1. Introduction
Woodwasps (Hymenoptera: Siricidae) are important hymenopterans that seriously damage various species of coniferous and hardwood trees. Female wasps generally attack weakened and stressed trees, and they may lay up to 400 eggs in the outer sapwood of their hosts [
1]. Females carry arthrospores of a specific fungus,
Amylostereum spp., as well as phytotoxic mucus in specialized internal organs called mycangia. Both fungus and mucus are synchronously injected into oviposition holes through the bark along with eggs [
2]. The fungus produces enzymes capable of decaying the wood around larval tunnels, providing a convenient food source for larval feeding. Eventually, the fungus, phytotoxic mucus, and wasps work together to kill the chosen host tree.
Sirex noctilio Fabricius are a kind of destructive woodwasp that attack trees in their native habitats and they have been introduced as invasive pests around the world.
S. noctilio is native to southern Europe, North Africa, and parts of Asia, but it has been an invasive species in the Southern Hemisphere since the early 1900s, as well as in North America since 2005 [
3,
4].
S. noctilio was introduced into China in Daqing, Heilongjiang Province, in 2013, after which it spread to Liaoning, Jilin, and Inner Mongolia in northeastern China, where it reportedly attacks only
Pinus sylvestris var.
mongolica Litv. [
5,
6].
S. nitobei is distributed throughout Japan, with the exception of Hokkaido Island, as well as in North Korea and China (Fukuda et al. 1993). In China, the species is native and mainly distributed in the provinces of Shanxi, Yunnan, Inner Mongolia, and Hebei [
7].
S. nitobei poses a hazard to ancient and debilitated pines, such as the
Pinus tabuliformis in Xiangshan Park in Beijing, and is a significant threat to other pine species such as
P. armandi and
Larix spp. [
8].
The morphology of
S. noctilio and
S. nitobei is very similar, with the most obvious differences being the differing colors of the abdomen and hindfoot. In China, both species are associated with the same fungal symbiont,
Amylostereum areolatum [
5,
9]. Recently,
S. noctilio and
S. nitobei were found to be attacking
P. sylvestris var.
mongolica from June to September from 2016 to 2018 successively, in Jinbaotun town, Inner Mongolia Autonomous Region [
10].
S. noctilio adults started to emerge in the field from late June to early September; subsequently,
S. nitobei emerged from the same trees, with a peak emergence rate from late August through late September. In order to inhibit the spread of these woodwasps and prevent them from inflicting damage upon trees, integrated pest management strategies are urgently needed, but the development of such strategies requires detailed knowledge of the biology and behavior of these wasps, including the cellular mechanisms underlying mating and host selection, as well as the environmental factors influencing such behavior.
As an early invasive species, the mating and oviposition behavior of
S. noctilio has been studied in South Africa, New Zealand, and South America [
11,
12]. However, immediately following emergence, males consistently flew up to the crown of the tree upon which they had emerged [
2]. Therefore, mating took place on the upper branches of trees, making it difficult to observe how female and male siricids found each other in the wild [
13]. In a recent study by Caetano and Hajeck [
14], the mating behavior of
S. noctilio was observed in a laboratory setting. However, as an invasive species, the in situ mating behavior of
S. noctilio on
P. sylvestris var.
mongolica in China and factors influencing such behavior are important considerations for the development of pest control strategies, yet they remain poorly understood.
Although
S. nitobei is native to China, it is relatively understudied because of its low abundance and habit of attacking only dead or highly stressed trees with little economic value. However, recently,
S. nitobei has attracted attention due to its sympatric coexistence with the quarantine pest
S. noctilio. Takao et al. [
15] found that the life cycle of
S. nitobei was usually completed in one year, and it was reported that the species may have been more responsible for the death of Japanese pine trees than had previously been considered up to that point. Fukuda et al. [
16] found that the oviposition success of
S. nitobei was related to host tree conditions and its body size. Fitza et al. [
17] showed the mutualism relationship between
S. nitobei and
Amylostereum fungus symbionts.
