
Discovery of Small Molecules from Echinacea angustifolia Targeting RNA-Dependent RNA Polymerase of Japanese Encephalitis Virus

 , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Methodology
2.1. Receptor and Ligand Collection
2.2. Virtual Screening, Redocking Analysis, and ADME
2.3. Molecular Dynamics Simulation
2.4. Post-Simulation Analysis
2.4.1. Essential Dynamics
2.4.2. Prime MM/GBSA Binding Free Energy Calculations
3. Results and Discussion
3.1. Structure-Based Virtual Screening and ADME
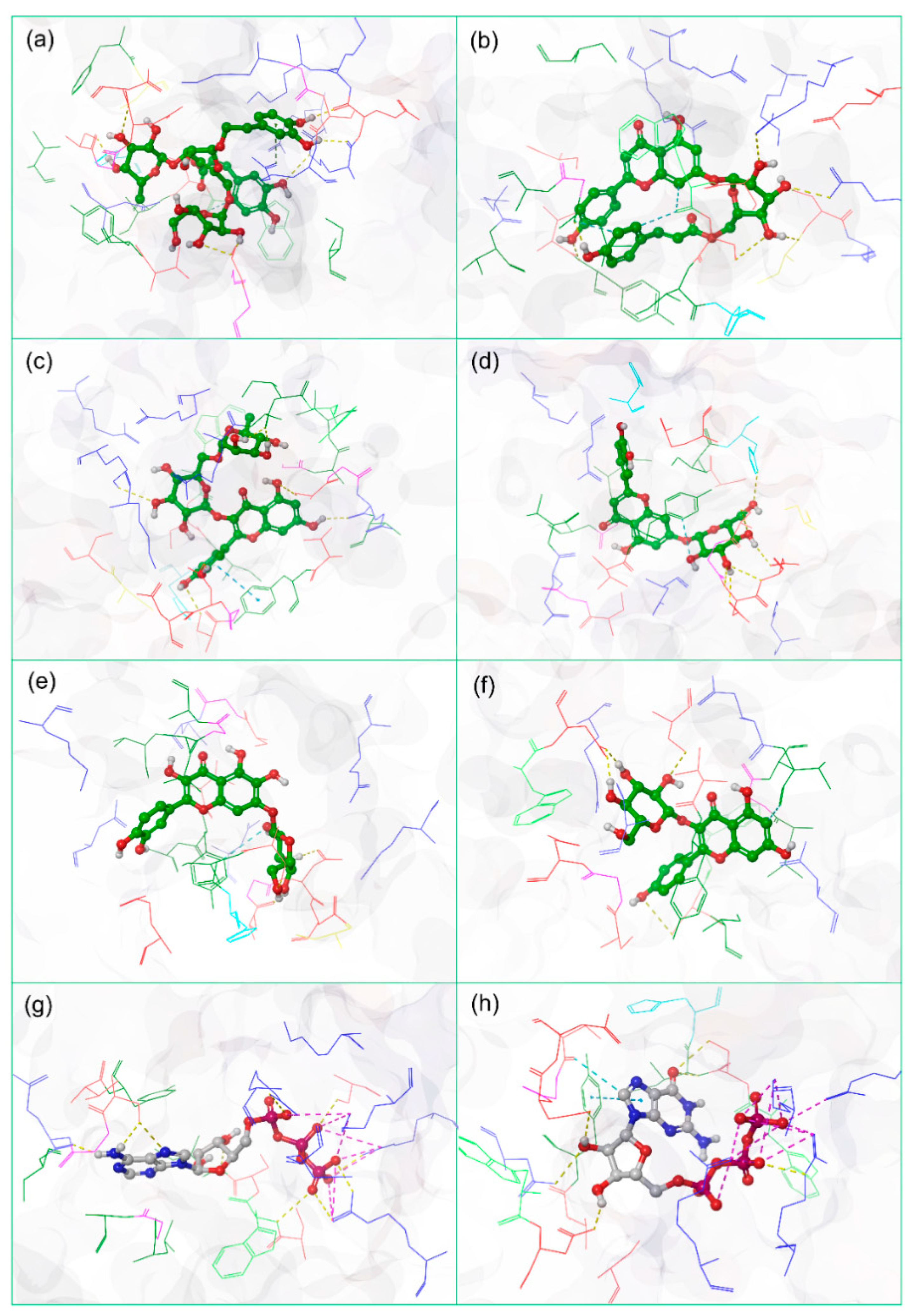
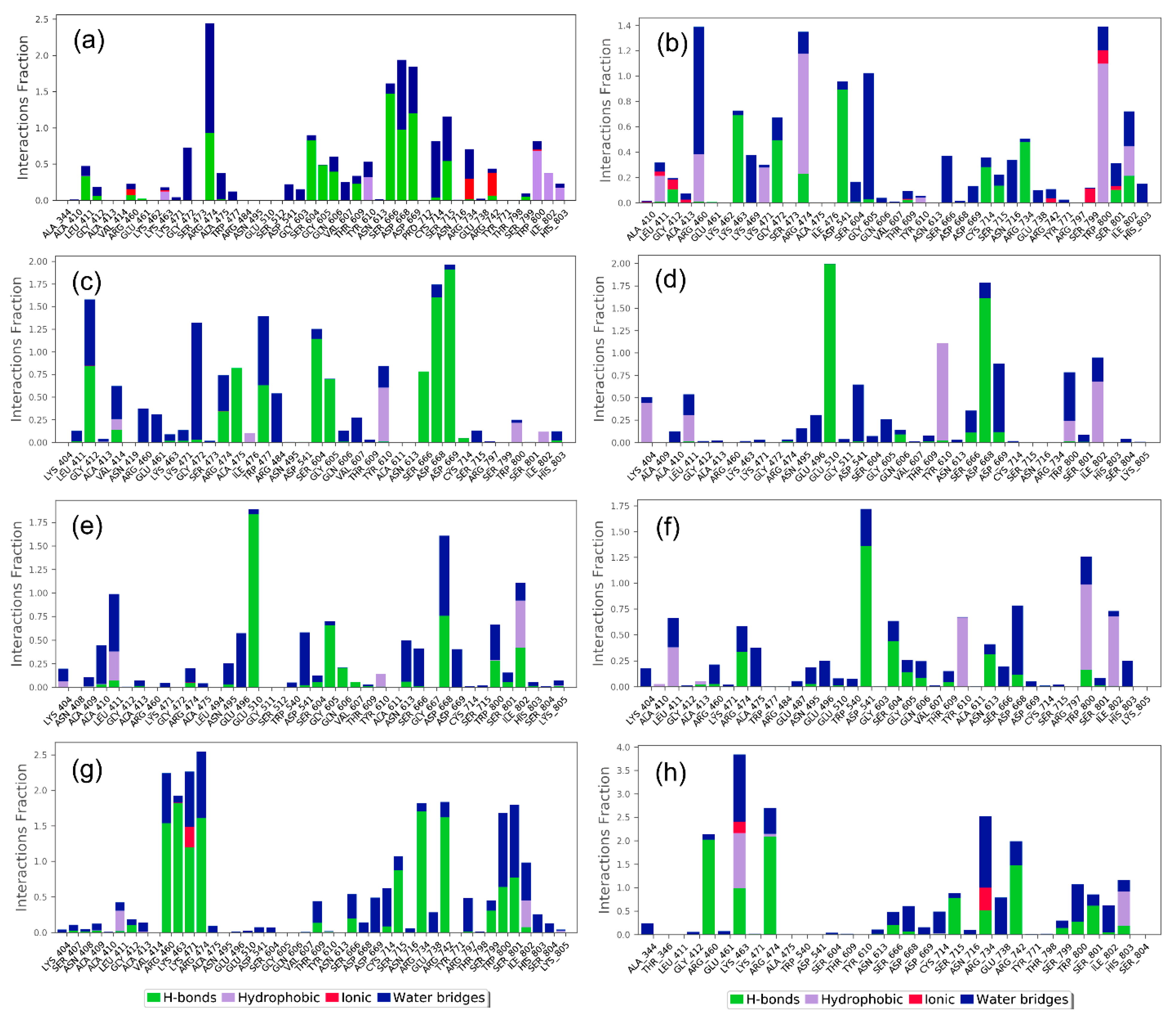
3.2. Redocking and Molecular Contact Analysis
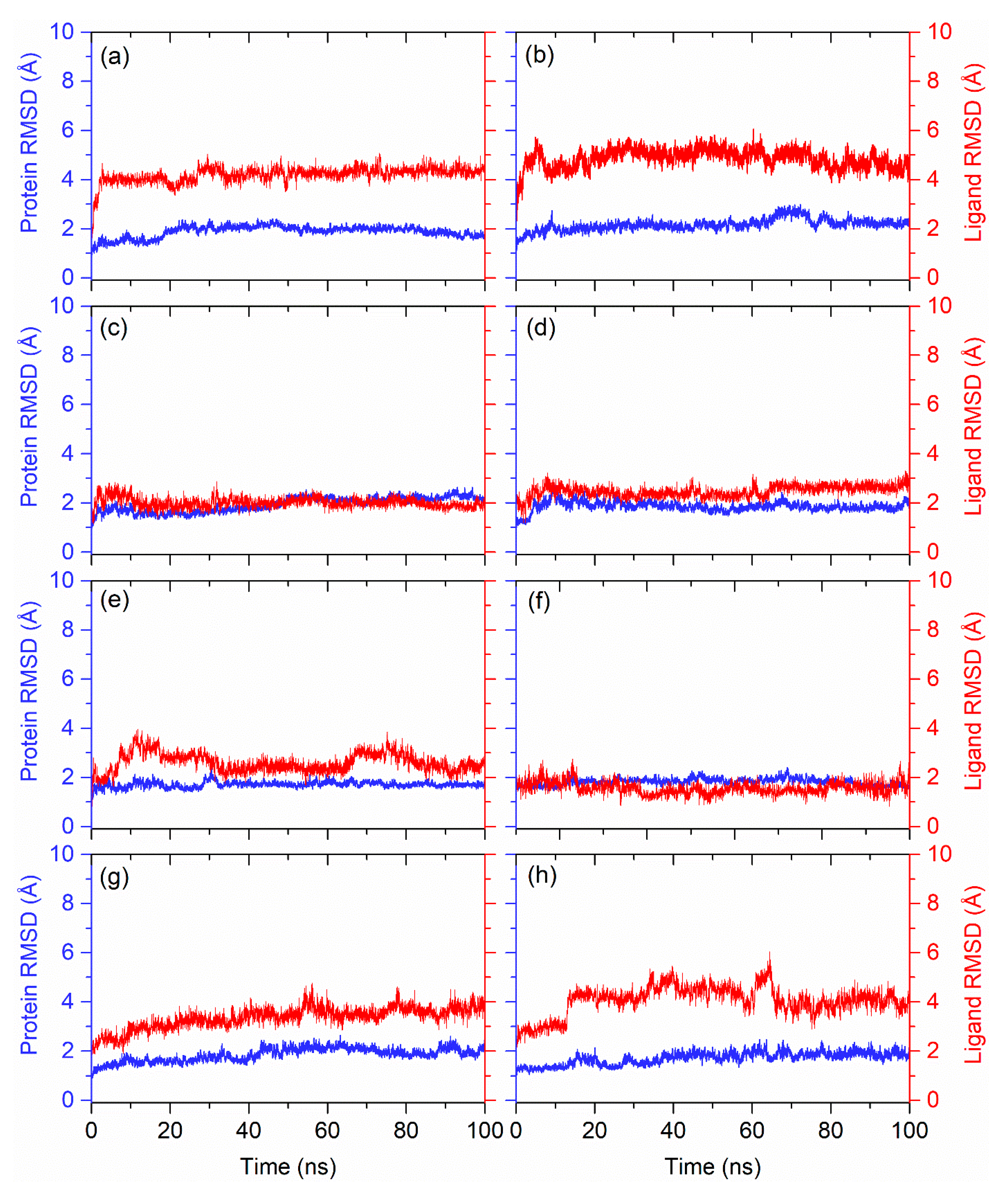
3.3. Molecular Dynamics Simulation Analysis
3.4. Post-Simulation Analysis
3.4.1. Principal Component Analysis
3.4.2. Binding Free Energy Calculation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uchil, P.D.; Satchidanandam, V. Phylogenetic analysis of Japanese encephalitis virus: Envelope gene based analysis reveals a fifth genotype, geographic clustering, and multiple