
Sympathetic Reinnervation of Intact and Upper Follicle Xenografts into BALB/c-nu/nu Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Xenografts of Intact and Upper Follicles Can Enter Anagens
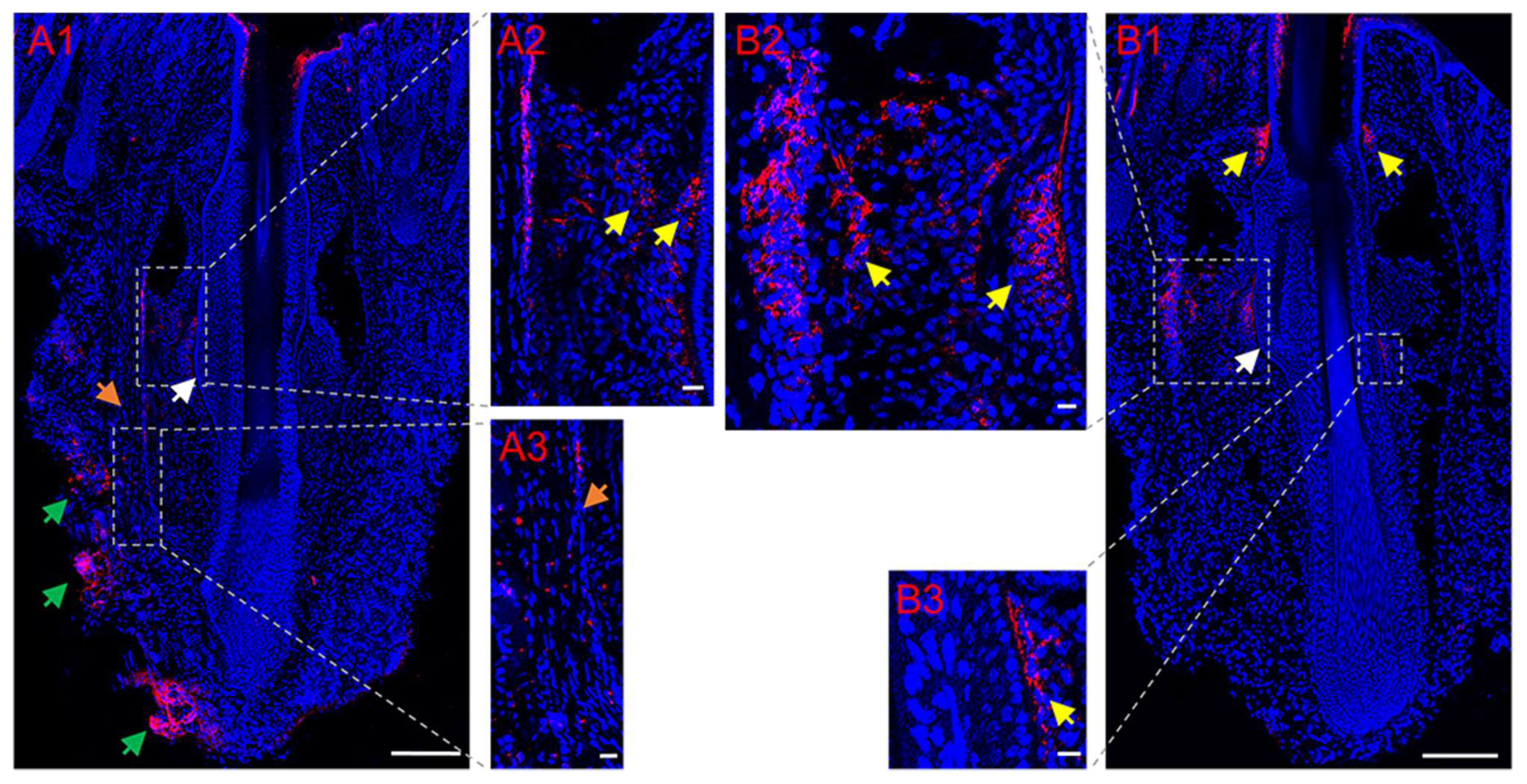
3.2. Partial Restoration of Sympathetic Reinnervation of the Transplanted Follicles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, S.; Goyal, I.; Mahendra, A. Quality of Life Assessment in Patients with Androgenetic Alopecia. Int. J. Trichol. 2019, 11, 147–152. [Google Scholar] [CrossRef]
- Rose, P.T. Hair restoration surgery: Challenges and solutions. Clin. Cosmet. Investig. Dermatol. 2015, 8, 361–370. [Google Scholar] [CrossRef]
- Betriu, N.; Jarrosson-Moral, C.; Semino, C.E. Culture and Differentiation of Human Hair Follicle Dermal Papilla Cells in a Soft 3D Self-Assembling Peptide Scaffold. Biomolecules 2020, 10, 684. [Google Scholar] [CrossRef]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, M.; Oliva, A.K.; Ke, M.S.; Ferdousi, F.; Isoda, H. 3D Spheroid Human Dermal Papilla Cell as an Effective Model for the Screening of Hair Growth Promoting Compounds: Examples of Minoxidil and 3,4,5-Tri-O-caffeoylquinic acid (TCQA). Cells 2022, 11, 2093. [Google Scholar] [CrossRef]
- Horne, K.A.; Jahoda, C.A. Restoration of hair growth by surgical implantation of follicular dermal sheath. Development 1992, 116, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Kazama, T.; Ito, M.; Urano, K.; Katakai, Y.; Yamaguchi, N.; Ueyama, Y. Histologic study of the regeneration process of human hair follicles grafted onto SCID mice after bulb amputation. J. Investig. Dermatol. Symp. Proc. 2001, 6, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.P.; Gong, Y.; Zhong, H.B.; Hua Wang, T.; Liao, X.H. EGFL6 expression in hair follicle central isthmus is dependent on the presence of terminal Schwann cells. Exp. Dermatol. 2020, 29, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Shwartz, Y.; Gonzalez-Celeiro, M.; Chen, C.L.; Pasolli, H.A.; Sheu, S.H.; Fan, S.M.; Shamsi, F.; Assaad, S.; Lin, E.T.; Zhang, B.; et al. Cell Types Promoting Goosebumps Form a Niche to Regulate Hair Follicle Stem Cells. Cell 2020, 182, 578–593. [Google Scholar] [CrossRef]
- Zhong, H.B.; Chu, Q.S.; Xiang, J.J.; Zhang, A.L.; Chen, E.Q.; Shen, X.R.; Liao, X.H. Spatial association of SEMA3C with nerve endings/terminal Schwann cells in hair follicle isthmus region. Int. J. Dev. Neurosci. 2020, 80, 737–741. [Google Scholar] [CrossRef]
- Fan, S.M.; Chang, Y.T.; Chen, C.L.; Wang, W.H.; Pan, M.K.; Chen, W.P.; Huang, W.Y.; Xu, Z.; Huang, H.E.; Chen, T.; et al. External light activates hair follicle stem cells through eyes via an ipRGC-SCN-sympathetic neural pathway. Proc. Natl. Acad. Sci. USA 2018, 115, E6880–E6889. [Google Scholar] [CrossRef] [PubMed]
- Crowe, R.; Mitsou, J.; McGrouther, D.A.; Burnstock, G. An increase in the growth of hair associated with hyperinnervation of the underlying vessels in rabbit skin. Neurosci. Lett. 1993, 161, 105–108. [Google Scholar] [CrossRef]
