
A Comprehensive Review on the Anti-Cancer Effects of Oleuropein





Abstract
:1. Introduction

2. Pharmacokinetics of Ole
2.1. Dose of Ole
2.2. Metabolism of Ole
3. Role of Ole in Cancer
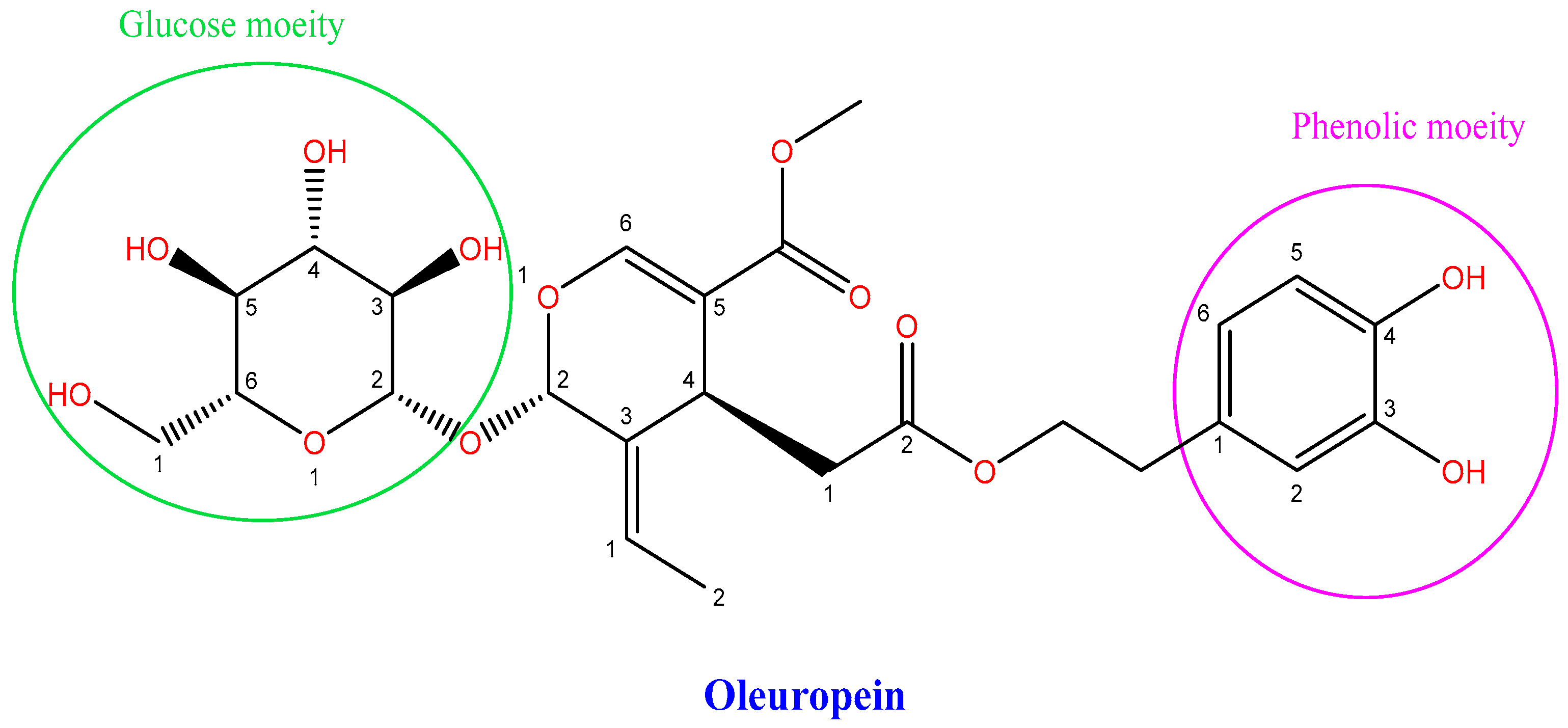
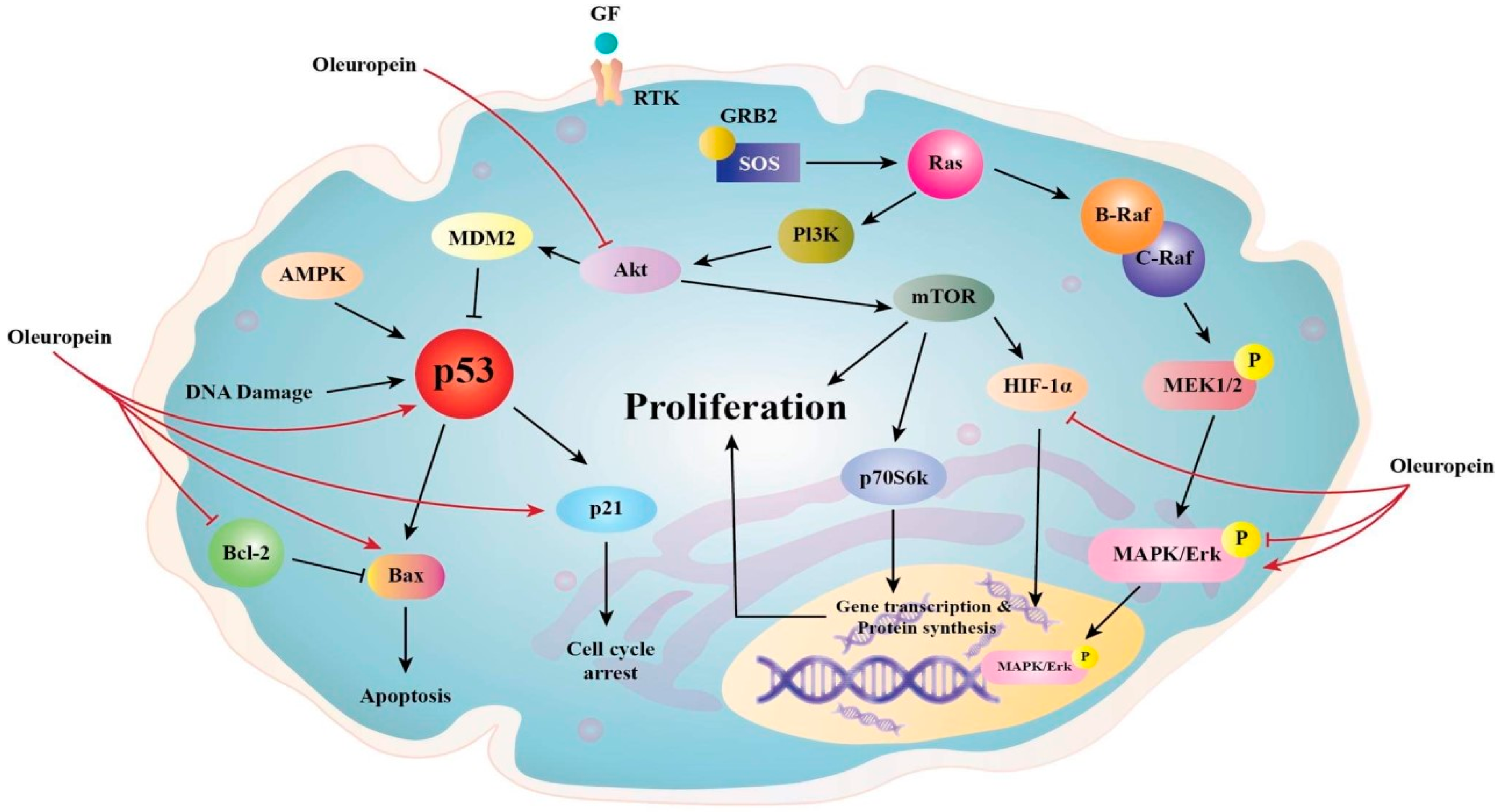
3.1. Anti-Proliferative Effects of Ole

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Vivo/In Vitro | Type of Cancer/Cell | Anti-Proliferative Effects of Ole | References |
|---|---|---|---|
| MCF-7 in vitro | Breast cancer cells |
| [25] |
| MDA-MB-231 in vitro | Breast cancer cells |
| [28] |
| TCAM-2 and SEM-1 in vitro | Seminoma Cells |
| [30] |
| HT 29 in vitro | Colon cancer cells |
| [31] |
| U251 in vitro | Glioblastoma cells |
| [45] |
| A172 in vitro | Glioblastoma cells |
| [45] |
| HepG2 in vitro | Hepatocarcinoma cells |
| [46] |
| TPC-1 and BCPAP in vitro | Thyroid cancer cell |
| [49] |
| LNCaP and DU145 in vitro | Prostate cancer cell |
| [52] |
| Caco-2 cells in vitro | Human colon cell |
| [54] |
| SH-SY5Y in vitro | Neuroblastoma cells |
| [55] |
3.2. Anti-Angiogenic and Apoptotic of Ole
| In Vivo/In Vitro | Type of Cancer/Cell | Anti-Angiogenic and Apoptotic of Ole | References |
|---|---|---|---|
| In vivo | Breast cancer cells |
| [9] |
| MDA-MB-321 in vitro | Breast cancer cells |
| [14] |
| NSCLC H1299 in vitro | Lung cancer cells |
| [57] |
| MIDA-MB-468 in vitro | Breast cancer cells |
| [58] |
| MDA-MB-231 in vitro | Breast cancer cells |
| [58] |
| MIA PaCa-2 in vitro | Pancreatic cancer cells |
| [59] |
| TCAM-2 and SEM-1 in vitro | Seminoma Cells |
| [30] |
| NSCLC H1299 in vitro | Lung cancer cells |
| [57] |
| In vitro | Breast neuroblastoma |
| [55] |
| In vitro | Hela cervical cancer cells |
| [24] |
| MCF-7 in vitro | Breast cancer cells |
| [25] |
| MIA PaCa-2 in vitro | Pancreatic cancer cells |
| [59] |
| NSCLC H1299 in vitro | Lung cancer cells |
| [57] |
| MCF-7 in vitro | Breast cancer cells |
| [25] |
| In vitro | Hela cervical cancer cell |
| [25] |
| A549 in vitro | Lung cancer |
| [60] |
| SH-SY5Y in vitro | Neuroblastoma |
| [55] |
| HT-29 in vitro | Human colon adenocarcinoma cells |
| [31] |
| B16F10 in vivo | Melanoma cells |
| [61] |
| In vitro | Prostate cancer |
| [52] |
| HepG2 in vitro | Hepatocellular |
| [62] |
| In vitro | HeLa cervical carcinoma cells |
| [64] |
| NSCLC A549 in vitro | Non-small-cell lung cancer |
| [65] |
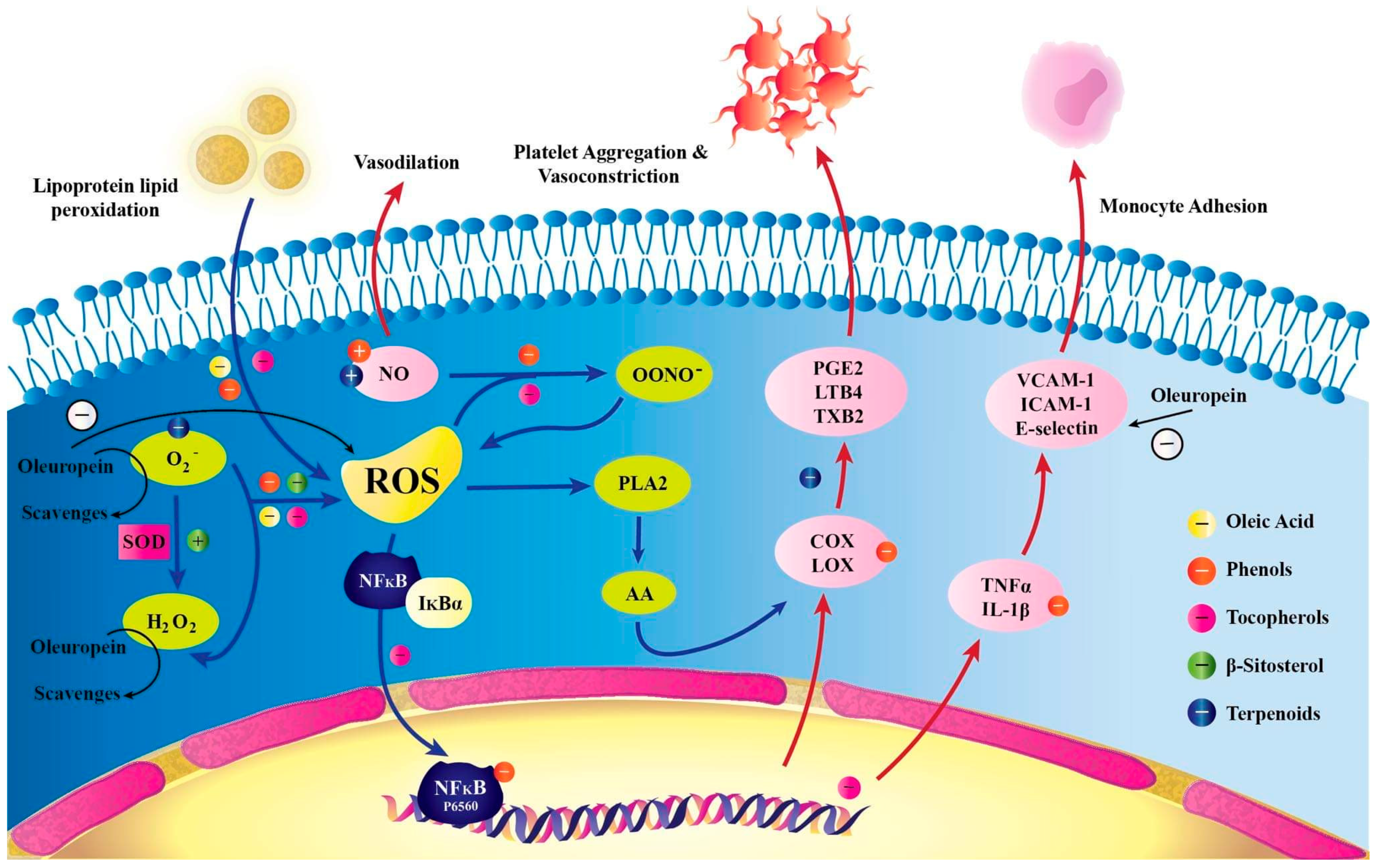
3.3. Antioxidant Properties of Ole
3.4. Ole and Cell Viability
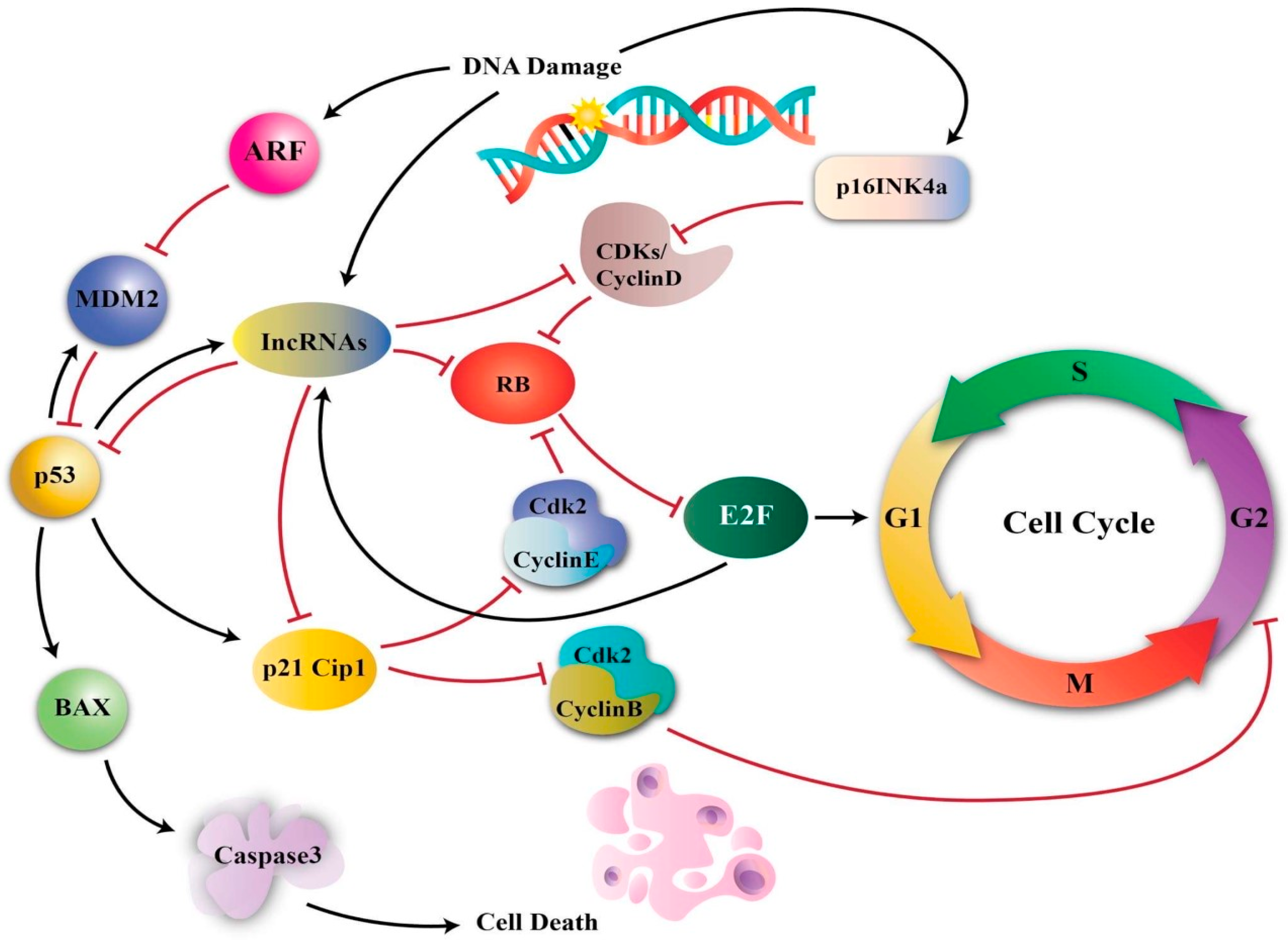
3.5. Ole and Cell Cycle Arrest
3.6. Ole as a Cytoskeleton Disruptor
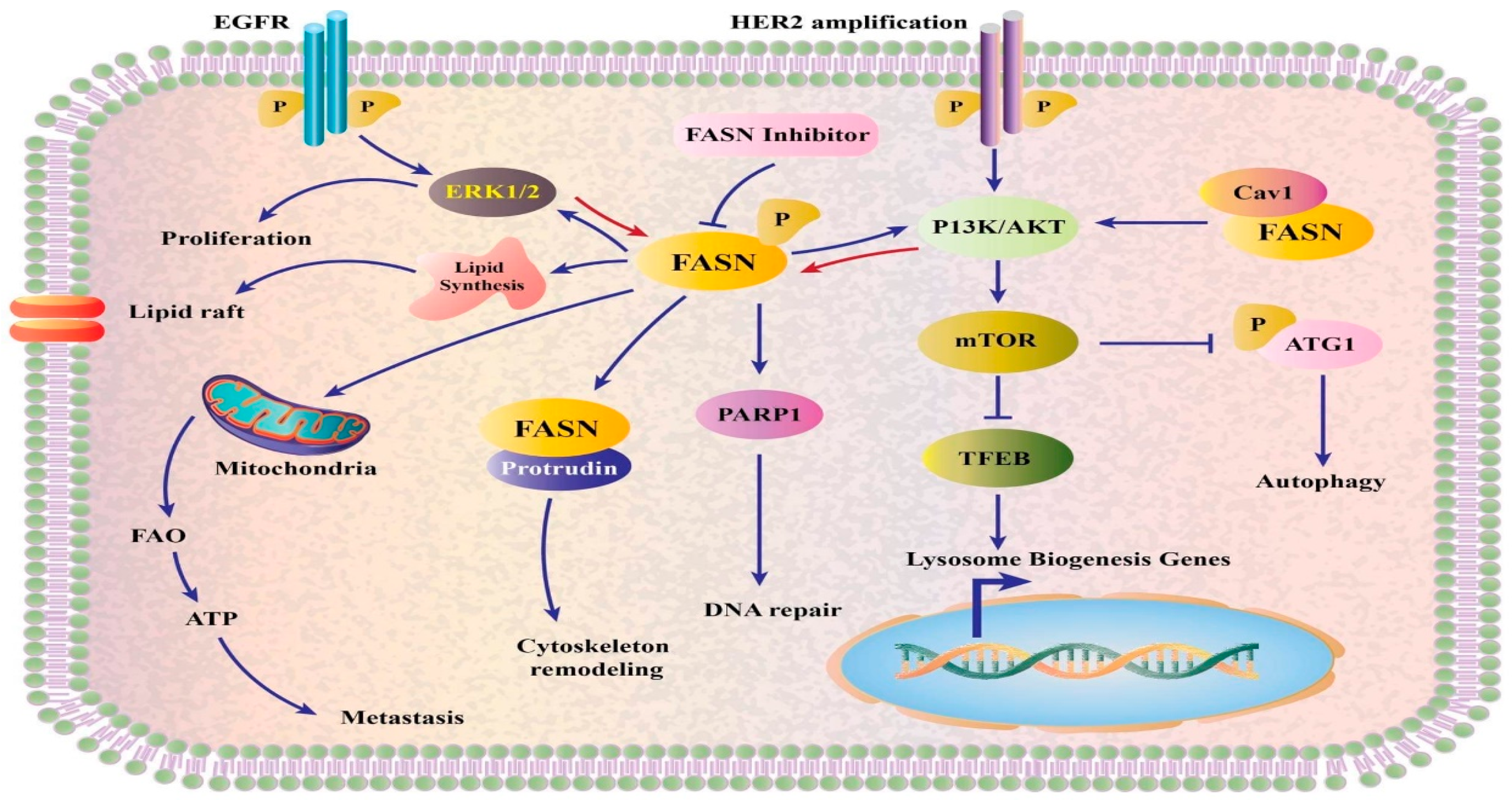
3.7. Ole and Fatty Acid Synthase
3.8. Ole and Inflammation
4. Derivatives of Ole
5. Ole in Combination with Chemotherapy
6. Summary and Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| Akt | Serine/threonine kinase |
| AMPK | 5′ adenosine monophosphate-activated protein kinase |
| ARF | Adenosine diphosphate-ribosylation factor |
| Atg1 | Autophagy-related 1 |
| Bad | BCL2-associated agonist of cell death |
| Bax | Bcl-2-associated X protein |
| Bcl2 | B-cell lymphoma 2 |
| Caspase | Cysteine proteases with aspartate specificity |
| CAV1 | Caveolin-1 |
| CB | Cannabinoid receptors |
| CREB | cAMP response element-binding protein |