The spread of
S. noctilio as an invasive species necessitates the development of a monitoring technique in newly invaded habitats. For example, trap trees treated with herbicide or girdling have been used to monitor and survey
S. noctilio populations [
18]. Kairomone (plant volatiles) lure traps were found to be the most effective method for trapping
S. noctilio individuals in areas where populations were large [
19]. Pheromones are commonly used as attractants to evaluate pest populations, and several pheromone compounds for
S. noctilio are known [
13,
20]. Female pheromones derived from cuticular washes were identified as (
Z)-7-heptacosene, (
Z)-7-nonacosene, and (
Z)-9-nonacosene; these pheromones were active in laboratory assay experiments and posited to be short-distance contact pheromones [
20]. A male-produced pheromone was identified as (
Z)-3-decen-1-ol and it was found to produce strong antennal response in males and females in gas chromatography-electroantennographic detection (GC-EAD) assays. In addition, (
Z)-3-decen-1-ol was found to be the most active substance in Y-tube olfactometer and wind tunnel assays. However, several unknown or suspected minor components, including (
Z)-4-decen-1-ol and (
E, E)-2,4-decadienal, were not identified by GC-MS and only active in behavioral tests [
13]. In contrast, no pheromones have been reported for
S. nitobei. However, as congeneric species,
S. noctilio and
S. nitobei might utilize similar or identical pheromones, therefore, previously identified pheromones with efficacy in
S. noctilio represent a rational starting point for an exploration of compounds with behavioral effects in
S. nitobei. Once such pheromones are identified, they have the potential to be used as effective synergistic components in the development of plant-derived lures.
In woodwasps such as S. noctilio and S. nitobei, pheromone release is closely related to mating behavior, and pheromones are useful tools for the development of pest evaluation and management strategies, which are urgently needed to protect vulnerable trees from woodwasp infestation in native and invaded habitats. Therefore, the objectives of this study were: (1) to investigate the mating behavior and circadian rhythms of S. noctilio and S. nitobei in China; (2) to analyze the factors influencing mating behavior in these two species; and (3) to identify male-produced pheromone components in both species based on three reported components from S. noctilio. The goal of this study was to provide useful basic biological information to researchers developing integrated pest management strategies, and to those developing monitoring strategies in particular.
2. Materials and Methods
2.1. Insects
Infested P. sylvestris var. mongolica were felled, cut into bolts of about 1 m in length, and sealed at the ends with paraffin in early May 2019, which was several months prior to adult emergence. Next, bolts with pores on the trunk were taken from the field into a quarantine room at Jinbaotun Forestry Station (JFS) in Inner Mongolia Autonomous Region, China (39°58′ N, 116°13′ E), and kept in a fine nylon mesh cage (3 m length, 3 m width, and 3 m height) until adult emergence. The average environmental conditions for log storage were approximately 24 ± 1 °C and 65–70% RH. The cages were monitored daily until siricid emergence.
After emergence, adult wasps were sexed and then placed individually in smaller cages (20 × 20 × 20 cm) with a piece of filter paper inside. The age of each adult was recorded. Subsequently, adults were transferred into other rooms with environmental conditions similar to those mentioned above. Cages containing individuals of the same sex were kept in each room, and exposure to direct sunlight was avoided. Prior to the mating behavior experiments, the wasps were not allowed to mate and were not exposed to any synthetic odor source.
Adults emerging between 0 and 24 h were considered to be 1-day-old individuals. Each wasp was used only once. The wasps used for mating observation were 1–7 days old.