- Miranda, M.; Avila, I.; Esparza, J.; Shwartz, Y.; Hsu, Y.C.; Berdeaux, R.; Lowry, W.E. Defining a Role for G-Protein Coupled Receptor/cAMP/CRE-Binding Protein Signaling in Hair Follicle Stem Cell Activation. J. Investig. Dermatol. 2022, 142, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Borden, P.; Houtz, J.; Leach, S.D.; Kuruvilla, R. Sympathetic innervation during development is necessary for pancreatic islet architecture and functional maturation. Cell Rep. 2013, 4, 287–301. [Google Scholar] [CrossRef]
- Katayama, Y.; Battista, M.; Kao, W.M.; Hidalgo, A.; Peired, A.J.; Thomas, S.A.; Frenette, P.S. Signals from the sympathetic nervous system regulate hematopoietic stem cell egress from bone marrow. Cell 2006, 124, 407–421. [Google Scholar] [CrossRef]
- Sugitani, A.; Reynolds, J.C.; Tsuboi, M.; Todo, S. Extrinsic intestinal reinnervation after canine small bowel autotransplantation. Surgery 1998, 123, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.; Lagercrantz, H.; Zimmerman, H. Chemical Neurotransmission—An Introduction. In Neurotransm Vesicles; Academic Press: New York, NY, USA, 1986; pp. 16–21. [Google Scholar]
- Hall, W.D. An Overview of the Autonomic Nervous System. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990. [Google Scholar]
- Msheik, Z.; El Massry, M.; Rovini, A.; Billet, F.; Desmoulière, A. The macrophage: A key player in the pathophysiology of peripheral neuropathies. J. Neuroinflamm. 2022, 19, 97. [Google Scholar] [CrossRef]
- Coradazzi, M.; Gulino, R.; Garozzo, S.; Leanza, G. Selective lesion of the developing central noradrenergic system: Short- and long-term effects and reinnervation by noradrenergic-rich tissue grafts. J. Neurochem. 2010, 114, 761–771. [Google Scholar] [CrossRef]
- Awad, M.; Czer, L.S.; Hou, M.; Golshani, S.S.; Goltche, M.; De Robertis, M.; Kittleson, M.; Patel, J.; Azarbal, B.; Kransdorf, E.; et al. Early Denervation and Later Reinnervation of the Heart Following Cardiac Transplantation: A Review. J. Am. Heart Assoc. 2016, 5, e004070. [Google Scholar] [CrossRef]
- Persson-Sjögren, S.; Forsgren, S.; Täljedal, I.B. Peptides and other neuronal markers in transplanted pancreatic islets. Peptides 2000, 21, 741–752. [Google Scholar] [CrossRef]
- Mysore, V.; Kumaresan, M.; Garg, A.; Dua, A.; Venkatram, A.; Dua, K.; Singh, M.; Madura, C.; Chandran, R.; Rajput, R.S.; et al. Hair Transplant Practice Guidelines. J. Cutan. Aesthet. Surg. 2021, 14, 265–284. [Google Scholar]
- Sato, A.; Toyoshima, K.E.; Toki, H.; Ishibashi, N.; Asakawa, K.; Iwadate, A.; Kanayama, T.; Tobe, H.; Takeda, A.; Tsuji, T. Single follicular unit transplantation reconstructs arrector pili muscle and nerve connections and restores functional hair follicle piloerection. J. Dermatol. 2012, 39, 682–687. [Google Scholar] [CrossRef]
- Oliver, R.F. Regeneration of dermal papillae in rat vibrissae. J. Investig. Dermatol. 1966, 47, 496–497. [Google Scholar] [CrossRef]
- Kobayashi, K.; Nishimura, E. Ectopic growth of mouse whiskers from implanted lengths of plucked vibrissa follicles. J. Investig. Dermatol. 1989, 92, 278–282. [Google Scholar] [CrossRef]