| COX2 | Cyclooxygenase-2 |
| ERK | Extracellular signal-regulated kinase |
| ER+ | Estrogen receptor-positive |
| FAO | Fatty acid oxidation |
| FASN | Fatty acid synthase enzyme |
| GCS | Glutamyl cysteine synthetase |
| GF | Growth factor |
| GRB2 | Growth factor receptor-bound protein 2 |
| Glo2 | Glyoxalase 2 |
| GLUTs | Glucose transporters |
| GSH | Glutathione |
| HER2+ | Human epidermal growth factor receptor 2 |
| HIF-1α | Hypoxia-inducible factor-alpha |
| HER2+ | Human epidermal growth factor receptor 2 |
| HT | Hydroxytyrosol |
| ICAM-1 | Intercellular Adhesion Molecule 1 |
| IκBα | Nuclear factor of kappa light polypeptide gene enhancer in B-cell inhibitor alpha |
| Inos | Inducible nitric oxide synthase |
| JNK | Jun N-terminal kinase |
| lncRNA | Long non-coding RNAs |
| LTB4 | Lipoxygenase |
| MDM2 | Murine double minute 2 |
| MMP | Matrix metalloproteinase |
| MEK | Mitogen-activated protein kinase |
| mTOR | Mammalian target of rapamycin |
| NF-kB | Nuclear factor-light-chain-enhancer of activated B |
| NO | Nitric oxide |
| NSAIDs | Non-steroidal anti-inflammatory drugs |
| Ole | Oleuropein |
| ONOO− | Peroxynitrite |
| PARPs | Poly (ADP-ribose) polymerases |
| PGE2 | Prostaglandin E2 |
| PI3K | Phosphatidylinositol 3-kinase |
| PI4K | Phosphatidylinositol 4-kinase |
| PPAR | Peroxisome proliferator-activated receptors |
| P38 MAPK | Mitogen-activated protein kinases |
| P53 | Tumor protein P53 |
| p70S6K | 70-kDa ribosomal protein S6 kinase |
| RAF | Rapidly accelerated fibrosarcoma |
| ROS | Reactive oxygen species |
| RTK | Receptor tyrosine kinase |
| TFEB | Transcription factor EB |
| TN | Triple-negative |
| TXB2 | Thromboxane B2 |
| VCAM-1 | Vascular cell adhesion molecule 1 |
| VEGF | Vascular endothelial growth factor |
References
- Nediani, C.; Ruzzolini, J.; Romani, A.; Calorini, L. Oleuropein, a bioactive compound from Olea europaea L., as a potential preventive and therapeutic agent in non-communicable diseases. Antioxidants 2019, 8, 578. [Google Scholar] [CrossRef] [Green Version]
- Rahiman, S.; El-Metwally, T.H.; Shrivastava, D.; Tantry, M.N.; Tantry, B.A. Oleuropein and oleic acid: A novel emerging dietary target for human chronic diseases. Indian J. Biochem. Biophys. (IJBB) 2019, 56, 263–268. [Google Scholar]
- Bulotta, S.; Oliverio, M.; Russo, D.; Procopio, A. Biological Activity of Oleuropein and Its Derivatives; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Al-Asmari, K.; Al-Attar, A.; Mohamed, I.; Zeid, A. Potential health benefits and components of olive oil: An overview. Biosci. Res. 2020, 17, 2673–2687. [Google Scholar]
- Shamshoum, H.; Vlavcheski, F.; Tsiani, E. Anticancer effects of oleuropein. Biofactors 2017, 43, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Moran, J.M.; Leal-Hernandez, O.; Canal-Macias, M.L.; Roncero-Martin, R.; Guerrero-Bonmatty, R.; Aliaga, I.; Zamorano, J.D.P. Antiproliferative properties of oleuropein in human osteosarcoma cells. Nat. Prod. Commun. 2016, 11, 491–492. [Google Scholar] [CrossRef] [Green Version]
- Omar, S.H. Cardioprotective and neuroprotective roles of oleuropein in olive. Saudi Pharm. J. 2010, 18, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breakspear, I.; Guillaume, C. A quantitative phytochemical comparison of olive leaf extracts on the Australian market. Molecules 2020, 25, 4099. [Google Scholar] [CrossRef] [PubMed]