2.2. Mating Behavior Observation and Circadian Rhythm
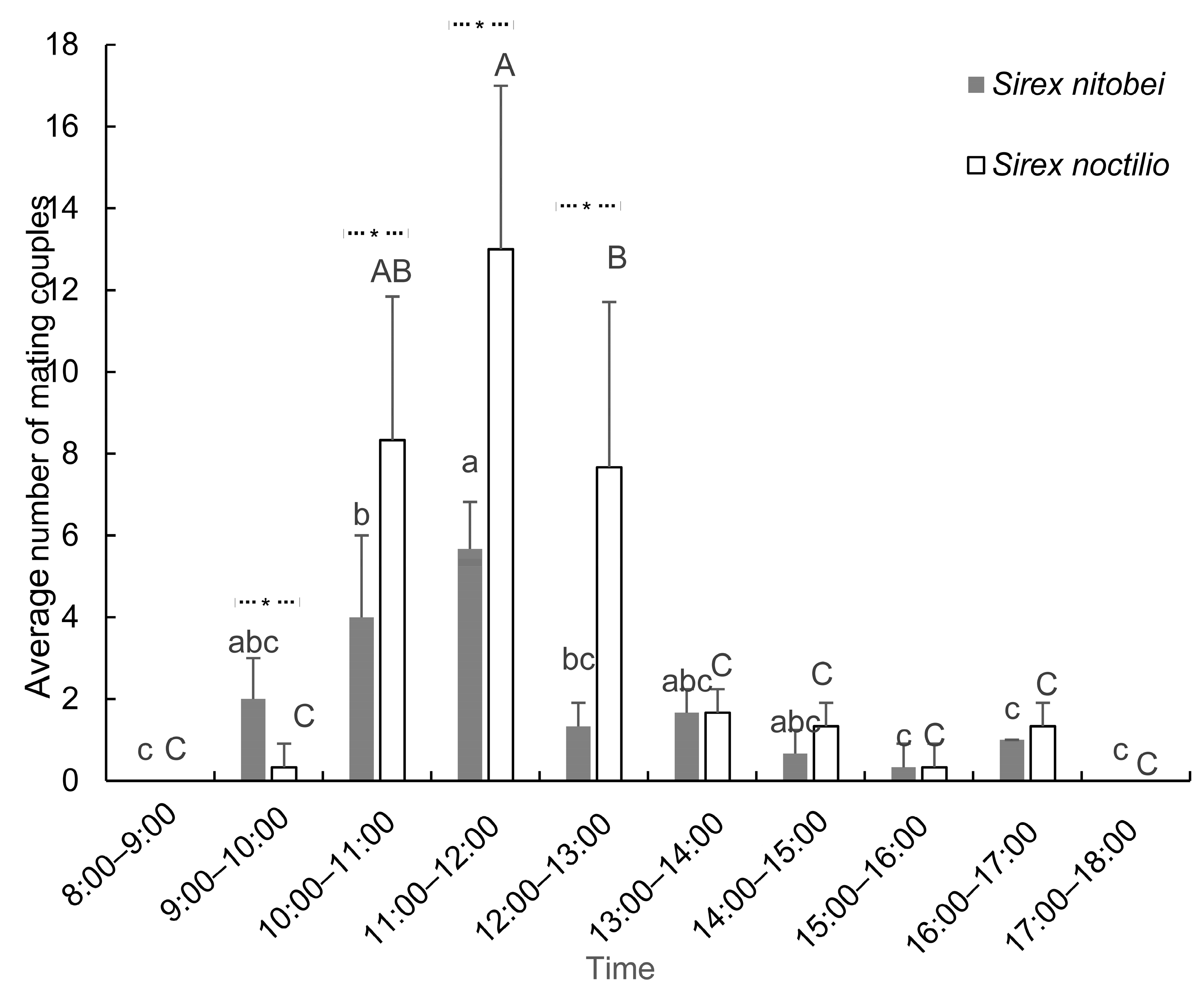
Upon emergence, active 2-day-old virgin wasps (11 female and 33 male) were introduced into a single cage (45 × 65 × 135 cm) to mate. The experiment was replicated three times (three cages). Observation began immediately after pairing. The daily mating rhythm was recorded every 20 min from 8:00 to 18:00. The onset time of mating and mating duration were recorded for each pair. The cumulative average number of pairs that had begun to mate in each cage was calculated once per hour and subjected to data analysis. Once per hour, light intensity was recorded by a TES1330A illuminometer (Tes Electrical Electronic Corp., Taipei, Taiwan, China), while temperature and humidity were recorded by a DeLi 9010 thermometer (Ningbo Deli E-Commerce Co., Ltd., Ningbo, China) The experiment was conducted in a quarantine room at JFS. During each observation, deceased individuals were removed from the cage and replaced with new 2-day-old active subjects.
2.3. Effect of Adult Body Size on Mating Success
After the trials described above, all mated females and males were collected, and their body length, thorax width, proala length, and ovipositor length were measured using a Vernier caliper (Pro’s Kit PD-151 Digital Caliper, Shenzhen Jin Tao Rui Technology Co., Ltd., Shenzhen, China).
