
Immunomodulation, Bioavailability and Safety of Bacteriocins


Abstract
:1. Introduction
2. Resources, Gene Organization and Biosynthesis of Bacteriocins
3. Mechanism of Action on Microorganisms
4. Immunomodulatory Activity of Bacteriocins

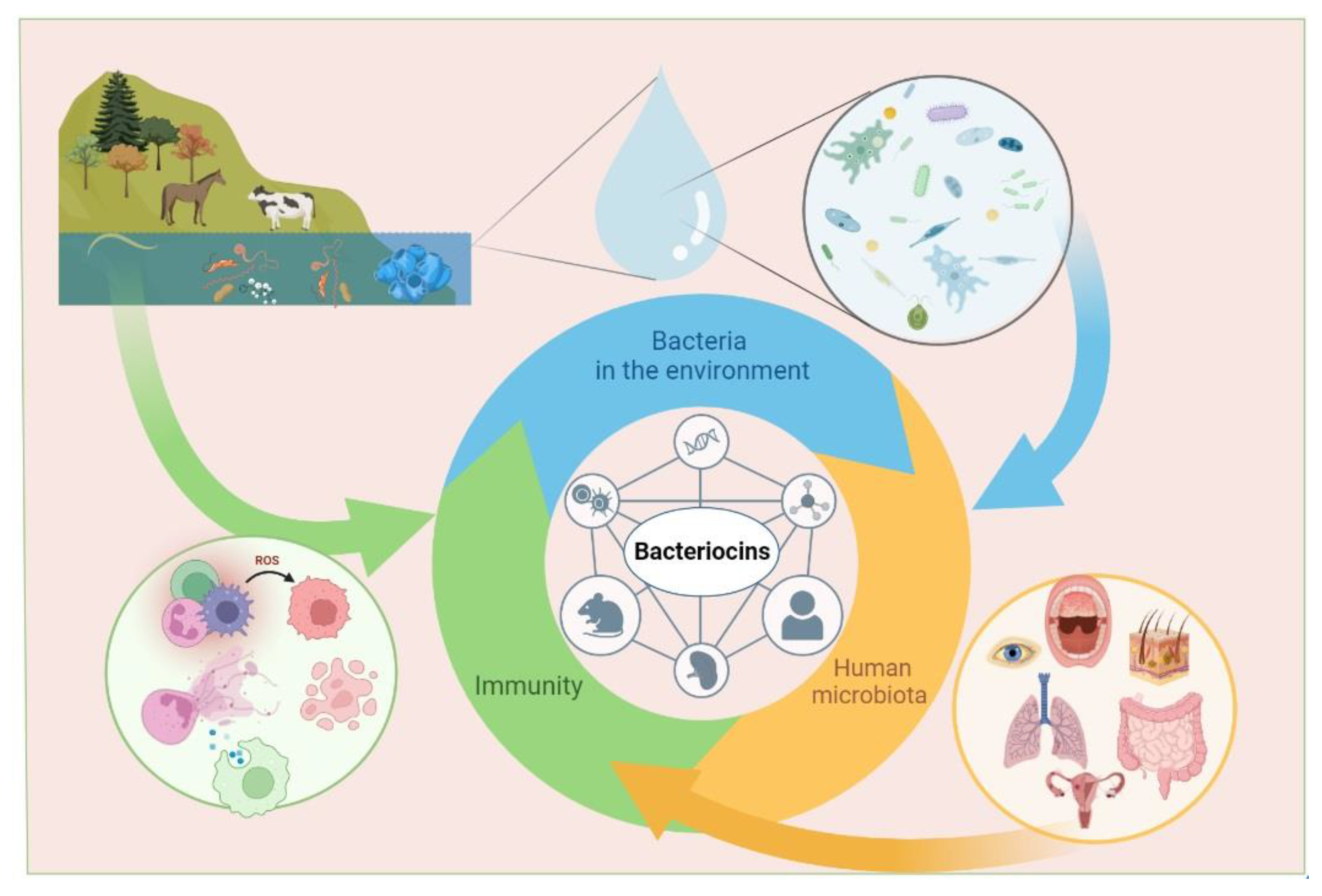{kind=link}
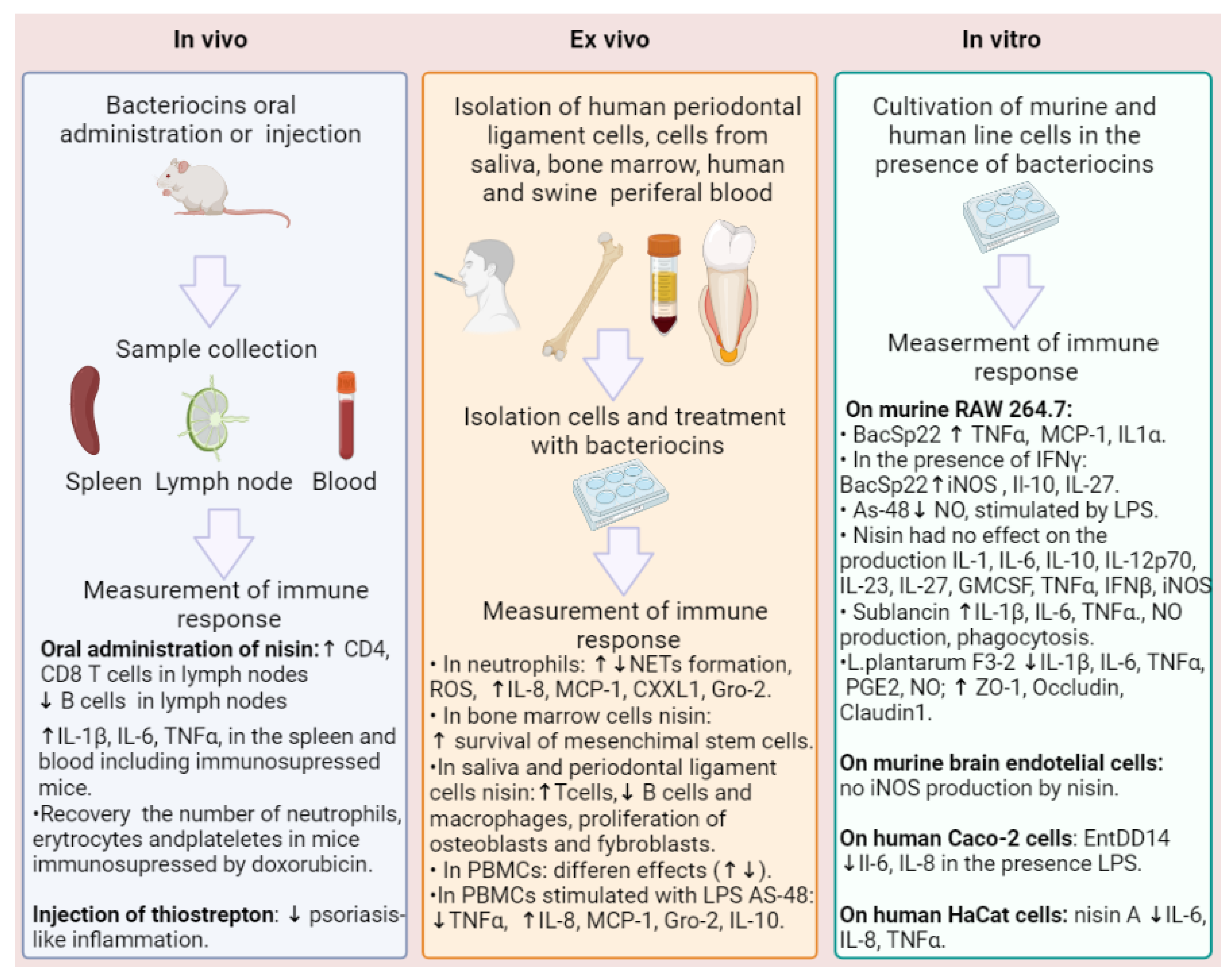{kind=link}
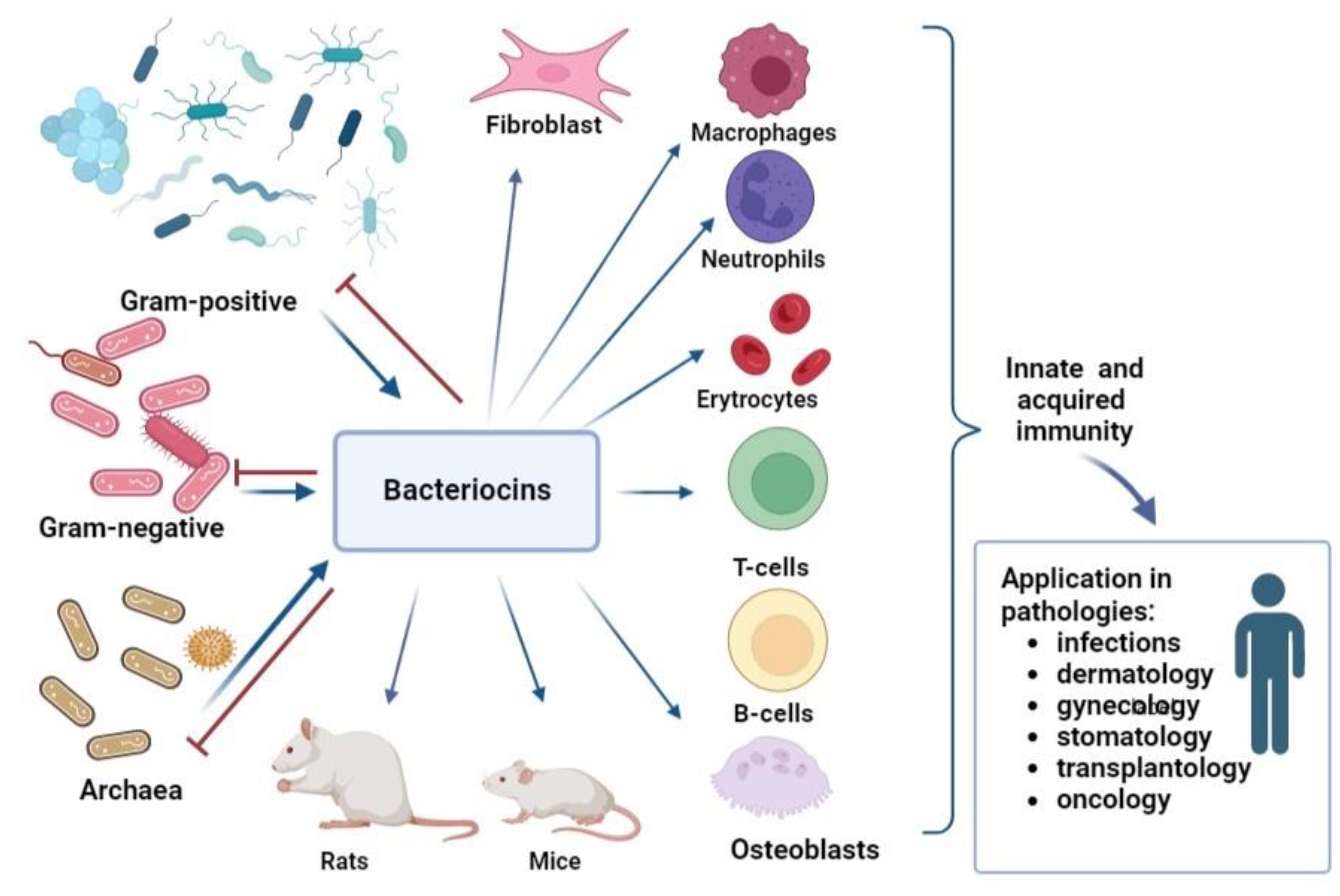{kind=link}
| Bacteriocin | Resource | Highlights | Ref. |
|---|---|---|---|
| Nisin Z | L. lactis | Inducing MCP-1, Gro-α, and IL-6 in human PBMC. In response to LPS, reducing TNF-α and inhibiting IL-6. | [99] |
| Nisin | L. lactis | Increasing T-cells, fibroblasts, and osteoblasts; increasing B cells. Increasing the expression of IL-10, FGF-2, and TGF-β genes and the synthesis of TGF-β and FGF-2. Decreasing phagocytosis and inhibiting the synthesis of IL-6. | [100,101,102,105,106,107] |
| Nisin A | L. lactis | No influence on IL-1α, IL-1β, IL-6, IL-10, IL-12p70, IL-17A, IL-23, IL-27, GMCSF, IFN-β, TNF-α, iNOS, NO production, and NETs. | [108] |
| Inducing NETs formation and ROX in human neutrophils. | [111] | ||
| BacSp222 | Staphylococcus pseudintermedius 222 | Increasing NO, iNOS in P388.D1, and RAW 264.7; increasing IL-10 and IL-27 in combination with IFN-γ. In human neutrophils, upregulating IL-8; absence of ROS production or NETs formation. | [108] |
| AS-48 | Enterococcus spp. | Decreasing NO production induced by LPS on RAW cells; absence of pro-inflammatory effects. | [110] |