- Milanizadeh, S.; Reza Bigdeli, M. Pro-apoptotic and anti-angiogenesis effects of olive leaf extract on spontaneous mouse mammary tumor model by balancing vascular endothelial growth factor and endostatin levels. Nutr. Cancer 2019, 71, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Emma, M.R.; Augello, G.; Di Stefano, V.; Azzolina, A.; Giannitrapani, L.; Montalto, G.; Cervello, M.; Cusimano, A. Potential uses of olive oil secoiridoids for the prevention and treatment of cancer: A narrative review of preclinical studies. Int. J. Mol. Sci. 2021, 22, 1234. [Google Scholar] [CrossRef]
- Horcajada, M.-N.; Beaumont, M.; Sauvageot, N.; Poquet, L.; Saboundjian, M.; Costes, B.; Verdonk, P.; Brands, G.; Brasseur, J.; Urbin-Choffray, D. An oleuropein-based dietary supplement may improve joint functional capacity in older people with high knee joint pain: Findings from a multicentre-RCT and post hoc analysis. Ther. Adv. Musculoskelet. Dis. 2022, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, R.; Silvestri, R.; Loffredo, L.; Novo, M.; Cammisotto, V.; Castellani, V.; Bartimoccia, S.; Nocella, C.; Violi, F. Oleuropein, a component of extra virgin olive oil, lowers postprandial glycaemia in healthy subjects. Br. J. Clin. Pharmacol. 2018, 84, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Kendall, M.; Batterham, M.; Callahan, D.L.; Jardine, D.; Prenzler, P.D.; Robards, K.; Ryan, D. Randomized controlled study of the urinary excretion of biophenols following acute and chronic intake of olive leaf supplements. Food Chem. 2012, 130, 651–659. [Google Scholar] [CrossRef]
- Elamin, M.H.; Elmahi, A.B.; Daghestani, M.H.; Al-Olayan, E.M.; Al-Ajmi, R.A.; Alkhuriji, A.F.; Hamed, S.S.; Elkhadragy, M.F. Synergistic anti-breast-cancer effects of combined treatment with oleuropein and doxorubicin in vivo. Altern. Ther. Health Med. 2017, 23, 17–24. [Google Scholar]
- Katsoulieris, E.N. The olive leaf extract oleuropein exerts protective effects against oxidant-induced cell death, concurrently displaying pro-oxidant activity in human hepatocarcinoma cells. Redox Rep. 2016, 21, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Nadeem, M.; Gilani, S.A.; Khan, S.; Sajid, M.W.; Amir, R.M. Antitumor perspectives of oleuropein and its metabolite hydroxytyrosol: Recent updates. J. Food Sci. 2018, 83, 1781–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Villalba, R.; Larrosa, M.; Possemiers, S.; Tomás-Barberán, F.; Espín, J. Bioavailability of phenolics from an oleuropein-rich olive (Olea europaea) leaf extract and its acute effect on plasma antioxidant status: Comparison between pre-and postmenopausal women. Eur. J. Nutr. 2014, 53, 1015–1027. [Google Scholar] [CrossRef]
- Kano, S.; Komada, H.; Yonekura, L.; Sato, A.; Nishiwaki, H.; Tamura, H. Absorption, metabolism, and excretion by freely moving rats of 3, 4-DHPEA-EDA and related polyphenols from olive fruits (Olea europaea). J. Nutr. Metab. 2016, 2016, 9104208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annunziata, G.; Maisto, M.; Schisano, C.; Barrea, L.; Ciampaglia, R.; Novellinoet, E. Oleuropein as a novel anti-diabetic nutraceutical. Overv. Arch. Diabetes Obes. 2018, 1, 54–58. [Google Scholar] [CrossRef]