2.4. Effect of Adult Age on the Mating Frequency
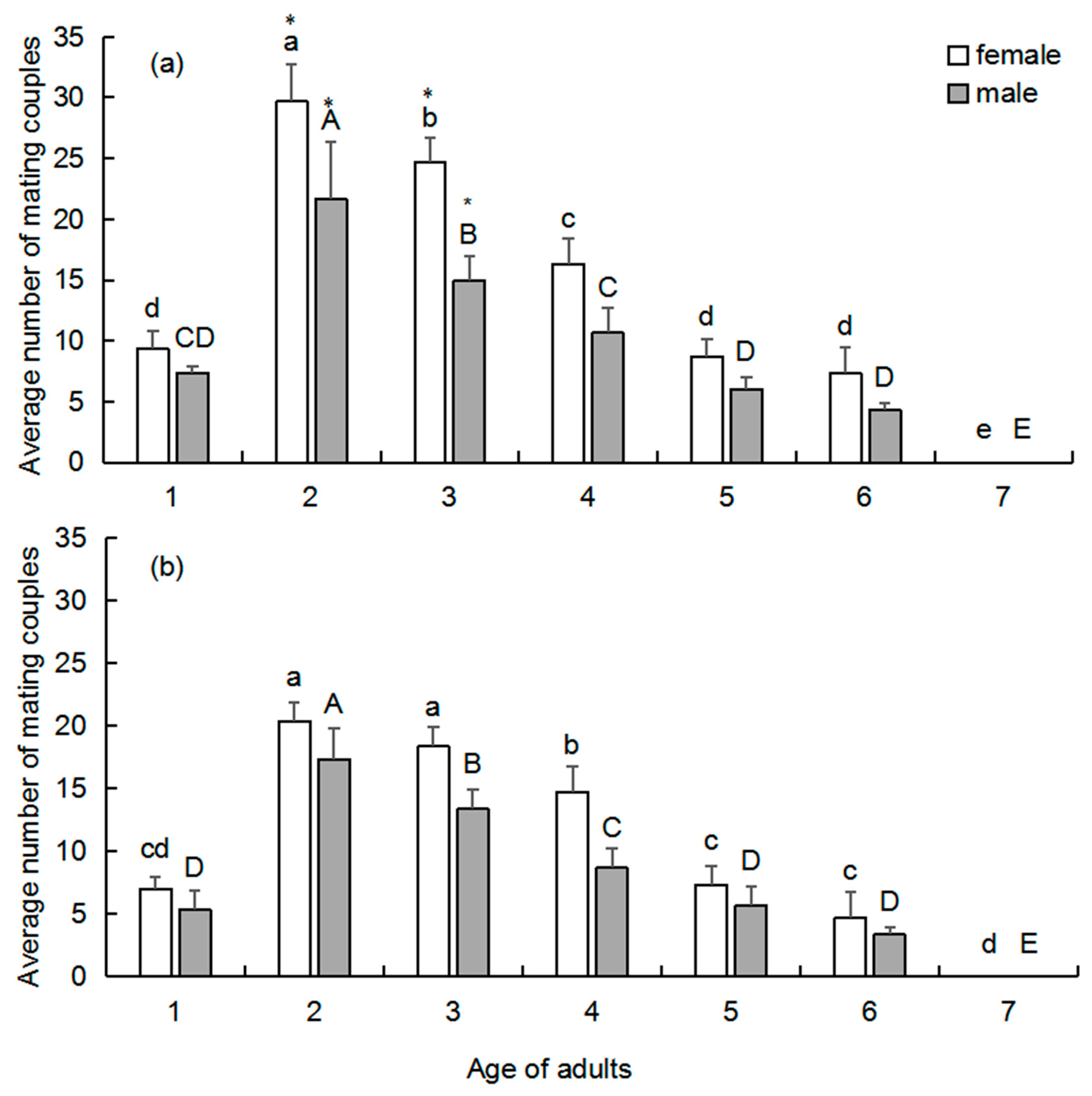
The effect of age on the mating frequency of female and male adults was studied. To test the effect of female age on mating frequency, 1-day-old females (N = 11) and 2-day-old males (N = 33) were introduced into a cage (45 × 65 × 135 cm) and observed continuously for 7 days. The age of the female subjects was gradually increased, but males were replaced with 2-day-old active wasps every day to ensure that their age remained constant. The same arrangement was applied to test the effect of male age on mating frequency, but the sexes of the retained and replaced groups were switched, i.e., females were replaced daily with 2-day-old active subjects, and males were allowed to age during the experiment. Three replicates (3 cages) were performed for each treatment. Observation began immediately after pairing. The observation was conducted continuously, and we count the cumulative average number of pairs during the whole 20 min from 10:00 to 13:00, which was the peak mating period. The cumulative average number of pairs was calculated once per hour and subjected to data analysis.
