

Molecular Characterization of Influenza A/H3N2 Virus Isolated from Indonesian Hajj and Umrah Pilgrims 2013 to 2014

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Population Study and Sampling Strategy
2.3. RNA Isolation and Confirmatory Real-Time RT-PCR for MERS-CoV and Influenza Virus
2.4. Direct Sequencing of Influenza A/H3N2 Virus
2.5. Phylogenetic Analysis of Influenza A/H3N2 Virus
3. Results
3.1. MERS-CoV and Influenza Virus Confirmatory
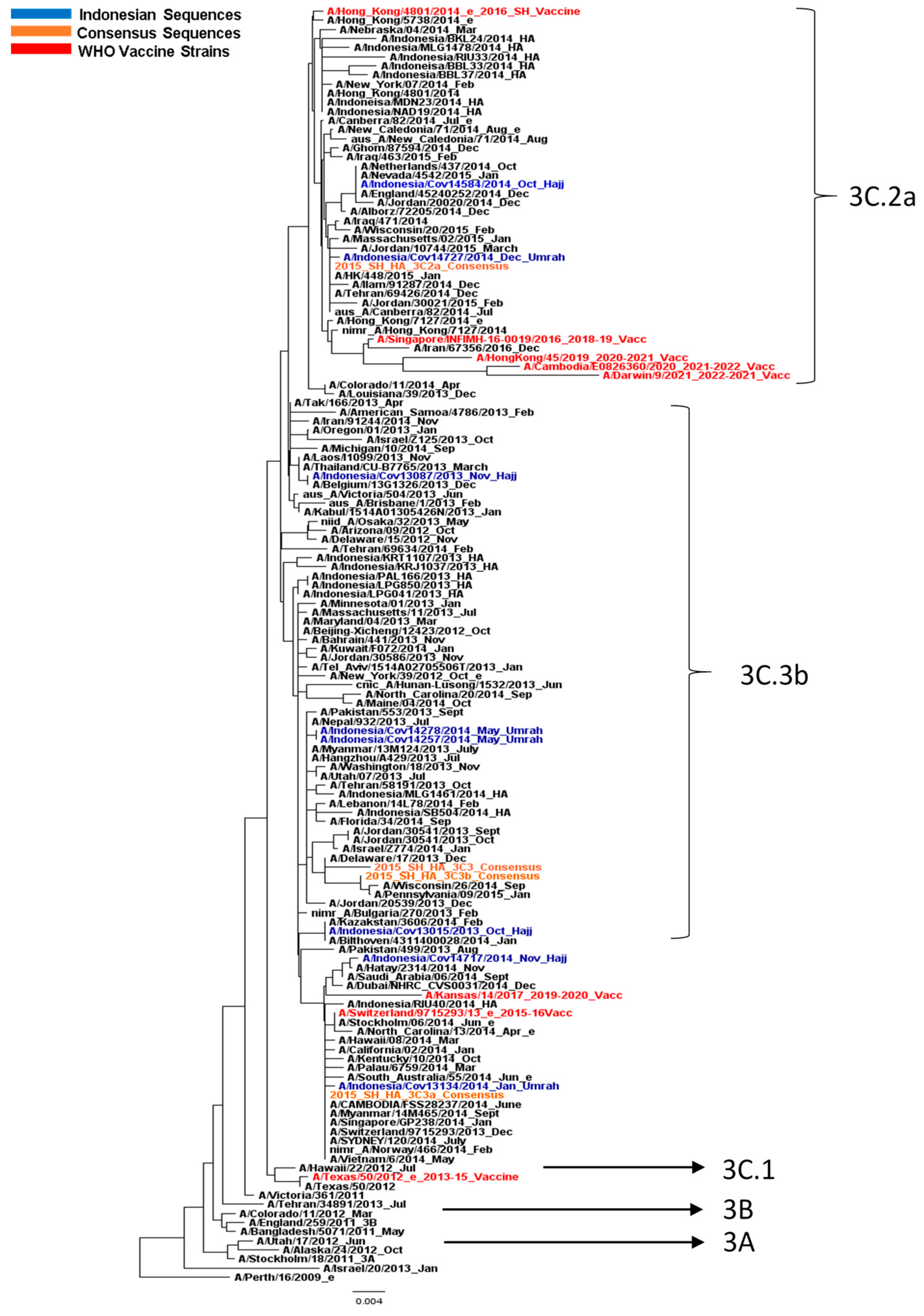
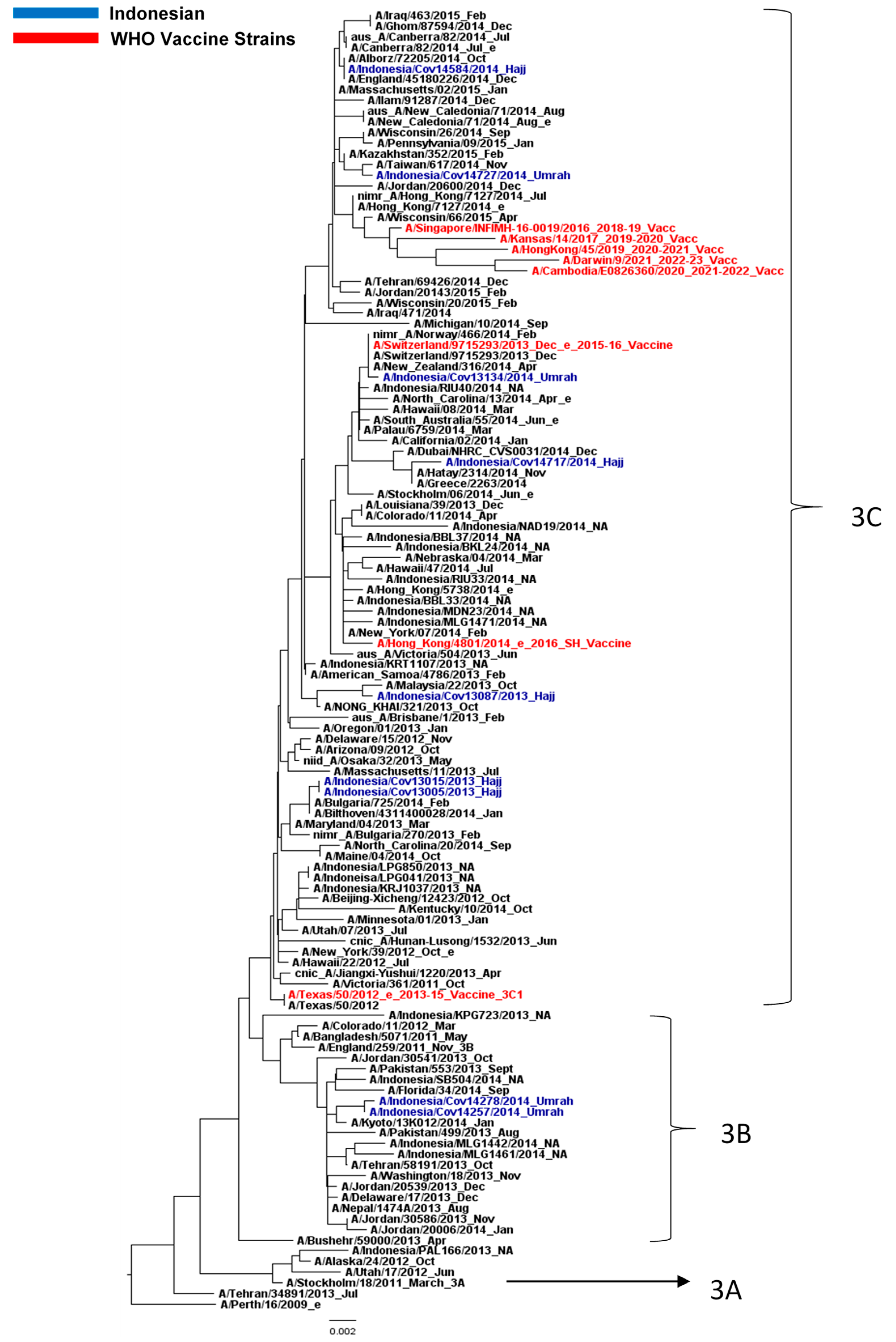
3.2. Influenza A/H3N2 Virus Analysis
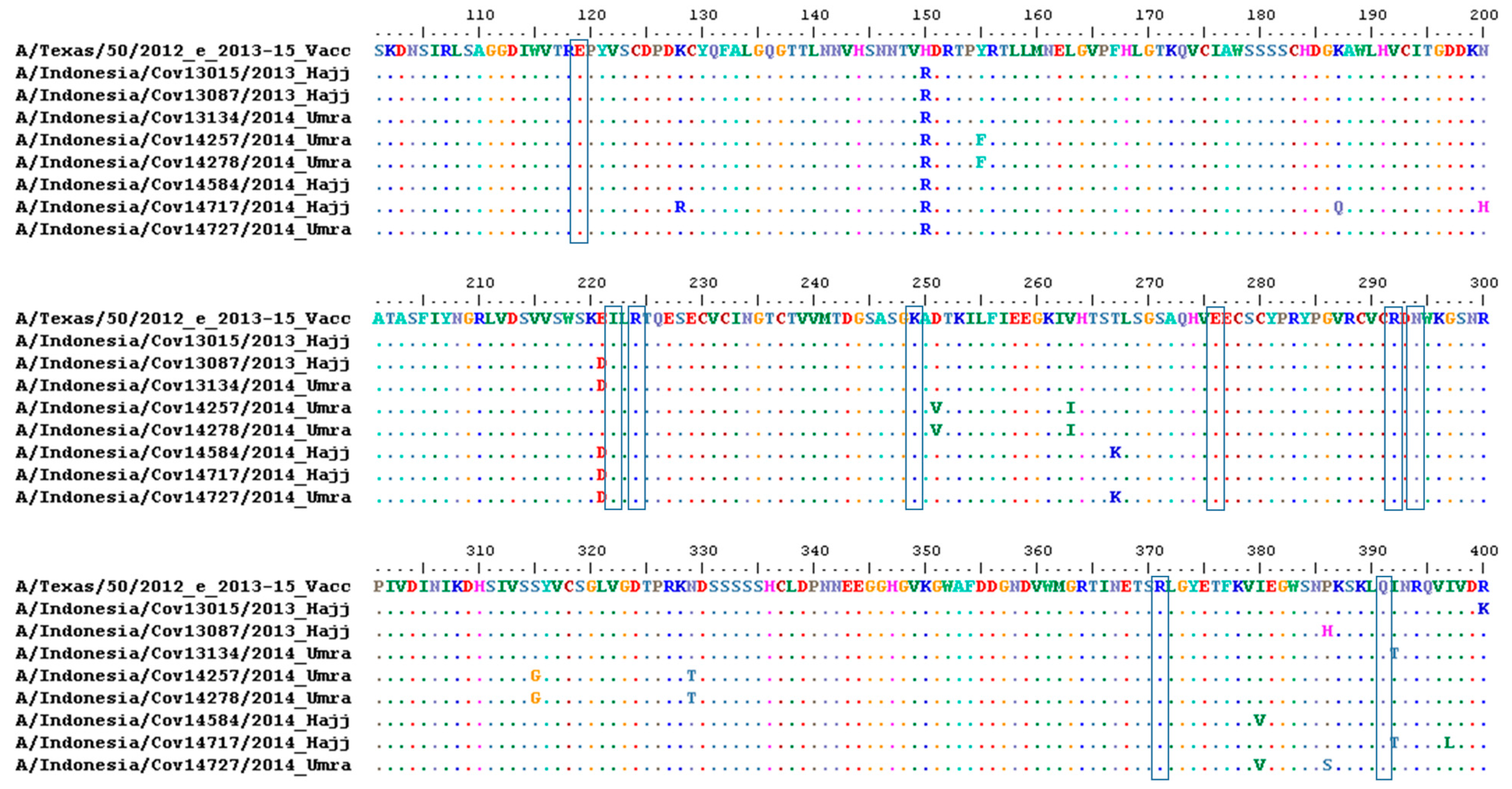
3.3. Amino Acid Analysis of HA and NA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Memish, Z.A.; Zumla, A.; Alhakeem, R.F.; Assiri, A.; Turkestani, A.; Al Harby, K.D.; Alyemni, M.; Dhafar, K.; Gautret, P.; Barbeschi, M.; et al. Hajj: Infectious disease surveillance and control. Lancet 2014, 383, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Benkouiten, S.; Charrel, R.; Belhouchat, K.; Drali, T.; Salez, N.; Nougairede, A.; Zandotti, C.; Memish, Z.A.; al Masri, M.; Gaillard, C.; et al. Circulation of respiratory viruses among pilgrims during the 2012 Hajj pilgrimage. Clin. Infect. Dis. 2013, 57, 