| Sublancin | Bacillus subtilis 168 | Increasing CXCL1 and MCP-1; decreasing TNF-α in murine peritoneal macrophages and neutrophils. Increasing IL-1β, IL-6, TNF-alpha, nitric oxide, and phagocytosis. Increasing T-cells in the mesenteric lymph nodes. Increasing IL-1β, IL-6, and TNF-α in the spleen of immunosuppressed BALB/c mice. | [112,113,114] |
| L. plantarum F3-2 | Lactiplantibacillus plantarum | Reducing elevated levels of IL1β, IL6, TNFα, nitric oxide, and prostaglandin E2 induced by LPS in RAW264.7 cells. Increasing the expression levels of ZO-1, Occludin, and Claudin 1. | [115] |
| Acidocin A | Lactobacillus acidophilus TK9201 | Increasing IL-6, TNFα, MIG/CXCL9, MCP-1/CCL2, MCP-3/CCL7, and MIP-1β in PBMCs. | [116] |
| Ent DD14 | Enterococcus faecalis 14 | Decreasing IL-6 and IL-8. | [117] |
| Thiostrepton | Streptomyces genus | Increasing HspA1A, Hsp70, Hsp90α, or Hsp105. | [127] |
5. Bioavailability and Safety of Bacteriocins
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S.; Kering, K.; Wairimu, C.; Onsare, R.; Mbae, C. Antimicrobial Resistance Rates and Surveillance in Sub-Saharan Africa: Where Are We Now? Infect. Drug Resist. 2022, 15, 3589–3609. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Aqib, A.I.; Muzammil, I.; Majeed, N.; Bhutta, Z.A.; Kulyar, M.F.-E.; Fatima, M.; Zaheer, C.-N.F.; Muneer, A.; Murtaza, M.; et al. MRSA compendium of epidemiology, transmission, pathophysiology, treatment, and prevention within one health framework. Front. Microbiol. 2023, 13, 1067284. [Google Scholar] [CrossRef]
- WHO. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report. 2022. Available online: https://www.who.int/publications/i/item/9789240062702 (accessed on 19 February 2023).