- De Bock, M.; Thorstensen, E.B.; Derraik, J.G.; Henderson, H.V.; Hofman, P.L.; Cutfield, W.S. Human absorption and metabolism of oleuropein and hydroxytyrosol ingested as olive (Olea europaea L.) leaf extract. Mol. Nutr. Food Res. 2013, 57, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Gikas, E.; Papadopoulos, N.; Tsarbopoulos, A. Kinetic study of the acidic hydrolysis of oleuropein, the major bioactive metabolite of olive oil. J. Liq. Chromatogr. Relat. Technol. 2006, 29, 497–508. [Google Scholar] [CrossRef]
- Galmés, S.; Reynés, B.; Palou, M.; Palou-March, A.; Palou, A. Absorption, distribution, metabolism, and excretion of the main olive tree phenols and polyphenols: A literature review. J. Agric. Food Chem. 2021, 69, 5281–5296. [Google Scholar] [CrossRef]
- Bonechi, C.; Donati, A.; Tamasi, G.; Pardini, A.; Rostom, H.; Leone, G.; Lamponi, S.; Consumi, M.; Magnani, A.; Rossi, C. Chemical characterization of liposomes containing nutraceutical compounds: Tyrosol, hydroxytyrosol and oleuropein. Biophys. Chem. 2019, 246, 25–34. [Google Scholar] [CrossRef]
- Rizzo, M.; Ventrice, D.; Giannetto, F.; Cirinnà, S.; Santagati, N.A.; Procopio, A.; Mollace, V.; Muscoli, C. Antioxidant activity of oleuropein and semisynthetic acetyl-derivatives determined by measuring malondialdehyde in rat brain. J. Pharm. Pharmacol. 2017, 69, 1502–1512. [Google Scholar] [CrossRef] [PubMed]
- Liman, R.; Çoban, F.; Ciğerci, I.; Bulduk, İ.; Bozkurt, S. Antiangiogenic and apoptotic effects of Oleuropein on breast cancer cells. Br. J. Pharm. Res 2017, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Talorete, T.P.; Yamada, P.; Isoda, H. Anti-proliferative and apoptotic effects of oleuropein and hydroxytyrosol on human breast cancer MCF-7 cells. Cytotechnology 2009, 59, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ci, Y.; Qiao, J.; Han, M. Molecular mechanisms and metabolomics of natural polyphenols interfering with breast cancer metastasis. Molecules 2016, 21, 1634. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Ahn, K.S.; Shanmugam, M.K.; Wang, H.; Shen, H.; Arfuso, F.; Chinnathambi, A.; Alharbi, S.A.; Chang, Y.; Sethi, G. Oleuropein induces apoptosis via abrogating NF-κB activation cascade in estrogen receptor–negative breast cancer cells. J. Cell. Biochem. 2019, 120, 4504–4513. [Google Scholar] [CrossRef]
- Brasier, A.R. The nuclear factor-κB–interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Bossio, S.; Perri, A.; Malivindi, R.; Giordano, F.; Rago, V.; Mirabelli, M.; Salatino, A.; Brunetti, A.; Greco, E.A.; Aversa, A. Oleuropein Counteracts Both the Proliferation and Migration of Intra-and Extragonadal Seminoma Cells. Nutrients 2022, 14, 2323. [Google Scholar] [CrossRef] [PubMed]
- Cárdeno, A.; Sánchez-Hidalgo, M.; Rosillo, M.A.; de la Lastra, C.A. Oleuropein, a secoiridoid derived from olive tree, inhibits the proliferation of human colorectal cancer cell through downregulation of HIF-1α. Nutr. Cancer 2013, 65, 147–156. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S. Signal Transduct. Target. Ther 2017, 2, e17023. [Google Scholar]
- Sheng, J.; Sun, H.; Yu, F.-B.; Li, B.; Zhang, Y.; Zhu, Y.-T. The role of cyclooxygenase-2 in colorectal cancer. Int. J. Med. Sci. 2020, 17, 1095. [Google Scholar] [CrossRef] [PubMed]