992–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautret, P.; Benkouiten, S.; Al-Tawfiq, J.A.; Memish, Z.A. The spectrum of respiratory pathogens among returning Hajj pilgrims: Myths and reality. Int. J. Infect. Dis. 2016, 47, 83–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, A.M.; Al-Subhi, T.L.; Badroon, N.A.; Hassan, A.M.; Bajrai, L.H.M.; Banassir, T.M.; Alquthami, K.M.; Azhar, E.I. MERS-CoV, influenza and other respiratory viruses among symptomatic pilgrims during 2014 Hajj season. J. Med. Virol. 2019, 91, 911–917. [Google Scholar] [CrossRef]
- WHO. WHO Statement on the third meeting of the IHR Emergency committee concerning Middle East respiratory syndrome coronavirus (MERS-CoV). Wkly. Epidemiol. Rec. 2013, 88, 435–436. [Google Scholar]
- WHO. MERS Situation Update, January 2023. Available online: http://www.emro.who.int/health-topics/mers-cov/mers-outbreaks.html (accessed on 1 March 2023).
- Corman, V.M.; Eckerle, I.; Bleicker, T.; Zaki, A.; Landt, O.; Eschbach-Bludau, M.; van Boheemen, S.; Gopal, R.; Ballhause, M.; Bestebroer, T.M.; et al. Detection of a novel human coronavirus by real-time reverse-transcription polymerase chain reaction. Euro Surveill. 2012, 17, 20285. [Google Scholar] [CrossRef] [Green Version]