- de Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, A.; Kurittu, P.; Al-Mustapha, A.I.; Heljanko, V.; Johansson, V.; Thakali, O.; Mishra, S.K.; Lehto, K.-M.; Lipponen, A.; Oikarinen, S.; et al. Wastewater surveillance of antibiotic-resistant bacterial pathogens: A systematic review. Front. Microbiol. 2022, 13, 977106. [Google Scholar] [CrossRef]
- Nguyen, A.Q.; Vu, H.P.; Nguyen, L.N.; Wang, Q.; Djordjevic, S.P.; Donner, E.; Yin, H.; Nghiem, L.D. Monitoring antibiotic resistance genes in wastewater treatment: Current strategies and future challenges. Sci. Total. Environ. 2021, 783, 146964. [Google Scholar] [CrossRef]
- Alam, M.-U.; Ferdous, S.; Ercumen, A.; Lin, A.; Kamal, A.; Luies, S.K.; Sharior, F.; Khan, R.; Rahman, Z.; Parvez, S.M.; et al. Effective Treatment Strategies for the Removal of Antibiotic-Resistant Bacteria, Antibiotic-Resistance Genes, and Antibiotic Residues in the Effluent From Wastewater Treatment Plants Receiving Municipal, Hospital, and Domestic Wastewater: Protocol for a Systematic Review. JMIR Res. Protoc. 2021, 10, e33365. [Google Scholar] [CrossRef]
- Meier, H.; Spinner, K.; Crump, L.; Kuenzli, E.; Schuepbach, G.; Zinsstag, J. State of Knowledge on the Acquisition, Diversity, Interspecies Attribution and Spread of Antimicrobial Resistance between Humans, Animals and the Environment: A Systematic Review. Antibiotics 2022, 12, 73. [Google Scholar] [CrossRef]
- Xiao, R.; Huang, D.; Du, L.; Song, B.; Yin, L.; Chen, Y.; Gao, L.; Li, R.; Huang, H.; Zeng, G. Antibiotic resistance in soil-plant systems: A review of the source, dissemination, influence factors, and potential exposure risks. Sci. Total. Environ. 2023, 869, 161855. [Google Scholar] [CrossRef]
- Kim, J.Y.; Suh, J.W.; Kang, J.S.; Kim, S.B.; Yoon, Y.K.; Sohn, J.W. Gram-Negative Bacteria’s Outer Membrane Vesicles. Infect. Chemother. 2023, 55, 557902. [Google Scholar] [CrossRef]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- Tang, K.L.; Caffrey, N.P.; Nóbrega, D.; Cork, S.C.; Ronksley, P.E.; Barkema, H.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.; et al. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: A systematic review and meta-analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef]
- WHO. Stop Using Antibiotics in Healthy Animals to Prevent the Spread of Antibiotic Resistance. 7 November 2017. Available online: https://www.who.int/news/item/07-11-2017-stop-using-antibiotics-in-healthy-animals-to-prevent-the-spread-of-antibiotic-resistance (accessed on 19 February 2023).
- Serwecińska, L. Antimicrobials and Antibiotic-Resistant Bacteria: A Risk to the Environment and to Public Health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- van Staa, T.P.; Palin, V.; Li, Y.; Welfare, W.; Felton, T.W.; Dark, P.; Ashcroft, D.M. The effectiveness of frequent antibiotic use in reducing the risk of infection-related hospital admissions: Results from two large population-based cohorts. BMC Med. 2020, 18, 40. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Sharma, S. Role of alternatives to antibiotics in mitigating the antimicrobial resistance crisis. Indian J. Med. Res. 2022, 156, 464–477. [Google Scholar]
- Ye, J.; Chen, X. Current Promising Strategies against Antibiotic-Resistant Bacterial Infections. Antibiotics 2022, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Guryanova, S.; Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry of Russian Academy of Sciences; Khaitov, R.; National Research Center—Institute of Immunology of the Federal Medico-Biological Agency. Glucosaminylmuramyldipeptide—GMDP: Effect on mucosal immunity (on the issue of immunotherapy and immunoprophylaxis). Immunologiya 2020, 41, 174–183. [Google Scholar] [CrossRef]
- Rechkina, E.A.; Denisova, G.F.; Masalova, O.V.; Lideman, L.F.; Denisov, D.A.; Lesnova, E.I.; Ataullakhanov, R.I.; Gur’Ianova, S.V.; Kushch, A. Epitope mapping of antigenic determinants of hepatitis C virus proteins by phage display. Mol. Biologiia 2006, 40, 357–368. (In Russian) [Google Scholar]
- Konorev, M.R.; Guryanova, S.V.; Tyshevich, E.N.; Pavlyukov, R.A.; Borisova, O.Y. Advisable including glucosaminylmuramyldipeptide in Helicobacter pylori therapy: Experience of ten-year investigation. Rudn. J. Med. 2020, 24, 269–282. [Google Scholar] [CrossRef]
- L’vov, V.L.; Gur’yanova, S.V.; Rodionov, A.V.; Gorshkova, R.P. Structure of the repeating unit of the O-specific polysaccha-ride of the lipopolysaccharide of yersinia kristensenii strain 490 (O:12,25). Carbohydr. Res. 1992, 228, 415–422. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- L’Vov, V.L.; Gur’Ianova, S.V.; Rodionov, A.V.; Dmitriev, B.A.; Shashkov, A.S.; Ignatenko, A.V.; Gorshkova, R.P.; Ovodov, I.S. The structure of a repetitive unit of the glycerolphosphate- containing O-specific polysaccharide chain from Yersinia kristensenii strain 103 (0:12,26) lipopolysaccharide. Bioorg. Khim. 1990, 16, 379–389. [Google Scholar] [PubMed]
- Tataurshchikova, N. OM-85: Personalized approach to the treatment of acute respiratory infections in children. Quest. Pract. Pediatr. 2020, 15, 61–68. [Google Scholar] [CrossRef]
- Ayswaria, R.; Vijayan, J.; Nathan, V.K. Antimicrobial peptides derived from microalgae for combating antibiotic resistance: Current status and prospects. Cell Biochem. Funct. 2023, 41, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Cooper, I.I. A review of the potential for bacteriophages to effect antibiofilm activity, using selected examples. J. Appl. Microbiol. 2022, 134, lxac056. [Google Scholar] [CrossRef]
- Tataurshchikova, N.S.; Sidorovich, I.G. The cytokine status as a criterion for the efficacy of intranasal aerosol therapy with the use of a cycloferon solution in the patients presenting with allergic rhinosinusitis. Vestn. Otorinolaringol. 2012, 3, 79–82. [Google Scholar]