- Giner, E.; Recio, M.C.; Ríos, J.L.; Cerdá-Nicolás, J.M.; Giner, R.M. Chemopreventive effect of oleuropein in colitis-associated colorectal cancer in c57bl/6 mice. Mol. Nutr. Food Res. 2016, 60, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, F.; Bravo, S.; Cruzat, F.; Montecino, M.; De Ferrari, G.V. Wnt/β-catenin signaling enhances cyclooxygenase-2 (COX2) transcriptional activity in gastric cancer cells. PLoS ONE 2011, 6, e18562. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Liang, H.; Han, Y. Cyclooxygenase-2 and β-catenin as potential diagnostic and prognostic markers in endometrial cancer. Front. Oncol. 2020, 10, 56. [Google Scholar] [CrossRef]
- Vallée, A.; Lecarpentier, Y.; Vallée, J.-N. Targeting the canonical WNT/β-catenin pathway in cancer treatment using non-steroidal anti-inflammatory drugs. Cells 2019, 8, 726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casado-Díaz, A.; Anter, J.; Müller, S.; Winter, P.; Quesada-Gómez, J.M.; Dorado, G. Transcriptomic analyses of the anti-adipogenic effects of oleuropein in human mesenchymal stem cells. Food Funct. 2017, 8, 1254–1270. [Google Scholar] [CrossRef] [PubMed]
- Svobodova, M.; Andreadou, I.; Skaltsounis, A.-L.; Kopecky, J.; Flachs, P. Oleuropein as an inhibitor of peroxisome proliferator-activated receptor gamma. Genes Nutr. 2014, 9, 1–8. [Google Scholar] [CrossRef]
- Gou, Q.; Gong, X.; Jin, J.; Shi, J.; Hou, Y. Peroxisome proliferator-activated receptors (PPARs) are potential drug targets for cancer therapy. Oncotarget 2017, 8, 60704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leto, G.; Flandina, C.; Crescimanno, M.; Giammanco, M.; Sepporta, M.V. Effects of oleuropein on tumor cell growth and bone remodelling: Potential clinical implications for the prevention and treatment of malignant bone diseases. Life Sci. 2021, 264, 118694. [Google Scholar] [CrossRef]
- Ritter, B.; Greten, F.R. Modulating inflammation for cancer therapy. J. Exp. Med. 2019, 216, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Razali, R.A.; Lokanathan, Y.; Yazid, M.D.; Ansari, A.S.; Saim, A.B.; Idrus, R.B. Modulation of epithelial to mesenchymal transition signaling pathways by Olea europaea and its active compounds. Int. J. Mol. Sci. 2019, 20, 3492. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cao, X. Characteristics and significance of the pre-metastatic niche. Cancer Cell 2016, 30, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Wang, J.; Huang, B.; Chen, A.; Li, X. Oleuropein inhibits the proliferation and invasion of glioma cells via suppression of the AKT signaling pathway. Oncol. Rep. 2016, 36, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Sherif, I.O.; Al-Gayyar, M.M. Oleuropein potentiates anti-tumor activity of cisplatin against HepG2 through affecting proNGF/NGF balance. Life Sci. 2018, 198, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Winer, A.; Adams, S.; Mignatti, P. Matrix metalloproteinase inhibitors in cancer therapy: Turning past failures into future successes. Mol. Cancer Ther. 2018, 17, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of matrix metalloproteinases in angiogenesis and cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulotta, S.; Corradino, R.; Celano, M.; Maiuolo, J.; D’Agostino, M.; Oliverio, M.; Procopio, A.; Filetti, S.; Russo, D. Antioxidant and antigrowth action of peracetylated oleuropein in thyroid cancer cells. J. Mol. Endocrinol. 