- Aberle, J.H.; Popow-Kraupp, T.; Kreidl, P.; Laferl, H.; Heinz, F.X.; Aberle, S.W. Influenza A and B Viruses but Not MERS-CoV in Hajj Pilgrims, Austria, 2014. Emerg. Infect. Dis. 2015, 21, 726–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Liu, F.; Liu, L.; Zhang, L.; Lu, M.; Abudukadeer, A.; Wang, L.; Tian, F.; Zhen, W.; Yang, P.; et al. No MERS-CoV but positive influenza viruses in returning Hajj pilgrims, China, 2013–2015. BMC Infect. Dis. 2017, 17, 715. [Google Scholar] [CrossRef] [Green Version]
- Memish, Z.A.; Almasri, M.; Turkestani, A.; Al-Shangiti, A.M.; Yezli, S. Etiology of severe community-acquired pneumonia during the 2013 Hajj-part of the MERS-CoV surveillance program. Int. J. Infect. Dis. 2014, 25, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2018, 16, 47–60. [Google Scholar] [CrossRef]
- WHO-SEARO. Seasonal influenza surveillance (2009–2017) for pandemic preparedness in the WHO South-East Asia Region. WHO South East Asia J. Public Health 2020, 9, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.D.; Ross, T.M. H3N2 influenza viruses in humans: Viral mechanisms, evolution, and evaluation. Hum. Vaccines Immunother. 2018, 14, 1840–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaji, M.; Watanabe, A.; Aizawa, H. Differences in clinical features between influenza A H1N1, A H3N2, and B in adult patients. Respirology 2003, 8, 231–233. [Google Scholar] [CrossRef] [PubMed]
- Agustiningsih, A.; Trimarsanto, H.; Restuadi, R.; Artika, I.M.; Hellard, M.; Muljono, D.H. Evolutionary study and phylodynamic pattern of human influenza A/H3N2 virus in Indonesia from 2008 to 2010. PLoS ONE 2018, 13, e0201427. [Google Scholar] [CrossRef]
- Mossad, S.B. Influenza update 2018-2019: 100 years after the great pandemic. Clevel. Clin. J. Med. 2018, 85, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.W.; Li, Z.J.; Hu, W.B. Global dynamic spatiotemporal pattern of seasonal influenza since 2009 influenza pandemic. Infect. Dis. Poverty 2020, 9, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corman, V.M.; Muller, M.A.; Costabel, U.; Timm, J.; Binger, T.; Meyer, B.; Kreher, P.; Lattwein, E.; Eschbach-Bludau, M.; Nitsche, A.; et al. Assays for laboratory confirmation of novel human coronavirus (hCoV-EMC) infections. Euro Surveill. 2012, 17, 20334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Laboratory Testing for Middle East Respiratory Syndrome Coronavirus. Interim recommendations: September 2013. Available online: https://www.who.int/publications/i/item/10665-259952 (accessed on 15 February 2022).
