain. Life. 2022; 12(10):1586. https://doi.org/10.3390/life12101586
Chicago/Turabian StyleGómez-Ramírez, Jaime, Miguel A. Fernández-Blázquez, and Javier J. González-Rosa. 2022. "A Causal Analysis of the Effect of Age and Sex Differences on Brain Atrophy in the Elderly Brain" Life 12, no. 10: 1586. https://doi.org/10.3390/life12101586
APA StyleGómez-Ramírez, J., Fernández-Blázquez, M. A., & González-Rosa, J. J. (2022). A Causal Analysis of the Effect of Age and Sex Differences on Brain Atrophy in the Elderly Brain. Life, 12(10), 1586. https://doi.org/10.3390/life12101586