introductions of the virus into the Indian subcontinent. Am. J. Trop. Med. Hyg. 2001, 65, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Fu, S.H.; Chen, W.X.; Wang, H.Y.; Guo, Y.H.; Liu, Q.Y.; Li, Y.X.; Luo, H.M.; Da, W.; Ji, D.Z.D.; et al. Genotype V Japanese Encephalitis Virus Is Emerging. PLoS Negl. Trop. Dis. 2011, 5, e1231. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Ni, H.; Beasley, D.W.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D. Origin and evolution of Japanese encephalitis virus in southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turtle, L.; Solomon, T. Japanese encephalitis—The prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef]
- Chen, K.M.; Tsai, H.C.; Sy, C.L.; Lee, S.S.J.; Liu, Y.C.; Wann, S.R.; Wang, Y.H.; Mai, M.H.; Chen, J.K.; Wu, K.S.; et al. Clinical manifestations of Japanese encephalitis in southern Taiwan. J. Microbiol. Immunol. 2009, 42, 296–302. [Google Scholar]
- Paul, W.S.; Moore, P.S.; Karabatsos, N.; Flood, S.P.; Yamada, S.; Jackson, T.; Tsai, T.F. Outbreak of Japanese encephalitis on the island of Saipan, 1990. J. Infect. Dis. 1993, 167, 1053–1058. [Google Scholar] [CrossRef]
- Wakai, S. Scourge of Japanese encephalitis in south-western Nepal. Lancet 1998, 351, 759. [Google Scholar] [CrossRef]
- Unni, S.K.; Ruzek, D.; Chhatbar, C.; Mishra, R.; Johri, M.K.; Singh, S.K. Japanese encephalitis virus: From genome to infectome. Microbes Infect. 2011, 13, 312–321. [Google Scholar] [CrossRef]
- Solomon, T.; Dung, N.M.; Kneen, R.; Gainsborough, M.; Vaughn, D.W.; Khanh, V.T. Japanese encephalitis. J. Neurol. Neurosurg. Psychiatry 2000, 68, 405. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, W.; Pan, Y.; Wang, G.; Yin, Q.; Fu, S.; Li, F.; He, Y.; Xu, S.; Wang, Z.; et al. A Bibliometric Analysis of Global Research on Japanese Encephalitis From 1934 to 2020. Front. Cell Infect. Microbiol. 2022, 12, 833701. [Google Scholar] [CrossRef]
- WHO. Japanese Encephalitis. WHO Fact. Sheets. 2019. Available online: https://www.who.int/en/news-room/fact-sheets/detail/japanese-encephalitis (accessed on 30 September 2021).