- Al-Awsi, G.R.L.; Alameri, A.A.; Al-Dhalimy, A.M.B.; Gabr, G.A.; Kianfar, E. Application of nano-antibiotics in the diagnosis and treatment of infectious diseases. Braz. J. Biol. 2023, 84, e264946. [Google Scholar] [CrossRef]
- Guryanova, S.V.; Ovchinnikova, T.V. Immunomodulatory and Allergenic Properties of Antimicrobial Peptides. Int. J. Mol. Sci. 2022, 23, 2499. [Google Scholar] [CrossRef]
- Reeves, P. The Bacteriocins. Bacteriol. Rev. 1965, 29, 24–45. [Google Scholar] [CrossRef]
- Gratia, A. “Sur un remarquable exemple d’antagonisme entre deux souches de coilbacille” [On a remarkable example of antagonism between two strains of coilbacille]. Compt. Rend. Soc. Biol. 1925, 93, 1040–1042. NAID 10027104803. (In French) [Google Scholar]
- Fredericq, P. Colicines et Autres Bacteriocines [Colicines and Other Bacteriocins]. Ergeb. Mikrobiol. Immun. Exp. Ther. 1963, 37, 114–161. PMID: 14324288. (In French) [Google Scholar] [PubMed]
- Cotter, P.D.; Ross, R.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Genet. 2012, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Hill, C.; Ross, R.P. Food microbiology: Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777. [Google Scholar] [CrossRef]
- Wilaipun, P.; Zendo, T.; Sangjindavong, M.; Nitisinprasert, S.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. The two-synergistic peptide bacteriocin produced by Enterococcus faecium NKR- 5-3 isolated from Thai fermented fish (Plara). Sci. Asia 2004, 30, 115–122. [Google Scholar] [CrossRef]
- Fatima, D.; Mebrouk, K. Characterization and determination of the factors affecting anti-listerial bacteriocins from Lactobacillus plantarum and Pediococcus pentosaceus isolated from dairy milk products. Afr. J. Food Sci. 2013, 7, 35–44. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Darbandi, A.; Asadi, A.; Ari, M.M.; Ohadi, E.; Talebi, M.; Zadeh, M.H.; Emamie, A.D.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and potential use as antimicrobials. J. Clin. Lab. Anal. 2021, 36, e24093. [Google Scholar] [CrossRef]
- Benech, R.-O.; Kheadr, E.E.; Lacroix, C.; Fliss, I. Antibacterial Activities of Nisin Z Encapsulated in Liposomes or Produced In Situ by Mixed Culture during Cheddar Cheese Ripening. Appl. Environ. Microbiol. 2002, 68, 5607–5619. [Google Scholar] [CrossRef] [Green Version]
- Chandrakasan, G.; Rodríguez-Hernández, A.-I.; López-Cuellar, M.D.R.; Palma-Rodríguez, H.-M.; Chavarría-Hernández, N. Bacteriocin encapsulation for food and pharmaceutical applications: Advances in the past 20 years. Biotechnol. Lett. 2019, 41, 453–469. [Google Scholar] [CrossRef]
- Habib, W.; Sakr, A. Development and human in vivo evaluation of a colonic drug delivery system. Pharm. Ind. 1999, 61, 1145–1149. [Google Scholar]
- Gough, R.; Cabrera Rubio, R.; O’Connor, P.M.; Crispie, F.; Brodkorb, A.; Miao, S.; Hill, C.; Ross, R.P.; Cotter, P.D.; Nilaweera, K.N.; et al. Oral delivery of nisin in resistant starch based matrices alter the gut micro- biota in mice. Front Microbiol. 2018, 9, 1186. [Google Scholar] [CrossRef] [Green Version]
- Rollema, H.S.; Kuipers, O.P.; Both, P.; De Vos, W.M.; Siezen, R.J. Improvement of solubility and stability of the antimicrobial peptide nisin by protein engineering. Appl. Environ. Microbiol. 1995, 61, 2873–2878. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Blake, T.; Mathur, H.; O’Connor, P.M.; Cotter, P.D.; Ross, R.P.; Hill, C. Bioengineering Nisin to overcome the Nisin Resistance Protein. Mol. Microbiol. 2019, 111, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Duquesne, S.; Destoumieux-Garz’on, D.; Peduzzi, J.; Rebuffat, S. Microcins, gene-encoded antibacterial peptides from enterobacteria. Nat. Prod. Rep. 2007, 24, 708–734. [Google Scholar] [CrossRef] [PubMed]
- Drider, D.; Rebuffat, S. Prokaryotic Antimicrobial Peptides: From Genes to Applications; Springer Science & Business Media: Berlin/Heidelburg, Germany, 2011. [Google Scholar]
- Hammami, R.; Fernandez, B.; Lacroix, C.; Fliss, I. Anti-infective properties of bacteriocins: An update. Cell. Mol. Life Sci. 2013, 70, 2947–2967. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.; Cotter, P.; Ross, R.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtenberg, B.; Sc, Y.; Ch, L.; Ct, S.; Jy, F. Faculty Opinions recommendation of Antibacterial activities of bacteriocins: Application in foods and pharmaceuticals. Front Microbiol. 2014, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Riley, M.A.; Wertz, J.E. Bacteriocins evolution, ecology, and application. Annu. Rev. Microbiol. 2002, 56, 117–137. [Google Scholar] [CrossRef] [Green Version]
- Shand, R.F.; Leyva, K.J. Archaeal antimicrobials: An undiscovered country. In Archaea: New Models for Prokaryotic Biology; Caister Academic Press: Norfolk, UK, 2008; 248p. [Google Scholar]
- Besse, A.; Peduzzi, J.; Rebuffat, S.; Carré-Mlouka, A. Antimicrobial peptides and proteins in the face of extremes: Lessons from archaeocins. Biochimie 2015, 118, 344–355. [Google Scholar] [CrossRef]
- Oman, T.J.; van der Donk, W.A. Insights into the mode of action of the two-peptide lantibiotic haloduracin. ACS Chem. Biol. 2009, 4, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Hanchi, H.; Hammami, R.; Gingras, H.; Kourda, R.; Bergeron, M.G.; Ben Hamida, J.; Ouellette, M.; Fliss, I. Inhibition of MRSA and of Clostridium difficile by durancin 61A: Synergy with bacteriocins and antibiotics. Futur. Microbiol. 2017, 12, 205–212. [Google Scholar] [CrossRef]