2013, 51, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Nitulescu, G.M.; Van De Venter, M.; Nitulescu, G.; Ungurianu, A.; Juzenas, P.; Peng, Q.; Olaru, O.T.; Grădinaru, D.; Tsatsakis, A.; Tsoukalas, D. The Akt pathway in oncology therapy and beyond. Int. J. Oncol. 2018, 53, 2319–2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acquaviva, R.; Di Giacomo, C.; Sorrenti, V.; Galvano, F.; Santangelo, R.; Cardile, V.; Gangia, S.; D’Orazio, N.; Abraham, N.G.; Vanella, L. Antiproliferative effect of oleuropein in prostate cell lines. Int. J. Oncol. 2012, 41, 31–38. [Google Scholar] [PubMed] [Green Version]
- Laezza, C.; Pagano, C.; Navarra, G.; Pastorino, O.; Proto, M.C.; Fiore, D.; Piscopo, C.; Gazzerro, P.; Bifulco, M. The endocannabinoid system: A target for cancer treatment. Int. J. Mol. Sci. 2020, 21, 747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Francesco, A.; Falconi, A.; Di Germanio, C.; Di Bonaventura, M.V.M.; Costa, A.; Caramuta, S.; Del Carlo, M.; Compagnone, D.; Dainese, E.; Cifani, C. Extravirgin olive oil up-regulates CB1 tumor suppressor gene in human colon cancer cells and in rat colon via epigenetic mechanisms. J. Nutr. Biochem. 2015, 26, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Seçme, M.; Eroğlu, C.; Dodurga, Y.; Bağcı, G. Investigation of anticancer mechanism of oleuropein via cell cycle and apoptotic pathways in SH-SY5Y neuroblastoma cells. Gene 2016, 585, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Walia, A.; Yang, J.F.; Huang, Y.-H.; Rosenblatt, M.I.; Chang, J.-H.; Azar, D.T. Endostatin’s emerging roles in angiogenesis, lymphangiogenesis, disease, and clinical applications. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2015, 1850, 2422–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wu, J.; Zhang, Q.; Li, X.; Zhu, X.; Wang, Q.; Cao, S.; Du, L. Mitochondria-mediated apoptosis was induced by oleuropein in H1299 cells involving activation of p38 MAP kinase. J. Cell. Biochem. 2019, 120, 5480–5494. [Google Scholar] [CrossRef]
- Messeha, S.S.; Zarmouh, N.O.; Asiri, A.; Soliman, K.F. Gene Expression Alterations Associated with Oleuropein-Induced Antiproliferative Effects and S-Phase Cell Cycle Arrest in Triple-Negative Breast Cancer Cells. Nutrients 2020, 12, 3755. [Google Scholar] [CrossRef]
- Goldsmith, C.D.; Bond, D.R.; Jankowski, H.; Weidenhofer, J.; Stathopoulos, C.E.; Roach, P.D.; Scarlett, C.J. The olive biophenols oleuropein and hydroxytyrosol selectively reduce proliferation, influence the cell cycle, and induce apoptosis in pancreatic cancer cells. Int. J. Mol. Sci. 2018, 19, 1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Zhu, X.; Du, L. P38 MAP kinase is involved in oleuropein-induced apoptosis in A549 cells by a mitochondrial apoptotic cascade. Biomed. Pharmacother. 2017, 95, 1425–1435. [Google Scholar] [CrossRef]