- WHO. WHO Global Influenza Surveillance Network: Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; WHO: Geneva, Switzerland, 2011.
- Agustiningsih, A.; Trimarsanto, H.; Setiawaty, V.; Artika, I.M.; Muljono, D.H. Primer development to obtain complete coding sequence of HA and NA genes of influenza A/H3N2 virus. BMC Res. Notes 2016, 9, 423. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Chambers, B.S.; Parkhouse, K.; Ross, T.M.; Alby, K.; Hensley, S.E. Identification of Hemagglutinin Residues Responsible for H3N2 Antigenic Drift during the 2014–2015 Influenza Season. Cell Rep. 2015, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Koel, B.F.; Burke, D.F.; Bestebroer, T.M.; van der Vliet, S.; Zondag, G.C.; Vervaet, G.; Skepner, E.; Lewis, N.S.; Spronken, M.I.; Russell, C.A.; et al. Substitutions near the receptor binding site determine major antigenic change during influenza virus evolution. Science 2013, 342, 976–979. [Google Scholar] [CrossRef] [PubMed]
- Tewawong, N.; Prachayangprecha, S.; Vichiwattana, P.; Korkong, S.; Klinfueng, S.; Vongpunsawad, S.; Thongmee, T.; Theamboonlers, A.; Poovorawan, Y. Assessing Antigenic Drift of Seasonal Influenza A(H3N2) and A(H1N1)pdm09 Viruses. PLoS ONE 2015, 10, e0139958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaraket, H.; Saito, R.; Sato, I.; Suzuki, Y.; Li, D.; Dapat, C.; Caperig-Dapat, I.; Oguma, T.; Sasaki, A.; Suzuki, H. Molecular evolution of human influenza A viruses in a local area during eight influenza epidemics from 2000 to 2007. Arch. Virol. 2009, 154, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Earl, D.J.; Deem, M.W. Quantifying influenza vaccine efficacy and antigenic distance. Vaccine 2006, 24, 3881–3888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samson, M.; Pizzorno, A.; Abed, Y.; Boivin, G. Influenza virus resistance to neuraminidase inhibitors. Antivir. Res. 2013, 98, 174–185. [Google Scholar] [CrossRef] [PubMed]
- WHO. Summary of Neuraminidase (NA) Amino Acid Substitutions Associated with Reduced Inhibition by Neuraminidase Inhibitors (NAIs). Available online: https://www.who.int/publications/m/item/summary-of-neuraminidase-(na)-amino-acid-substitutions-associated-with-reduced-inhibition-by-neuraminidase-inhibitors-(nais) (accessed on 1 March 2023).
- Kementerian Agama Republik Indonesia Direktorat Jenderal Penyelenggaraan Haji dan Umrah. Haji Dalam Angka: Jumlah Jemaah Haji Indonesia Dalam Seabad Lebih. Available online: https://haji.kemenag.go.id/v4/haji-dalam-angka-jumlah-jemaah-haji-indonesia-dalam-seabad-lebih (accessed on 12 December 2022).
- Gatrad, A.R.; Sheikh, A. Hajj: Journey of a lifetime. BMJ 2005, 330, 133–137. [Google Scholar] [CrossRef]
- Kementerian Kesehatan Republik Indonesia Pusat Kesehatan Haji. Laporan Evaluasi Penyelenggaraan Kesehatan Haji Tahun 2017; Kementerian Kesehatan Republik Indonesia Pusat Kesehatan Haji: Jakarta, Indonesia, 2017.