- Cui, Z.; Chen, Z.-H.; Zhang, Q.; Gribova, V.V.; Filaretov, V.F.; Huang, D.-S. RMSCNN: A Random Multi-Scale Convolutional Neural Network for Marine Microbial Bacteriocins Identification. IEEE/ACM Trans. Comput. Biol. Bioinform. 2021, 19, 3663–3672. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Kolożyn-Krajewska, D. Food-Origin Lactic Acid Bacteria May Exhibit Probiotic Properties: Review. BioMed Res. Int. 2018, 2018, 5063185. [Google Scholar] [CrossRef] [Green Version]
- Verso, L.L.; Lessard, M.; Talbot, G.; Fernandez, B.; Fliss, I. Isolation and Selection of Potential Probiotic Bacteria from the Pig Gastrointestinal Tract. Probiotics Antimicrob. Proteins 2018, 10, 299–312. [Google Scholar] [CrossRef]
- Ryan, K.; Jayaraman, T.; Daly, P.; Canchaya, C.; Curran, S.; Fang, F.; Quigley, E.; O’toole, P. Isolation of lactobacilli with probiotic properties from the human stomach. Lett. Appl. Microbiol. 2008, 47, 269–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef] [PubMed]
- Gautam, N.; Sharma, N. Purification and characterization of bacteriocin produced by strain of Lactobacillus brevis MTCC 7539. Indian J. Biochem. Biophys. 2009, 46, 337–341. [Google Scholar] [PubMed]
- Knorr, D. Technology aspects related to microorganisms in functional foods. Trends Food Sci. Technol. 1998, 9, 295–306. [Google Scholar] [CrossRef]
- Karpinski, T.; Szkaradkiewicz, A.K. Characteristic of Bacteriocines and Their Application. Pol. J. Microbiol. 2013, 62, 223–235. [Google Scholar] [CrossRef]
- Choi, S.; Baek, M.-G.; Chung, M.-J.; Lim, S.; Yi, H. Distribution of bacteriocin genes in the lineages of Lactiplantibacillus plantarum. Sci. Rep. 2021, 11, 20063. [Google Scholar] [CrossRef]
- Collins, F.W.J.; O’Connor, P.M.; O’Sullivan, O.; Gómez-Sala, B.; Rea, M.C.; Hill, C.; Ross, R.P. Bacteriocin Gene-Trait matching across the complete Lactobacillus Pan-genome. Sci. Rep. 2017, 7, 3481. [Google Scholar] [CrossRef] [Green Version]
- Chérier, D.; Patin, D.; Blanot, D.; Touzé, T.; Barreteau, H. The Biology of Colicin M and Its Orthologs. Antibiotics 2021, 10, 1109. [Google Scholar] [CrossRef]
- De Jong, A.; Van Hijum, S.A.F.T.; Bijlsma, J.J.E.; Kok, J.; Kuipers, O.P. BAGEL: A web-based bacteriocin genome mining tool. Nucleic Acids Res. 2006, 34, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Spangler, R.; Zhang, S.P.; Krueger, J.; Zubay, G. Colicin synthesis and cell death. J. Bacteriol. 1985, 163, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Freire Bastos, M.d.C.; Coelho, M.L.V.; da Silva Santos, O.C. Resistance to bacteriocins produced by Gram-positive bacteria. Microbiol 2015, 161, 683–700. [Google Scholar] [CrossRef] [Green Version]
- Bountra, K.; Hagelueken, G.; Choudhury, H.G.; Corradi, V.; El Omari, K.; Wagner, A.; Mathavan, I.; Zirah, S.; Wahlgren, W.Y.; Tieleman, D.P.; et al. Structural basis for antibacterial peptide selfimmunity by the bacterial ABC transporter McjD. EMBO J. 2017, 36, 3062–3079. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [Green Version]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Circular and Leaderless Bacteriocins: Biosynthesis, Mode of Action, Applications, and Prospects. Front. Microbiol. 2018, 9, 2085. [Google Scholar] [CrossRef] [Green Version]
- Žgur-Bertok, D. Regulating colicin synthesis to cope with stress and lethality of colicin production. Biochem. Soc. Trans. 2012, 40, 1507–1511. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Moia, G.d.S.; Silva, A.; Baraúna, R.A.; Veras, A.A.d.O. BADASS: BActeriocin-Diversity ASsessment Software. BMC Bioinform. 2023, 24, 24. [Google Scholar] [CrossRef]
- Hammami, R.; Zouhir, A.; Cetal, L.L. BACTIBASE second release: A database and tool platform for bacteriocin characteriza-tion. BMC Microbiol 2010, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- The sbv IMPROVER Project Team (in Alphabetical Order); Boué, S.; Fields, B.; Hoeng, J.; Park, J.; Peitsch, M.C.; Schlage, W.K.; Talikka, M.; Binenbaum, I.; Bondarenko, V.; et al. Enhancement of COPD biological networks using a web-based collaboration interface. F1000Research 2015, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Guryanova, S.; Guryanova, A. sbv IMPROVER: Modern Approach to Systems Biology. Methods Mol. Biol. 2017, 1613, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Cai, J.; Zhang, B.; Wang, Y.; Wong, D.F.; Siu, S.W.I. Recent Progress in the Discovery and Design of Antimicrobial Peptides Using Traditional Machine Learning and Deep Learning. Antibiotics 2022, 11, 1451. [Google Scholar] [CrossRef] [PubMed]
- Andrès, E. Cationic antimicrobial peptides in clinical development, with special focus on thanatin and heliomicin. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 31, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Panina, I.S.; Balandin, S.V.; Tsarev, A.V.; Chugunov, A.O.; Tagaev, A.A.; Finkina, E.I.; Antoshina, D.V.; Sheremeteva, E.V.; Paramonov, A.S.; Rickmeyer, J.; et al. Specific Binding of the α-Component of the Lantibiotic Lichenicidin to the Peptidoglycan Precursor Lipid II Predetermines Its Antimicrobial Activity. Int. J. Mol. Sci. 2023, 24, 1332. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Bacteriocins. In Encyclopedia of Food and Health; Caballero, B., Ed.; Elsivier Science: Amsterdam, The Netherlands, 2016; pp. 312–319. [Google Scholar]
- Cotter, P.D. An ‘Upp’-turn in bacteriocin receptor identification. Mol. Microbiol. 2014, 92, 1159–1163. [Google Scholar] [CrossRef]
- Kouwen, T.R.H.M.; Trip, E.N.; Denham, E.L.; Sibbald, M.J.J.B.; Dubois, J.-Y.F.; van Dijl, J.M. The Large Mechanosensitive Channel MscL Determines Bacterial Susceptibility to the Bacteriocin Sublancin 168. Antimicrob. Agents Chemother. 2009, 53, 4702–4711. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Tarr, A.W.; Penfold, C.N. Colicin import into E. coli cells: A model system for insights into the import mechanisms of bacteriocins. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2014, 1843, 1717–1731. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Khait, R.; Harmalkar, A.; Pham, P.; Webby, M.N.; Housden, N.G.; Elliston, E.; Hopper, J.T.S.; Mohammed, S.; Robinson, C.V.; Gray, J.J.; et al. Colicin-Mediated Transport of DNA through the Iron Transporter FepA. mBio 2021, 12, e0178721. [Google Scholar] [CrossRef]