- Yuan, Y.P.; Huang, K.; Xu, Y.M.; Chen, X.C.; Li, H.H.; Cai, B.Z.; Liu, Y.; Zhang, H.; Li, Y.; Lin, C.M. Canonical and non-canonical Wnt signaling control the regeneration of amputated rodent vibrissae follicles. J. Mol. Histol. 2016, 47, 1–8. [Google Scholar] [CrossRef]
- Iida, M.; Ihara, S.; Matsuzaki, T. Follicular epithelia and dermal papillae of mouse vibrissal follicles qualitatively change their hair-forming ability during anagen. Differentiation 2007, 75, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Müller-Röver, S.; Handjiski, B.; van der Veen, C.; Eichmüller, S.; Foitzik, K.; McKay, I.A.; Stenn, K.S.; Paus, R. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Investig. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef]
- Lam, S.M. Hair transplant and local anesthetics. Clin. Plast. Surg. 2013, 40, 615–625. [Google Scholar] [CrossRef]
- Kite, A.; Lucas, V.S. Hair transplant: A basic review. Plast. Surg. Nurs. 2015, 35, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Haslam, I.S.; Sharov, A.A.; Botchkarev, V.A. Pathobiology of chemotherapy-induced hair loss. Lancet Oncol. 2013, 14, e50–e59. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Kazama, T.; Ito, M.; Urano, K.; Katakai, Y.; Yamaguchi, N.; Ueyama, Y. Histologic and cell kinetic studies of hair loss and subsequent recovery process of human scalp hair follicles grafted onto severe combined immunodeficient mice. J. Investig. Dermatol. 2000, 115, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Geary, J.R., Jr. Effect of roentgen rays during various phases of the hair cycle of the albino rat. Am. J. Anat. 1952, 91, 51–105. [Google Scholar] [CrossRef] [PubMed]
- Wolbach, S.B. The hair cycle of the mouse and its importance in the study of sequences of experimental carcinogenesis. Ann. N. Y. Acad. Sci. 1951, 53, 517–536. [Google Scholar] [CrossRef] [PubMed]
- Crounse, R.G.; Stengle, J.M. Influence of the dermal papilla on survival of isolated human scalp hair roots in an heterologous host. J. Investig. Dermatol. 1959, 32, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Rompolas, P.; Deschene, E.R.; Zito, G.; Gonzalez, D.G.; Saotome, I.; Haberman, A.M.; Greco, V. Live imaging of stem cell and progeny behaviour in physiological hair-follicle regeneration. Nature 2012, 487, 496–499. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, T. Local and systemic mechanisms that control the hair follicle stem cell niche. Nat. Rev. Mol. Cell Biol. 2023. [Google Scholar] [CrossRef]
- Edmunds, L.H., Jr.; Nadel, J.A.; Graf, P.D. Reinnervation of the reimplanted canine lung. J. Appl. Physiol. 1971, 31, 722–727. [Google Scholar] [CrossRef]
- Korsgren, O.; Jansson, L.; Andersson, A.; Sundler, F. Reinnervation of transplanted pancreatic islets. A comparison among islets implanted into the kidney, spleen, and liver. Transplantation 1993, 56, 138–143. [Google Scholar] [CrossRef]
- Myrsén, U.; Keymeulen, B.; Pipeleers, D.G.; Sundler, F. Beta cells are important for islet innervation: Evidence from purified rat islet-cell grafts. Diabetologia 1996, 39, 54–59. [Google Scholar] [CrossRef]