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774, 774A–774E. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccines Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovanich, D.; Saisawang, C.; Sittipaisankul, P.; Ramphan, S.; Kalpongnukul, N.; Somparn, P.; Pisitkun, T.; Smith, D.R. Analysis of the Zika and Japanese Encephalitis Virus NS5 Interactomes. J. Proteome. Res. 2019, 18, 3203–3218. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Verma, A.; Yadav, P.; Dubey, S.K.; Azhar, E.I.; Maitra, S.S.; Dwivedi, V.D. Molecular pathogenesis of Japanese encephalitis and possible therapeutic strategies. Arch. Virol. 2022, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Zhang, L.W.; Ramachandra, M.; Kusukawa, J.; Ebner, K.E.; Padmanabhan, R. Association between Ns3 and Ns5 Proteins of Dengue Virus Type-2 in the Putative Rna Replicase Is Linked to Differential Phosphorylation of Ns5. J. Biol. Chem. 1995, 270, 19100–19106. [Google Scholar] [CrossRef] [Green Version]
- Uchil, P.D.; Kumar, A.V.; Satchidanandam, V. Nuclear localization of flavivirus RNA synthesis in infected cells. J. Virol. 2006, 80, 5451–5464. [Google Scholar] [CrossRef] [Green Version]
- Selisko, B.; Wang, C.; Harris, E.; Canard, B. Regulation of Flavivirus RNA synthesis and replication. Curr. Opin. Virol. 2014, 9, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Hodel, A.E.; Gershon, P.D.; Shi, X.N.; Quiocho, F.A. The 1.85 angstrom structure of vaccinia protein VP39: A bifunctional enzyme that participates in the modification of both mRNA ends. Cell 1996, 85, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Egloff, M.P.; Benarroch, D.; Selisko, B.; Romette, J.L.; Canard, B. An RNA cap (nucleoside-2′-O-)-methyltransferase in the flavivirus RNA polymerase NS5: Crystal structure and functional characterization. EMBO J. 2002, 21, 2757–2768. [Google Scholar] [CrossRef] [Green Version]
- El Sahili, A.; Soh, T.S.; Schiltz, J.; Gharbi-Ayachi, A.; Seh, C.C.; Shi, P.Y.; Lim, S.P.; Lescar, J. NS5 from Dengue Virus Serotype 2 Can Adopt a Conformation Analogous to That of Its Zika Virus and Japanese Encephalitis Virus Homologues. J. Virol. 2019, 94, e01294-19. [Google Scholar] [CrossRef]
- Surana, P.; Satchidanandam, V.; Nair, D.T. RNA-dependent RNA polymerase of Japanese encephalitis virus binds the initiator nucleotide GTP to form a mechanistically important pre-initiation state. Nucleic Acids Res. 2014, 42, 2758–2773. [Google Scholar] [CrossRef]
- Hoke, C.H.; Nisalak, A.; Sangawhipa, N.; Jatanasen, S.; Laorakapongse, T.; Innis, B.L.; Kotchasenee, S.; Gingrich, J.B.; Latendresse, J.; Fukai, K.; et al. Protection against Japanese encephalitis by inactivated vaccines. N. Engl. J. Med. 1988, 319, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.Y.; Ming, Z.G.; Peng, G.Y.; Jian, A.; Min, L.H. Safety of a live-attenuated Japanese encephalitis virus vaccine (SA14-14-2) for children. Am. J. Trop. Med. Hyg. 1988, 39, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.C.; Chen, Y.Y.; Chen, J.M.; Huang, C.; Huang, M.; Chiou, S.S. Effectiveness of Live-Attenuated Genotype III Japanese Encephalitis Viral Vaccine against Circulating Genotype I Viruses in Swine. Viruses 2022, 14, 114. [Google Scholar] [CrossRef] [PubMed]
- Ngwe Tun, M.M.; Kyaw, A.K.; Nwe, K.M.; Inoue, S.; Thant, K.Z.; Morita, K. Effectiveness of the SA 14-14-2 Live-Attenuated Japanese Encephalitis Vaccine in Myanmar. Vaccines 2021, 9, 568. [Google Scholar] [CrossRef] [PubMed]
- Tandale, B.V.; Khan, S.A.; Kushwaha, K.P.; Rahman, H.; Gore, M.M.; Japanese Encephalitis Vaccination Efficacy Case Control Study, G. Effectiveness of Japanese encephalitis SA 14-14-2 live attenuated vaccine among Indian children: Retrospective 1:4 matched case-control study. J. Infect. Public Health 2018, 11, 713–719. [Google Scholar] [CrossRef]