- Bieler, S.; Silva, F.; Soto, C.; Belin, D. Bactericidal Activity of both Secreted and Nonsecreted Microcin E492 Requires the Mannose Permease. J. Bacteriol. 2006, 188, 7049–7061. [Google Scholar] [CrossRef] [Green Version]
- James, R.; Lazdunski, C.; Pattus, F. Bacteriocins, Microcins and Lantibiotics; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; Volume 65. [Google Scholar]
- Chiumento, S.; Roblin, C.; Kieffer-Jaquinod, S.; Tachon, S.; Leprètre, C.; Basset, C.; Aditiyarini, D.; Olleik, H.; Nicoletti, C.; Bornet, O.; et al. Ruminococcin C, a promising antibiotic produced by a human gut symbiont. Sci. Adv. 2019, 5, eaaw9969. [Google Scholar] [CrossRef] [Green Version]
- Hatakka, K.; Saxelin, M. Probiotics in intestinal and non-intestinal infectious diseases—Clinical evidence. Curr. Pharm. Design. 2008, 14, 1351–1367. [Google Scholar] [CrossRef] [PubMed]
- Katla, T.; Naterstad, K.; Vancanneyt, M.; Swings, J.; Axelsson, L. Differences in Susceptibility of Listeria monocytogenes Strains to Sakacin P, Sakacin A, Pediocin PA-1, and Nisin. Appl. Environ. Microbiol. 2003, 69, 4431–4437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, B.; Guinane, C.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Assessing the Contributions of the LiaS Histidine Kinase to the Innate Resistance of Listeria monocytogenes to Nisin, Cephalosporins, and Disinfectants. Appl. Environ. Microbiol. 2012, 78, 2923–2929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, B.; Curtis, N.; Cotter, P.D.; Hill, C.; Ross, R.P. The ABC Transporter AnrAB Contributes to the Innate Resistance of Listeria monocytogenes to Nisin, Bacitracin, and Various β-Lactam Antibiotics. Antimicrob. Agents Chemother. 2010, 54, 4416–4423. [Google Scholar] [CrossRef] [Green Version]
- Rasch, M.; Knochel, S. Variations in tolerance of Listeria monocytogenes to nisin, pediocin PA-1 and bavaricin A. Lett. Appl. Microbiol. 1998, 27, 275–278. [Google Scholar] [CrossRef]
- Nes, I.F.; Gabrielsen, C.; Brede, D.A.; Diep, D.B. Novel developments in bacteriocins from lactic acid bacteria. In Biotechnology of Lactic Acid Bacteria: Novel Applications, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 80–99. [Google Scholar]
- Sedgley, C.M.; Clewell, D.B.; Flannagan, S.E. Plasmid pAMS1-Encoded, Bacteriocin-Related “Siblicide” in Enterococcus faecalis. J. Bacteriol. 2009, 191, 3183–3188. [Google Scholar] [CrossRef] [Green Version]
- Martãnez, B.; Rodrãguez, A. Antimicrobial susceptibility of nisin resistant Listeria monocytogenes of dairy origin. FEMS Microbiol. Lett. 2005, 252, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Maaß, S.; Bartel, J.; Mücke, P.-A.; Schlüter, R.; Sura, T.; Zaschke-Kriesche, J.; Smits, S.H.J.; Becher, D. Proteomic Adaptation of Clostridioides difficile to Treatment with the Antimicrobial Peptide Nisin. Cells 2021, 10, 372. [Google Scholar] [CrossRef]
- Lohans, C.T.; Vederas, J.C. Development of Class IIa Bacteriocins as Therapeutic Agents. Int. J. Microbiol. 2012, 2012, 386410. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Nijnik, A.; Magrangeas-Janot, L.; Pasupuleti, M.; Thorson, L.; Ma, S.; Easton, D.M.; Bains, M.; et al. Manipulation of innate immunity by a bacterial secreted peptide: Lantibiotic nisin Z is selectively immunomodulatory. J. Endotoxin Res. 2013, 19, 315–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pablo, M.A.; Gaforio, J.J.; Gallego, A.M.; Ortega, E.; Gã¡Lvez, A.M.; Lã³Pez, G.A.d.C. Evaluation of immunomodulatory effects of nisin-containing diets on mice. FEMS Immunol. Med. Microbiol. 1999, 24, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.M.; Ateia, I.; Paulus, J.R.; Liu, H.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Antimicrobial nisin acts against saliva derived multi-species biofilms without cytotoxicity to human oral cells. Front. Microbiol. 2015, 6, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.C.; Schroeder, H.E. Pathogenesis of inflammatory periodontal disease. A summary of current work. Lab. Investig. 1976, 34, 235. [Google Scholar] [PubMed]
- Radaic, A.; Brody, H.; Contreras, F.; Hajfathalian, M.; Lucido, L.; Kamarajan, P.; Kapila, Y.L. Nisin and Nisin Probiotic Disrupt Oral Pathogenic Biofilms and Restore Their Microbiome Composition towards Healthy Control Levels in a Peri-Implantitis Setting. Microorganisms 2022, 10, 1336. [Google Scholar] [CrossRef]
- Jia, Z.; He, M.; Wang, C.; Chen, A.; Zhang, X.; Xu, J.; Fu, H.; Liu, B. Nisin reduces uterine inflammation in rats by modulating concentrations of pro and antiinflammatory cytokines. Am. J. Reprod. Immunol. 2019, 81, e13096. [Google Scholar] [CrossRef]
- Gao, L.; Kuraji, R.; Zhang, M.J.; Martinez, A.; Radaic, A.; Kamarajan, P.; Le, C.; Zhan, L.; Ye, C.; Rangé, H.; et al. Nisin probiotic prevents inflammatory bone loss while promoting reparative proliferation and a healthy microbiome. NPJ Biofilms Microbiomes 2022, 8, 45. [Google Scholar] [CrossRef]
- Karimi, M.; Maghsoud, Z.; Halabian, R. Effect of Preconditioned Mesenchymal Stem Cells with Nisin Prebiotic on the Expression of Wound Healing Factors Such as TGF-β1, FGF-2, IL-1, IL-6, and IL-10. Regen. Eng. Transl. Med. 2021, 7, 30–40. [Google Scholar] [CrossRef]
- Małaczewska, J.; Kaczorek-Łukowska, E.; Wójcik, R.; Rękawek, W.; Siwicki, A.K. In vitro immunomodulatory effect of nisin on porcine leucocytes. J. Anim. Physiol. Anim. Nutr. 2019, 103, 882–893. [Google Scholar] [CrossRef]