- Penttilä, H.; Huttunen, P.; von Smitten, K.; Ashammakhi, N.; Waris, T. Reinnervation of arterial grafts by adrenergic nerves occurs in rats as indicated by increased levels of noradrenaline. Plast. Reconstr. Surg. 2004, 113, 2057–2060. [Google Scholar] [CrossRef]
- Grisk, O.; Rose, H.J.; Rettig, R. The development of post-transplantation hypertension in recipients of an SHR kidney is independent of reinnervation of the graft. Pflug. Arch. 1999, 438, 502–507. [Google Scholar] [CrossRef]
- Grisk, O.; Gröne, H.J.; Rose, H.J.; Rettig, R. Sympathetic reinnervation of rat kidney grafts. Transplantation 2001, 72, 1153–1155. [Google Scholar] [CrossRef]
- Guimarães, G.V.; D’Avila, V.; Bocchi, E.A.; Carvalho, V.O. Norepinephrine remains increased in the six-minute walking test after heart transplantation. Clinics 2010, 65, 587–591. [Google Scholar] [CrossRef]
- Bengel, F.M.; Ueberfuhr, P.; Hesse, T.; Schiepel, N.; Ziegler, S.I.; Scholz, S.; Nekolla, S.G.; Reichart, B.; Schwaiger, M. Clinical determinants of ventricular sympathetic reinnervation after orthotopic heart transplantation. Circulation 2002, 106, 831–835. [Google Scholar] [CrossRef]
- Olson, L.; Malmfors, T. Growth characteristics of adrenergic nerves in the adult rat. Fluorescence histochemical and 3H-noradrenaline uptake studies using tissue transplantations to the anterior chamber of the eye. Acta Physiol. Scand. Suppl. 1970, 348, 1–112. [Google Scholar] [PubMed]
- Buckley, G.; Wong, J.; Metcalfe, A.D.; Ferguson, M.W. Denervation affects regenerative responses in MRL/MpJ and repair in C57BL/6 ear wounds. J. Anat. 2012, 220, 3–12. [Google Scholar] [CrossRef]
- Pius-Sadowska, E.; Machaliński, B. Pleiotropic activity of nerve growth factor in regulating cardiac functions and counteracting pathogenesis. ESC Heart Fail. 2021, 8, 974–987. [Google Scholar] [CrossRef] [PubMed]
- Mattson, E.E.; Marshall, C.D. Follicle Microstructure and Innervation Vary between Pinniped Micro- and Macrovibrissae. Brain Behav. Evol. 2016, 88, 43–58. [Google Scholar] [CrossRef]
- Drake, S.E.; Crish, S.D.; George, J.C.; Stimmelmayr, R.; Thewissen, J.G. Sensory Hairs in the Bowhead Whale, Balaena mysticetus (Cetacea, Mammalia). Anat. Rec. 2015, 298, 1327–1335. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.-W.; Ni, N.; Xie, X.-J.; Zhao, Y.-L.; Liang, W.-Z.; Huang, Y.-X.; Lin, C.-M. Sympathetic Reinnervation of Intact and Upper Follicle Xenografts into BALB/c-nu/nu Mice. Life 2023, 13, 2163. https://doi.org/10.3390/life13112163
Chen X-W, Ni N, Xie X-J, Zhao Y-L, Liang W-Z, Huang Y-X, Lin C-M. Sympathetic Reinnervation of Intact and Upper Follicle Xenografts into BALB/c-nu/nu Mice. Life. 2023; 13(11):2163. https://doi.org/10.3390/life13112163
Chicago/Turabian StyleChen, Xiu-Wen, Na Ni, Xiao-Jun Xie, Ying-Lin Zhao, Wen-Zi Liang, Yu-Xin Huang, and Chang-Min Lin. 2023. "Sympathetic Reinnervation of Intact and Upper Follicle Xenografts into BALB/c-nu/nu Mice" Life 13, no. 11: 2163. https://doi.org/10.3390/life13112163
APA StyleChen, X. -W., Ni, N., Xie, X. -J., Zhao, Y. -L., Liang, W. -Z., Huang, Y. -X., & Lin, C. -M. (2023). Sympathetic Reinnervation of Intact and Upper Follicle Xenografts into BALB/c-nu/nu Mice. Life, 13(11), 2163. https://doi.org/10.3390/life13112163