- Hennessy, S.; Liu, Z.; Tsai, T.F.; Strom, B.L.; Wan, C.M.; Liu, H.L.; Wu, T.X.; Yu, H.J.; Liu, Q.M.; Karabatsos, N.; et al. Effectiveness of live-attenuated Japanese encephalitis vaccine (SA14-14-2): A case-control study. Lancet 1996, 347, 1583–1586. [Google Scholar] [CrossRef]
- Gao, N.; Chen, W.; Zheng, Q.; Fan, D.Y.; Zhang, J.L.; Chen, H.; Gao, G.F.; Zhou, D.S.; An, J. Co-expression of Japanese encephalitis virus prM-E-NS1 antigen with granulocyte-macrophage colony-stimulating factor enhances humoral and anti-virus immunity after DNA vaccination. Immunol. Lett. 2010, 129, 23–31. [Google Scholar] [CrossRef]
- Wu, S.F.; Lee, C.J.; Liao, C.L.; Dwek, R.A.; Zitzmann, N.; Lin, Y.L. Antiviral effects of an iminosugar derivative on flavivirus infections. J. Virol. 2002, 76, 3596–3604. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Ou, Y.C.; Raung, S.L.; Chen, C.J. Antiviral effect of dehydroepiandrosterone on Japanese encephalitis virus infection. J. Gen. Virol. 2005, 86, 2513–2523. [Google Scholar] [CrossRef] [Green Version]
- Sebastian, L.; Desai, A.; Shampur, M.N.; Perumal, Y.; Sriram, D.; Vasanthapuram, R. N-methylisatin-beta-thiosemicarbazone derivative (SCH 16) is an inhibitor of Japanese encephalitis virus infection in vitro and in vivo. Virol. J. 2008, 5, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.J.; Chang, Y.C.; Lu, K.Z.; Tsou, Y.Y.; Lin, C.W. Antiviral Activity of Isatis indigotica Extract and Its Derived Indirubin against Japanese Encephalitis Virus. Evid.-Based Complement. Altern. Med 2012, 2012, 925830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Liu, Y.; Guo, J.; Wang, P.; Zhang, L.; Xiao, G.; Wang, W. Screening of FDA-Approved Drugs for Inhibitors of Japanese Encephalitis Virus Infection. J. Virol. 2017, 91, e01055-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawa, M.; Takasaki, T.; Yamada, K.I.; Kurane, I.; Akatsuka, T. Interference in Japanese encephalitis virus infection of Vero cells by a cationic amphiphilic drug, chlorpromazine. J. Gen. Virol. 2003, 84, 1737–1741. [Google Scholar] [CrossRef]
- Ye, J.; Jiang, R.; Cui, M.; Zhu, B.; Sun, L.; Wang, Y.; Zohaib, A.; Dong, Q.; Ruan, X.; Song, Y.; et al. Etanercept reduces neuroinflammation and lethality in mous.se model of Japanese encephalitis. J. Infect. Dis. 2014, 210, 875–889. [Google Scholar] [CrossRef]
- Mishra, M.K.; Basu, A. Minocycline neuroprotects, reduces microglial activation, inhibits caspase 3 induction, and viral replication following Japanese encephalitis. J. Neurochem. 2008, 105, 1582–1595. [Google Scholar] [CrossRef]
- Dwivedi, V.D.; Singh, A.; El-Kafraway, S.A.; Alandijany, T.A.; Faizo, A.A.; Bajrai, L.H.; Kamal, M.A.; Azhar, E.I. Mechanistic insights into the Japanese encephalitis virus RNA dependent RNA polymerase protein inhibition by bioflavonoids from Azadirachta indica. Sci. Rep. 2021, 11, 18125. [Google Scholar] [CrossRef]
- Dwivedi, V.D.; Bharadwaj, S.; Afroz, S.; Khan, N.; Ansari, M.A.; Yadava, U.; Tripathi, R.C.; Tripathi, I.P.; Mishra, S.K.; Kang, S.G. Anti-dengue infectivity evaluation of bioflavonoid from Azadirachta indica by dengue virus serine protease inhibition. J. Biomol. Struct. Dyn. 2021, 39, 1417–1430. [Google Scholar] [CrossRef]
- Dwivedi, V.D.; Tripathi, I.P.; Tripathi, R.C.; Singh, R.; Yadava, U.; Mishra, S.K. In silico docking of quercetin-3-O-β-D-glucoside from Azadirachta indica with NS2B-NS3 protease in Dengue virus. Online J. Bioinform. 2018, 19, 175–180. [Google Scholar]