- Śmiałek, J.; Bzowska, M.; Hinz, A.; Mężyk-Kopeć, R.; Sołtys, K.; Mak, P. Bacteriocin BacSp222 and Its Succinylated Forms Exhibit Proinflammatory Activities Toward Innate Immune Cells. J. Inflamm. Res. 2022, 15, 4601–4621. [Google Scholar] [CrossRef]
- Śmiałek, J.; Nowakowski, M.; Bzowska, M.; Bocheńska, O.; Wlizło, A.; Kozik, A.; Dubin, G.; Mak, P. Structure, Biosynthesis, and Biological Activity of Succinylated Forms of Bacteriocin BacSp222. Int. J. Mol. Sci. 2021, 22, 6256. [Google Scholar] [CrossRef] [PubMed]
- Cebrian, R.; Rodriguez-Cabezas, M.E.; Martín-Escolano, R.; Rubiño, S.; Barros, M.G.; Montalbán-López, M.; Rosales, M.J.; Sánchez-Moreno, M.; Valdivia, E.; Martínez-Bueno, M.; et al. Preclinical studies of toxicity and safety of the AS-48 bacteriocin. J. Adv. Res. 2019, 20, 129–139. [Google Scholar] [CrossRef]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory efficacy of nisin-a bacterial lantibiotic peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, J.; Yang, G.; Tao, L. Sublancin protects against methicillin-resistant Staphylococcus aureus infection by the combined modulation of innate immune response and microbiota. Peptides 2021, 141, 170533. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ye, Q.; Wang, K.; Zeng, X.; Huang, S.; Yu, H.; Ge, Q.; Qi, D.; Qiao, S. Enhancement of Macrophage Function by the Antimicrobial Peptide Sublancin Protects Mice from Methicillin-Resistant Staphylococcus aureus. J. Immunol. Res. 2019, 2019, 3979352. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Huang, S.; Ye, Q.; Zeng, X.; Yu, H.; Qi, D.; Qiao, S. Prevention of Cyclophosphamide-Induced Immunosuppression in Mice with the Antimicrobial Peptide Sublancin. J. Immunol. Res. 2018, 2018, 4353580. [Google Scholar] [CrossRef] [Green Version]
- Bu, Y.; Liu, Y.; Liu, Y.; Wang, S.; Liu, Q.; Hao, H.; Yi, H. Screening and Probiotic Potential Evaluation of Bacteriocin-Producing Lactiplantibacillus plantarum In Vitro. Foods 2022, 11, 1575. [Google Scholar] [CrossRef]
- Antoshina, D.V.; Balandin, S.V.; Bogdanov, I.V.; Vershinina, M.A.; Sheremeteva, E.V.; Toropygin, I.Y.; Finkina, E.I.; Ovchinnikova, T.V. Antimicrobial Activity and Immunomodulatory Properties of Acidocin A, the Pediocin-like Bacteriocin with the Non-Canonical Structure. Membranes 2022, 12, 1253. [Google Scholar] [CrossRef]
- Teiar, R.; Pérez-Ramos, A.; Zgheib, H.; Cudennec, B.; Belguesmia, Y.; Drider, D. Anti-adhesion and Anti-inflammatory Potential of the Leaderless Class IIb Bacteriocin Enterocin DD14. Probiotics Antimicrob. Proteins 2022, 14, 613–619. [Google Scholar] [CrossRef]
- Donovick, R.; Pagano, J.F.; Stout, H.A.; Weinstein, M.J. Thiostrepton, a new antibi-otic. I. In vitro studies. Antibiot Annu. 1955, 3, 554–559. [Google Scholar]
- Chiu, M.L.; Folcher, M.; Griffin, P.; Holt, T.; Klatt, T.; Thompson, C.J. Characterization of the covalent binding of thiostrepton to a thiostrepton-induced protein from Streptomyces lividans. Biochemistry 1996, 35, 2332–2341. [Google Scholar] [CrossRef] [PubMed]
- Halasi, M.; Zhao, H.; Dahari, H.; Bhat, U.G.; Gonzalez, E.B.; Lyubimo, A.V.; Tonetti, D.A.; Gartel, A.L. Thiazole antibiotics against breast cancer. Cell Cycle 2010, 9, 1214–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, S.; Lamore, S.D.; Cabello, C.M.; Lesson, J.L.; Munoz-Rodriguez, J.L.; Wondrak, G.T. Thiostrepton is an inducer of oxidative and proteotoxic stress that impairs viability of human melanoma cells but not primary melanocytes. Biochem. Pharmacol. 2012, 83, 1229–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newick, K.; Cunniff, B.; Preston, K.; Held, P.; Arbiser, J.; Pass, H.; Mossman, B.; Shukla, A.; Heintz, N. Peroxiredoxin 3 is a re-dox-dependent target of thiostrepton in malignant mesothelioma cells. PLoS ONE 2012, 7, e39404. [Google Scholar] [CrossRef] [Green Version]
- Hegde, N.S.; Sanders, D.A.; Rodriguez, R.; Balasubramanian, S. The transcription factor FOXM1 is a cellular target of the natural product thiostrepton. Nat. Chem. 2011, 3, 725–731. [Google Scholar] [CrossRef]
- Bhat, U.G.; Halasi, M.; Gartel, A.L. FoxM1 is a general target for proteasome in-hibitors. PLoS ONE 2009, 4, e6593. [Google Scholar] [CrossRef] [Green Version]
- Bailly, C. The bacterial thiopeptide thiostrepton. An update of its mode of action, pharmacological properties and applications. Eur. J. Pharmacol. 2022, 914, 174661. [Google Scholar] [CrossRef]
- Sandu, C.; Wetie, A.G.N.; Darie, C.C.; Steller, H. Thiostrepton, a Natural Compound That Triggers Heat Shock Response and Apoptosis in Human Cancer Cells: A Proteomics Investigation. Adv. Exp. Med. Biol. 2014, 806, 443–451. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Yeh, D.-W.; Lu, C.-H.; Liu, Y.-L.; Huang, L.-R.; Kao, C.-Y.; Chen, H.-Y.; Huang, C.-Y.F.; Chang, C.-H.; Luo, Y.; et al. Identification of Thiostrepton as a Novel Inhibitor for Psoriasis-like Inflammation Induced by TLR7–9. J. Immunol. 2015, 195, 3912–3921. [Google Scholar] [CrossRef] [Green Version]
- Salvador, P.B.U.; Dalmacio, L.M.M.; Kim, S.H.; Kang, D.-K.; Balolong, M.P. Immunomodulatory potential of four candidate probiotic Lactobacillus strains from plant and animal origin using comparative genomic analysis. Access Microbiol. 2021, 3, 000299. [Google Scholar] [CrossRef]
- Paiva, A.D.; de Oliveira, M.D.; de Paula, S.O.; Baracat-Pereira, M.C.; Breukink, E.; Mantovani, H.C. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiology 2012, 158, 2851–2858. [Google Scholar] [CrossRef]