- Song, H.; Do Young Lim, J.I.J.; Cho, H.J.; Park, S.Y.; Kwon, G.T.; Kang, Y.-H.; Lee, K.W.; Choi, M.-S.; Park, J.H.Y. Dietary oleuropein inhibits tumor angiogenesis and lymphangiogenesis in the B16F10 melanoma allograft model: A mechanism for the suppression of high-fat diet-induced solid tumor growth and lymph node metastasis. Oncotarget 2017, 8, 32027. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.M.; Chai, E.Q.; Cai, H.Y.; Miao, G.Y.; Ma, W. Oleuropein induces apoptosis via activation of caspases and suppression of phosphatidylinositol 3-kinase/protein kinase B pathway in HepG2 human hepatoma cell line. Mol. Med. Rep. 2015, 11, 4617–4624. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Wu, W.; Jacevic, V.; Franca, T.C.; Wang, X.; Kuca, K. Selective inhibitors for JNK signalling: A potential targeted therapy in cancer. J. Enzym. Inhib. Med. Chem. 2020, 35, 574–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Wu, J.; Yang, X.; Yang, J.; Zhang, Y.; Du, L. Oleuropein induced apoptosis in HeLa cells via a mitochondrial apoptotic cascade associated with activation of the c-Jun NH2-terminal kinase. J. Pharmacol. Sci. 2014, 125, 300–311. [Google Scholar] [CrossRef] [Green Version]
- Antognelli, C.; Frosini, R.; Santolla, M.F.; Peirce, M.J.; Talesa, V.N. Oleuropein-induced apoptosis is mediated by mitochondrial glyoxalase 2 in NSCLC A549 cells: A mechanistic inside and a possible novel nonenzymatic role for an ancient enzyme. Oxidative Med. Cell. Longev. 2019, 2019, 8576961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgenstern, J.; Campos Campos, M.; Nawroth, P.; Fleming, T. The glyoxalase system—new insights into an ancient metabolism. Antioxidants 2020, 9, 939. [Google Scholar] [CrossRef]
- Bulotta, S.; Corradino, R.; Celano, M.; D’Agostino, M.; Maiuolo, J.; Oliverio, M.; Procopio, A.; Iannone, M.; Rotiroti, D.; Russo, D. Antiproliferative and antioxidant effects on breast cancer cells of oleuropein and its semisynthetic peracetylated derivatives. Food Chem. 2011, 127, 1609–1614. [Google Scholar] [CrossRef]
- Capo, C.R.; Pedersen, J.Z.; Falconi, M.; Rossi, L. Oleuropein shows copper complexing properties and noxious effect on cultured SH-SY5Y neuroblastoma cells depending on cell copper content. J. Trace Elem. Med. Biol. 2017, 44, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Denoyer, D.; Masaldan, S.; La Fontaine, S.; Cater, M.A. Targeting copper in cancer therapy:‘Copper That Cancer’. Metallomics 2015, 7, 1459–1476. [Google Scholar] [CrossRef]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Kucukgul, A.; Isgor, M.M.; Duzguner, V.; Atabay, M.N.; Kucukgul, A. Antioxidant effects of oleuropein on hydrogen peroxide-induced neuronal stress-an in vitro study. Anti-Inflamm. Anti-Allergy Agents Med. Chem. (Former. Curr. Med. Chem.-Anti-Inflamm. Anti-Allergy Agents) 2020, 19, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Koundouros, N.; Poulogiannis, G. Phosphoinositide 3-kinase/Akt signaling and redox metabolism in cancer. Front. Oncol. 2018, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Qie, S.; Diehl, J.A. Cyclin D1, cancer progression, and opportunities in cancer treatment. J. Mol. Med. 2016, 94, 1313–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal-Esteban, L.C.; Fajas, L. Cell cycle regulators in cancer cell metabolism. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165715. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, M.; Jones, M.F.; Lal, A. Long non-coding RNAs embedded in the Rb and p53 pathways. Cancers 2013, 5, 1655–1675. [Google Scholar] [CrossRef] [Green Version]