- Pane, M.; Imari, S.; Alwi, Q.; Nyoman Kandun, I.; Cook, A.R.; Samaan, G. Causes of mortality for Indonesian Hajj Pilgrims: Comparison between routine death certificate and verbal autopsy findings. PLoS ONE 2013, 8, e73243. [Google Scholar] [CrossRef]
- Amin, M.; Bakhtiar, A.; Subarjo, M.; Aksono, E.B.; Widiyanti, P.; Shimizu, K.; Mori, Y. Screening for Middle East respiratory syndrome coronavirus among febrile Indonesian Hajj pilgrims: A study on 28,197 returning pilgrims. J. Infect. Prev. 2018, 19, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Gautret, P.; Charrel, R.; Benkouiten, S.; Belhouchat, K.; Nougairede, A.; Drali, T.; Salez, N.; Memish, Z.A.; Al Masri, M.; Lagier, J.C.; et al. Lack of MERS coronavirus but prevalence of influenza virus in French pilgrims after 2013 Hajj. Emerg. Infect. Dis. 2014, 20, 728–730. [Google Scholar] [CrossRef]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [Green Version]
- Tamerius, J.; Nelson, M.I.; Zhou, S.Z.; Viboud, C.; Miller, M.A.; Alonso, W.J. Global influenza seasonality: Reconciling patterns across temperate and tropical regions. Environ. Health Perspect. 2011, 119, 439–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colizza, V.; Barrat, A.; Barthelemy, M.; Vespignani, A. The role of the airline transportation network in the prediction and predictability of global epidemics. Proc. Natl. Acad. Sci. USA 2006, 103, 2015–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevilla, N.L. Germs on a Plane: The Transmission and Risks of Airplane-Borne Diseases. Transp. Res. Rec. 2018, 2672, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfelali, M.; Khandaker, G.; Booy, R.; Rashid, H. Mismatching between circulating strains and vaccine strains of influenza: Effect on Hajj pilgrims from both hemispheres. Hum. Vaccines Immunother. 2016, 12, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Greenbaum, B.D.; Ghedin, E. Viral evolution: Beyond drift and shift. Curr. Opin. Microbiol. 2015, 26, 109–115. [Google Scholar] [CrossRef]
- Webster, R.G.; Govorkova, E.A. Continuing challenges in influenza. Ann. N. Y. Acad. Sci. 2014, 1323, 115–139. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Xu, H.; Guan, S.; Wang, C.; Dong, G. Frequency and distribution of H1N1 influenza A viruses with oseltamivir-resistant mutations worldwide before and after the 2009 pandemic. J. Med. Virol. 2022, 94, 4406–4416. [Google Scholar] [CrossRef]
- Koel, B.F.; Vigeveno, R.M.; Pater, M.; Koekkoek, S.M.; Han, A.X.; Tuan, H.M.; Anh, T.T.N.; Hung, N.T.; Thinh, L.Q.; Hai, L.T.; et al. Longitudinal sampling is required to maximize detection of intrahost A/H3N2 virus variants. Virus Evol. 2020, 6, veaa088. [Google Scholar] [CrossRef]
- Pawestri, H.A.; Nugraha, A.A.; Hariastuti, N.I.; Setiawaty, V. Detection of neuraminidase inhibitor-resistant influenza A (H1N1)pdm09 viruses obtained from influenza surveillance in Indonesia. SAGE Open Med. 2018, 6, 2050312118818293. [Google Scholar] [CrossRef] [Green Version]