- Dreyer, L.; Smith, C.; Deane, S.M.; Dicks, L.M.T.; van Staden, A.D. Migration of Bacteriocins across Gastrointestinal Epithelial and Vascular Endothelial Cells, as Determined Using In Vitro Simulations. Sci. Rep. 2019, 9, 11481. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications. Microb. Physiol. 2007, 13, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, B.; Le Lay, C.; Jean, J.; Fliss, I. Growth, acid production and bacteriocin production by probiotic candidates under simulated colonic conditions. J. Appl. Microbiol. 2013, 114, 877–885. [Google Scholar] [CrossRef]
- Kheadr, E.; Zihler, A.; Dabour, N.; Lacroix, C.; Le Blay, G.; Fliss, I. Study of the physicochemical and biological stability of pediocin PA-1 in the upper gastrointestinal tract conditions using a dynamic in vitro model. J. Appl. Microbiol. 2010, 109, 54–64. [Google Scholar] [CrossRef]
- Birri, D.J.; Brede, D.A.; Nes, I.F. Salivaricin D, a Novel Intrinsically Trypsin-Resistant Lantibiotic from Streptococcus salivarius 5M6c Isolated from a Healthy Infant. Appl. Environ. Microbiol. 2012, 78, 402–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.M.; Jung, Y.-G.; Jin, Y.-Y.; Jayabalan, R.; Yang, S.H.; Suh, J.W. Bacteriocins as food preservatives: Challenges and emerging horizons. Crit. Rev. Food Sci. Nutr. 2018, 58, 2743–2767. [Google Scholar] [CrossRef]
- Jarvis, B.; Mahoney, R. Inactivation of Nisin by Alpha-Chymotrypsin. J. Dairy Sci. 1969, 52, 1448–1450. [Google Scholar] [CrossRef]
- Gough, R.; O’Connor, P.M.; Rea, M.C.; Gómez-Sala, B.; Miao, S.; Hill, C.; Brodkorb, A. Simulated gastrointestinal digestion of nisin and interaction between nisin and bile. LWT 2017, 86, 530–537. [Google Scholar] [CrossRef] [Green Version]
- Pomares, M.F.; Salomón, R.A.; Pavlova, O.; Severinov, K.; Farías, R.; Vincent, P.A. Potential Applicability of Chymotrypsin-Susceptible Microcin J25 Derivatives to Food Preservation. Appl. Environ. Microbiol. 2009, 75, 5734–5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naimi, S.; Zirah, S.; Hammami, R.; Fernandez, B.; Rebuffat, S.; Fliss, I. Fate and Biological Activity of the Antimicrobial Lasso Peptide Microcin J25 under Gastrointestinal Tract Conditions. Front. Microbiol. 2018, 9, 1764. [Google Scholar] [CrossRef] [PubMed]
- Soleimanpour, M.; Mirhaji, S.S.; Jafari, S.; Derakhshankhah, H.; Mamashli, F.; Nedaei, H.; Karimi, M.R.; Motasadizadeh, H.; Fatahi, Y.; Ghasemi, A.; et al. Designing a new alginate-fibrinogen biomaterial composite hydrogel for wound healing. Sci. Rep. 2022, 12, 7213. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Pourakbari, B.; Bahador, A. Contribution of antimicrobial photo-sonodynamic therapy in wound healing: An in vivo effect of curcumin-nisin-based poly (L-lactic acid) nanoparticle on Acinetobacter baumannii biofilms. BMC Microbiol. 2022, 22, 28. [Google Scholar] [CrossRef]
- Cunha, E.; Tavares, L.; Oliveira, M. Revisiting Periodontal Disease in Dogs: How to Manage This New Old Problem? Antibiotics 2022, 11, 1729. [Google Scholar] [CrossRef]
- Lallukka, M.; Gamna, F.; Gobbo, V.A.; Prato, M.; Najmi, Z.; Cochis, A.; Rimondini, L.; Ferraris, S.; Spriano, S. Surface Functionalization of Ti6Al4V-ELI Alloy with Antimicrobial Peptide Nisin. Nanomaterials 2022, 12, 4332. [Google Scholar] [CrossRef]
- Azmi, M.F.; Al Khateeb, A.; Ab Rahim, S.; Froemming, G.R.A.; Omar, E. Effect of different solvents on nisin ZP potential as anticancer agent against MG-63 osteosarcoma cells. Asia Pac. J. Mol. Biol. Biotechnol. 2022, 30, 43–54. [Google Scholar] [CrossRef]
- Shin, J.; Gwak, J.; Kamarajan, P.; Fenno, J.; Rickard, A.; Kapila, Y. Biomedical applications of nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [Green Version]
- Thanjavur, N.; Sangubotla, R.; Lakshmi, B.A.; Rayi, R.; Mekala, C.D.; Reddy, A.S.; Viswanath, B. Evaluating the antimicrobial and apoptogenic properties of bacteriocin (nisin) produced by Lactococcus lactis. Process. Biochem. 2022, 122, 76–86. [Google Scholar] [CrossRef]
- Ortiz-Rodríguez, T.; Mendoza-Acosta, F.; Martínez-Zavala, S.A.; Salcedo-Hernández, R.; Casados-Vázquez, L.E.; Bideshi, D.K.; Barboza-Corona, J.E. Thurincin H Is a Nonhemolytic Bacteriocin of Bacillus thuringiensis with Potential for Applied Use. Probiotics Antimicrob. Proteins 2022, 6, 77–84. [Google Scholar] [CrossRef]
- Cox, P.S.C.A.M.S.G.C.R.; Coburn, P.S.; Gilmore, M.S. Enterococcal Cytolysin: A Novel Two Component Peptide System that Serves as a Bacterial Defense against Eukaryotic and Prokaryotic Cells. Curr. Protein Pept. Sci. 2005, 6, 77–84. [Google Scholar] [CrossRef]
- Taggar, R.; Jangra, M.; Dwivedi, A.; Bansal, K.; Patil, P.B.; Bhattacharyya, M.S.; Nandanwar, H.; Sahoo, D.K. Bacteriocin isolated from the natural inhabitant of Allium cepa against Staphylococcus aureus. World J. Microbiol. Biotechnol. 2021, 37, 20. [Google Scholar] [CrossRef]
- Aguilar-Pérez, C.; Gracia, B.; Rodrigues, L.; Vitoria, A.; Cebrián, R.; Deboosère, N.; Song, O.-R.; Brodin, P.; Maqueda, M.; Aínsa, J.A. Synergy between Circular Bacteriocin AS-48 and Ethambutol against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2018, 62, e00359-18. [Google Scholar] [CrossRef] [Green Version]
- Behrens, H.M.; Six, A.; Walker, D.; Kleanthous, C. The therapeutic potential of bacteriocins as protein antibiotics. Emerg. Top. Life Sci. 2017, 1, 65–74. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guryanova, S.V. Immunomodulation, Bioavailability and Safety of Bacteriocins. Life 2023, 13, 1521. https://doi.org/10.3390/life13071521
Guryanova SV. Immunomodulation, Bioavailability and Safety of Bacteriocins. Life. 2023; 13(7):1521. https://doi.org/10.3390/life13071521
Chicago/Turabian StyleGuryanova, Svetlana V. 2023. "Immunomodulation, Bioavailability and Safety of Bacteriocins" Life 13, no. 7: 1521. https://doi.org/10.3390/life13071521
APA StyleGuryanova, S. V. (2023). Immunomodulation, Bioavailability and Safety of Bacteriocins. Life, 13(7), 1521. https://doi.org/10.3390/life13071521