- Ali, S.I.; Sheikh, W.M.; Rather, M.A.; Venkatesalu, V.; Muzamil Bashir, S.; Nabi, S.U. Medicinal plants: Treasure for antiviral drug discovery. Phytother. Res. PTR 2021, 35, 3447–3483. [Google Scholar] [CrossRef]
- Ganjhu, R.K.; Mudgal, P.P.; Maity, H.; Dowarha, D.; Devadiga, S.; Nag, S.; Arunkumar, G. Herbal plants and plant preparations as remedial approach for viral diseases. Virusdisease 2015, 26, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Nedyalkova, M.; Vasighi, M.; Sappati, S.; Kumar, A.; Madurga, S.; Simeonov, V. Inhibition Ability of Natural Compounds on Receptor-Binding Domain of SARS-CoV2: An In Silico Approach. Pharmaceuticals 2021, 14, 1328. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, S.; Kumar, A. Screening of phytoconstituents of Andrographis paniculata against various targets of Japanese encephalitis virus: An in-silico and in-vitro target-based approach. Curr. Res. Pharmacol. Drug Discov. 2021, 2, 100043. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, K.; Takagi, Y.; Ueba, N.; Yamasaki, K.; Sakagami, Y.; Yokoyama, H.; Yoneda, K. Protective effect of astragali radix by oral administration against Japanese encephalitis virus infection in mice. Biol. Pharm. Bull. 1996, 19, 1166–1169. [Google Scholar] [CrossRef] [Green Version]
- Swarup, V.; Ghosh, J.; Mishra, M.K.; Basu, A. Novel strategy for treatment of Japanese encephalitis using arctigenin, a plant lignan. J. Antimicrob. Chemoth. 2008, 61, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Johari, J.; Kianmehr, A.; Mustafa, M.R.; Abubakar, S.; Zandi, K. Antiviral activity of baicalein and quercetin against the Japanese encephalitis virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef]
- Barnes, J.; Anderson, L.A.; Gibbons, S.; Phillipso.on, J.D. Echinacea species (Echinacea angustifolia (DC.) Hell., Echinacea pallida (Nutt.) Nutt., Echinacea purpurea (L.) Moench): A review of their chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2005, 57, 929–954. [Google Scholar] [CrossRef] [Green Version]
- Signer, J.; Jonsdottir, H.R.; Albrich, W.C.; Strasser, M.; Zust, R.; Ryter, S.; Ackermann-Gaumann, R.; Lenz, N.; Siegrist, D.; Suter, A.; et al. In vitro virucidal activity of Echinaforce(R), an Echinacea purpurea preparation, against coronaviruses, including common cold coronavirus 229E and SARS-CoV-2. Virol. J. 2020, 17, 136. [Google Scholar] [CrossRef]
- Aucoin, M.; Cooley, K.; Saunders, P.R.; Cardozo, V.; Remy, D.; Cramer, H.; Abad, C.N.; Hannan, N. The effect of quercetin on the prevention or treatment of COVID-19 and other respiratory tract infections in humans: A rapid review. Adv. Integr. Med. 2020, 7, 247–251. [Google Scholar] [CrossRef]
- Hudson, J.; Vimalanathan, S. Echinacea—A Source of Potent Antivirals for Respiratory Virus Infections. Pharmaceuticals 2011, 4, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, S.; El-Kafrawy, S.A.; Alandijany, T.A.; Bajrai, L.H.; Shah, A.A.; Dubey, A.; Sahoo, A.K.; Yadava, U.; Kamal, M.A.; Azhar, E.I.; et al. Structure-Based Identification of Natural Products as SARS-CoV-2 M(pro) Antagonist from Echinacea angustifolia Using Computational Approaches. Viruses 2021, 13, 305. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.B. Applications of the phytomedicine Echinacea purpurea (Purple Coneflower) in infectious diseases. J. Biomed. Biotechnol. 2012, 2012, 769896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Lee, K.E.; Dwivedi, V.D.; Yadava, U.; Nees, M.; Kang, S.G. Density functional theory and molecular dynamics simulation support Ganoderma lucidum triterpenoids as broad range antagonist of matrix metalloproteinases. J. Mol. Liq. 2020, 311, 113322. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger Release 2020-4; Maestro, Schrödinger, LLC: New York, NY, USA, 2020.