- Hamadi, H.; Castellon, R. Oleuropein, a non-toxic olive iridoid, is an anti-tumor agent and cytoskeleton disrupter. Biochem. Biophys. Res. Commun. 2005, 334, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Flavin, R.; Peluso, S.; Nguyen, P.L.; Loda, M. Fatty acid synthase as a potential therapeutic target in cancer. Future Oncol. 2010, 6, 551–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fhu, C.W.; Ali, A. Fatty acid synthase: An emerging target in cancer. Molecules 2020, 25, 3935. [Google Scholar] [CrossRef] [PubMed]
- Qabaha, K.; Al-Rimawi, F.; Qasem, A.; Naser, S.A. Oleuropein is responsible for the major anti-inflammatory effects of olive leaf extract. J. Med. Food 2018, 21, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Huguet-Casquero, A.; Moreno-Sastre, M.; López-Méndez, T.B.; Gainza, E.; Pedraz, J.L. Encapsulation of oleuropein in nanostructured lipid carriers: Biocompatibility and antioxidant efficacy in lung epithelial cells. Pharmaceutics 2020, 12, 429. [Google Scholar] [CrossRef]
- Castejon, M.L.; Sánchez-Hidalgo, M.; Aparicio-Soto, M.; González-Benjumea, A.; Fernández-Bolaños, J.G.; Alarcón-de-la-Lastra, C. Olive secoiridoid oleuropein and its semisynthetic acetyl-derivatives reduce LPS-induced inflammatory response in murine peritoneal macrophages via JAK-STAT and MAPKs signaling pathways. J. Funct. Foods 2019, 58, 95–104. [Google Scholar] [CrossRef]
- Torić, J.; Karković Marković, A.; Jakobušić Brala, C.; Barbarić, M. Anticancer effects of olive oil polyphenols and their combinations with anticancer drugs. Acta Pharm. 2019, 69, 461–482. [Google Scholar] [CrossRef] [Green Version]
- Potočnjak, I.; Škoda, M.; Pernjak-Pugel, E.; Peršić, M.P.; Domitrović, R. Oral administration of oleuropein attenuates cisplatin-induced acute renal injury in mice through inhibition of ERK signaling. Mol. Nutr. Food Res. 2016, 60, 530–541. [Google Scholar] [CrossRef] [PubMed]




| In Vivo/In Vitro | Type of Cancer/Cell | Antioxidant Properties of Ole | References |
|---|---|---|---|
| MCF-7 in vitro | Breast cancer cells |
| [25] |
| SH-SY5Y in vitro | Neuroblastoma cancer cells |
| [68] |
| HepG2 in vitro | Hepatocarcinoma cells |
| [15] |
| BPH-1 in vitro | Prostate cells |
| [52] |
| DU145 in vitro | Prostate cells |
| [52] |
| TPC-1 and BCPAP in vitro | Thyroid cancer cell |
| [49] |
| U87 in vitro | Human glioblastoma cells |
| [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rishmawi, S.; Haddad, F.; Dokmak, G.; Karaman, R. A Comprehensive Review on the Anti-Cancer Effects of Oleuropein. Life 2022, 12, 1140. https://doi.org/10.3390/life12081140
Rishmawi S, Haddad F, Dokmak G, Karaman R. A Comprehensive Review on the Anti-Cancer Effects of Oleuropein. Life. 2022; 12(8):1140. https://doi.org/10.3390/life12081140
Chicago/Turabian StyleRishmawi, Sabreen, Fatma Haddad, Ghadeer Dokmak, and Rafik Karaman. 2022. "A Comprehensive Review on the Anti-Cancer Effects of Oleuropein" Life 12, no. 8: 1140. https://doi.org/10.3390/life12081140
APA StyleRishmawi, S., Haddad, F., Dokmak, G., & Karaman, R. (2022). A Comprehensive Review on the Anti-Cancer Effects of Oleuropein. Life, 12(8), 1140. https://doi.org/10.3390/life12081140