- Bright, R.A.; Medina, M.J.; Xu, X.; Perez-Oronoz, G.; Wallis, T.R.; Davis, X.M.; Povinelli, L.; Cox, N.J.; Klimov, A.I. Incidence of adamantane resistance among influenza A (H3N2) viruses isolated worldwide from 1994 to 2005: A cause for concern. Lancet 2005, 366, 1175–1181. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Demography | Total | % | Influenza A | Influenza B | Influenza A and B | Negative | ||
|---|---|---|---|---|---|---|---|---|
| H1pdm | H3 | Non H1/H3/H5 | ||||||
| Sex | ||||||||
| Male | 126 | 50.2 | 29 | 5 | 1 | 15 | 1 | 75 |
| Female | 125 | 49.8 | 22 | 10 | 3 | 14 | 0 | 76 |
| Age (y.o.) | ||||||||
| ≤50 | 65 | 25.9 | 11 | 3 | 3 | 6 | 0 | 42 |
| >50 | 176 | 70.1 | 38 | 12 | 1 | 23 | 0 | 102 |
| No Data | 10 | 4.0 | 2 | 0 | 0 | 0 | 1 | 7 |
| Origin | ||||||||
| Sumatra | 67 | 26.7 | 14 | 3 | 0 | 13 | 1 | 36 |
| Java | 133 | 53.0 | 24 | 10 | 4 | 14 | 0 | 81 |
| Bali | 11 | 4.4 | 3 | 0 | 0 | 0 | 0 | 8 |
| Sulawesi | 3 | 1.2 | 1 | 0 | 0 | 1 | 0 | 1 |
| Kalimantan | 37 | 14.7 | 9 | 2 | 0 | 1 | 0 | 25 |
| AA Position * | AA 2013–2015 Vaccine Strain (A/Texas/50/2012) | AA Sample (n) | Association with Antigenicity |
|---|---|---|---|
| 3 | L | I (2) | no association [23,24] |
| 10 | T | M (2) | no association [23,24] |
| 128 | N | A (4); T (4) | antigenic B site [25] |
| 138 | A | S (2) | antigenic site A [25] |
| 142 | R | G (5) | antigenic site A [25] |
| 144 | N | S (2) | antigenic site A, RBD [23,24] |
| 145 | N | S (8) | antigenic site A, RBD [23] |
| 157 | L | S (2) | antigenic site B [25] |
| 159 | F | S (2); Y (2) | antigenic site B [23,25] |
| 160 | K | T (2) | antigenic site B [25] |
| 186 | V | G (8) | antigenic B, RBD [26] |
| 198 | P | S (8) | antigenic site B [25] |
| 219 | F | S (8) | antigenic site D [27] |
| 225 | N | D (4) | RBD [26] |
| 311 | G | H (2) | no association [24] |
| 347 | V | M (2) | no association [24] |
| 160 | K | T (2) | antigenic site B [25] |
| 186 | V | G (8) | antigenic B, RBD [26] |
| 198 | P | S (8) | antigenic site B [25] |
| 219 | F | S (8) | antigenic site D [27] |
| 489 | D | N (4) | no association [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agustiningsih, A.; Indalao, I.L.; Pangesti, K.A.; Sukowati, C.H.C.; Ramadhany, R. Molecular Characterization of Influenza A/H3N2 Virus Isolated from Indonesian Hajj and Umrah Pilgrims 2013 to 2014. Life 2023, 13, 1100. https://doi.org/10.3390/life13051100
Agustiningsih A, Indalao IL, Pangesti KA, Sukowati CHC, Ramadhany R. Molecular Characterization of Influenza A/H3N2 Virus Isolated from Indonesian Hajj and Umrah Pilgrims 2013 to 2014. Life. 2023; 13(5):1100. https://doi.org/10.3390/life13051100
Chicago/Turabian StyleAgustiningsih, Agustiningsih, Irene Lorinda Indalao, Krisnanur A. Pangesti, Caecilia H. C. Sukowati, and Ririn Ramadhany. 2023. "Molecular Characterization of Influenza A/H3N2 Virus Isolated from Indonesian Hajj and Umrah Pilgrims 2013 to 2014" Life 13, no. 5: 1100. https://doi.org/10.3390/life13051100
APA StyleAgustiningsih, A., Indalao, I. L., Pangesti, K. A., Sukowati, C. H. C., & Ramadhany, R. (2023). Molecular Characterization of Influenza A/H3N2 Virus Isolated from Indonesian Hajj and Umrah Pilgrims 2013 to 2014. Life, 13(5), 1100. https://doi.org/10.3390/life13051100