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D. Scalable algorithms for molecular dynamics simulations on commodity clusters. In Proceedings of the SC’06: Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; p. 43. [Google Scholar]
- Schrödinger Release 2018-4: Desmond Molecular Dynamics System; D. E. Shaw Research: New York, NY, USA; Maestro-Desmond Interoperability Tools, Schrödinger: New York, NY, USA, 2018.
- Martyna, G.J.; Klein, M.L.; Tuckerman, M. Nose-Hoover Chains—The Canonical Ensemble Via Continuous Dynamics. J. Chem. Phys. 1992, 97, 2635–2643. [Google Scholar] [CrossRef]
- Toukmaji, A.Y.; Board, J.A. Ewald summation techniques in perspective: A survey. Comput. Phys. Commun. 1996, 95, 73–92. [Google Scholar] [CrossRef]
- Grant, B.J.; Rodrigues, A.P.; ElSawy, K.M.; McCammon, J.A.; Caves, L.S. Bio3d: An R package for the comparative analysis of protein structures. Bioinformatics 2006, 22, 2695–2696. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Compound | Docking Score (kcal/mol) | H-Bond | π–π/π–Cation Stacking | Hydrophobic | Polar | Negative | Positive | Glycine | Salt Bridge |
|---|---|---|---|---|---|---|---|---|---|---|
| a. | Echinacoside | −11.1 | Arg460(2), Glu461, Ser604(2), Ser666, Asp669 | Arg474 | Ile476, Val509, Tyr610, Phe713, Cys714, Trp800, Ile802 | Ser473, Ser604, Thr609, Asn613, Ser666, Ser801, His803 | Glu461, Asp668, Asp669 | Arg460, Lys462, Lys463, Lys471, Arg474, Arg742 | Gly412, Gly472, Gly605, Gly667 | - |
| b. | Echinacin | −10.4 | Leu411, Tyr610, Cys714, Ser715, Arg734, Arg742, Ser801 | - | Leu411, Ala413, Ile476, Val607, Tyr610, Cys714, Trp800, Ile802 | Ser604, Gln606, Thr609, Ser715, Asn716, Ser799 Ser801, His803 | Glu738 | Arg460, Arg474, Arg734, Arg742, | Gly412 | - |
| c. | Rutin | −10.4 | Lyn463, Ala475, Ser604, Gln606, Ser666, Asp668 | Tyr610 | Ala413, Val414, Ala475, Ile476, Trp477, Val607, Tyr610, Cys714, Trp800, Ile802 | Ser604, Gln606, Thr609, Ser666, Ser715, Ser801, His803 | Asp668, Asp669 | Arg460, Lys471, Arg474, Arg742 | Gly412, Gly605, Gly667 | - |
| d. | Cynaroside | −10.0 | Ser666, Asp668, Asp669, His803 | - | Leu411, Ala413, Val607, Tyr610, Ala611, Cys714, Ile802 | Asn495, Ser604, Gln606, Thr609, Asn613, Ser666, Ser801, His803 | Glu510, Asp668, Asp669 | Lys404, Lys471, Hip498 | Gly412, Gly605, Gly667 | - |
| e. | Quercetagetin 7-glucoside | −10.0 | Ser666, Asp668, Ser801 | - | Leu411, Ala413, Val607, Tyr610, Ala611, Cys714, Ile802 | Asn495, Ser604, Gln606, Thr609, Asn613, Ser666, Ser801, His803 | Glu510, Asp668, Asp669 | Lys404, Lys471, Arg474 | Gly412, Gly605, Gly667 | - |
| f. | Kaempferol-3-glucoside | −10.0 | Asp541, Ser604, Ser801 | - | Leu411, Ala413, Trp540, Val607, Tyr610, Trp800, Ile802 | Asn495, Ser604, Gln605, Thr609, Asn613, Ser666, Ser801 | Asp541, Asp668 | Arg474 | Gly412, Gly667, | - |
| G | Adenosine triphosphate | −8.6 | Ser604, Gln606, Ser715, Arg742, Trp800, Ile802 | -- | Leu411, Ala413, Val607, Tyr610, Trp800, Ile802 | Ser604, Gln606, Thr609, Ser715, Ser799, Ser801 | - | Arg460, Lys463, Lys471, Arg474, Arg734, Arg742 | Gly412, Gly605 | Lys463, Arg474, Arg734, Arg742 |
| h | Guanosine-5′-triphosphate | −9.0 | Asp541, Asn613, Asp668, Ser801, Ile802 | Tyr610 | Trp540, Tyr610, Trp800, Ile802 | Ser604, Asn613, Thr609, Ser666, Ser801, His803 | Asp541, Asp668, Asp669 | Arg460, Lys463, Arg474, Arg742 | Gly667 | Arg460, Lys463, Arg474, Arg742 |
| Energy (kcal/mol) | JEV–RdRP Poses with Natural Compounds from Echinacea angustifolia | |||||||
|---|---|---|---|---|---|---|---|---|
| Echinacoside | Echinacin | Rutin | Cynaroside | Quercetagetin 7-Glucoside | Kaempferol-3-Glucoside | Adenosine Triphosphate | Guanosine-5′-Triphosphate | |
| ΔGBind | −80.15 ± 8.04 | −81.67 ± 8.31 | −80.33 ± 5.54 | −57.26 ± 4.13 | −66.24 ± 5.6 | −62.13 ± 5.97 | −69.62 ± 8.62 | −47.98 ± 11.41 |
| ΔGBindCoulomb | −57.04 ± 8.1 | −32.93 ± 8.82 | −41.87 ± 5.69 | −28.94 ± 6.36 | −30.43 ± 6.17 | −28.68 ± 5.86 | −242.97 ± 54.21 | −242.62 ± 52.45 |
| ΔGBindCovalent | 4.62 ± 2.38 | 4.17 ± 2.11 | 5.19 ± 2.51 | 6.06 ± 2.1 | 4.29 ± 1.87 | 3.94 ± 1.67 | 7.83 ± 4.45 | 5.13 ± 2.45 |
| ΔGBindHbond | −7.11 ± 1 | −3.95 ± 0.55 | −6.11 ± 0.69 | −3.68 ± 0.79 | −4.56 ± 0.92 | −3.46 ± 0.63 | −15.77 ± 2.08 | −10.65 ± 2.24 |
| ΔGBindLipo | −18.26 ± 2.1 | −16.1 ± 1.22 | −17.43 ± 1.37 | −13.59 ± 1.49 | −16.4 ± 1.04 | −16.22 ± 1.34 | −5.09 ± 0.58 | −3.56 ± 0.61 |
| ΔGBindPacking | −2.04 ± 0.63 | −7.16 ± 1.27 | −0.71 ± 0.17 | −3.35 ± 0.62 | −2.07 ± 0.35 | −2.38 ± 0.76 | −0.15 ± 0.28 | −1.48 ± 0.61 |
| ΔGBindSolvGB | 66.69 ± 6.11 | 41.52 ± 3.6 | 46.21 ± 3.44 | 42.08 ± 3.18 | 42.84 ± 4.72 | 39.37 ± 3.82 | 224.6 ± 49.37 | 236.56 ± 49.6 |
| ΔGBindvdW | −67 ± 4.05 | −67.22 ± 3.38 | −65.54 ± 3.71 | −55.81 ± 3.28 | −59.9 ± 3.17 | −54.7 ± 3.41 | −38.07 ± 4.69 | −31.35 ± 4.63 |
| Ligand Strain Energy | 11.92 ± 3.73 | 7.91 ± 2.37 | 10.64 ± 3.46 | 6.0 ±2.13 | 5.2 ± 2.19 | 5.57 ± 2.51 | 7.55 ± 4.06 | 3.99 ± 2.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, P.; El-Kafrawy, S.A.; El-Day, M.M.; Alghafari, W.T.; Faizo, A.A.; Jha, S.K.; Dwivedi, V.D.; Azhar, E.I. Discovery of Small Molecules from Echinacea angustifolia Targeting RNA-Dependent RNA Polymerase of Japanese Encephalitis Virus. Life 2022, 12, 952. https://doi.org/10.3390/life12070952
Yadav P, El-Kafrawy SA, El-Day MM, Alghafari WT, Faizo AA, Jha SK, Dwivedi VD, Azhar EI. Discovery of Small Molecules from Echinacea angustifolia Targeting RNA-Dependent RNA Polymerase of Japanese Encephalitis Virus. Life. 2022; 12(7):952. https://doi.org/10.3390/life12070952
Chicago/Turabian StyleYadav, Pardeep, Sherif A. El-Kafrawy, Mai M. El-Day, Wejdan T. Alghafari, Arwa A. Faizo, Saurabh Kumar Jha, Vivek Dhar Dwivedi, and Esam I. Azhar. 2022. "Discovery of Small Molecules from Echinacea angustifolia Targeting RNA-Dependent RNA Polymerase of Japanese Encephalitis Virus" Life 12, no. 7: 952. https://doi.org/10.3390/life12070952
APA StyleYadav, P., El-Kafrawy, S. A., El-Day, M. M., Alghafari, W. T., Faizo, A. A., Jha, S. K., Dwivedi, V. D., & Azhar, E. I. (2022). Discovery of Small Molecules from Echinacea angustifolia Targeting RNA-Dependent RNA Polymerase of Japanese Encephalitis Virus. Life, 12(7), 952. https://doi.org/10.3390/life12070952