2.5. Effect of Sex Ratio on Daily Mating Frequency
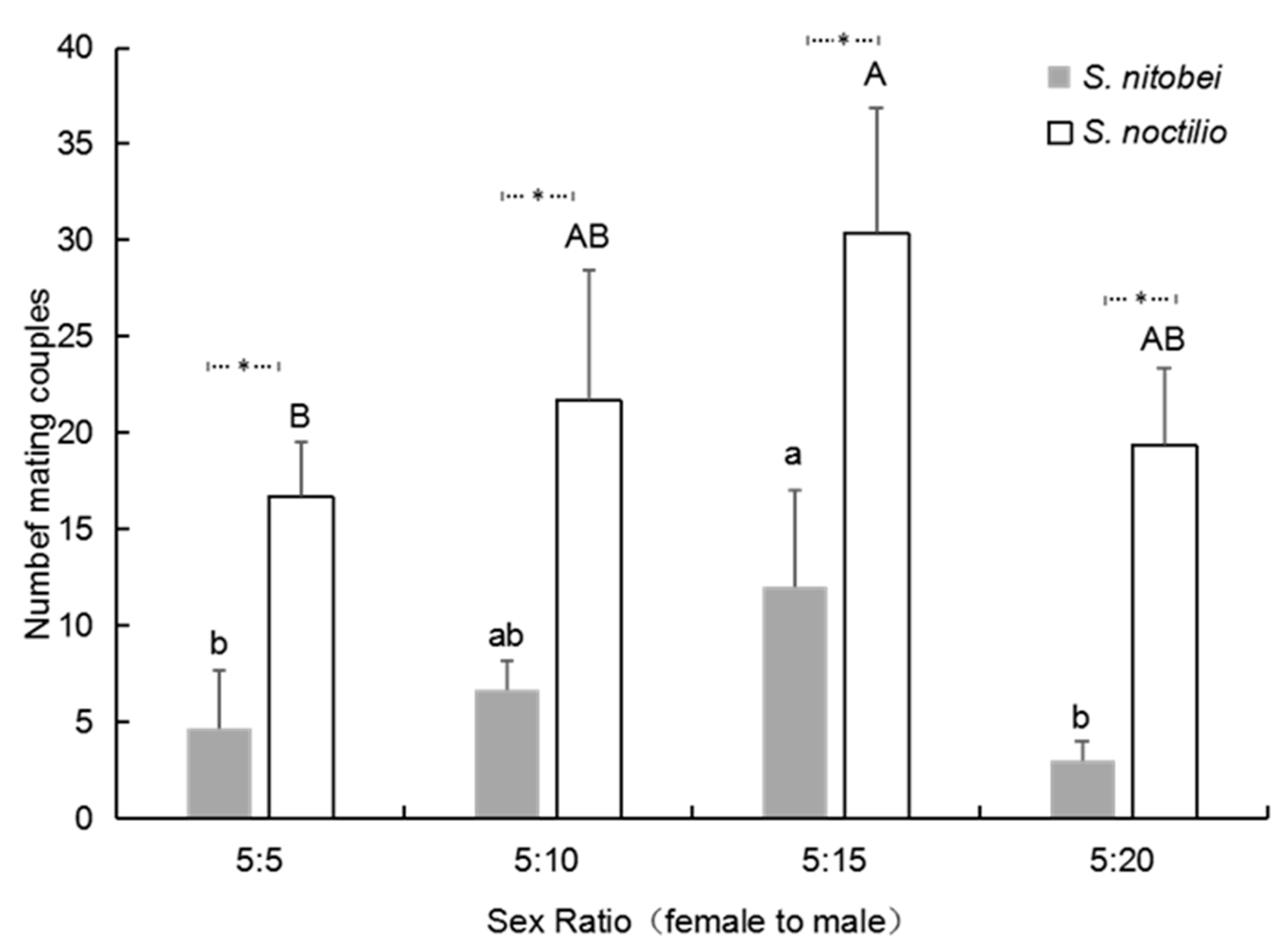
After emergence, 2-day-old virgin females and males were introduced into the copulating cage (45 × 65 × 135 cm) at different female: male ratios (5:5; 5:10, 5:15, 5:20) with 3 replicates (3 cages) for each ratio. Observation began immediately after pairing. Mating was recorded every 20 min from 10:00 to 13:00, which was the peak mating period. The cumulative average number of pairs that had begun to mate per cage was calculated once per hour and subjected to data analysis.
2.6. Headspace Collection of Male-Produced Pheromones
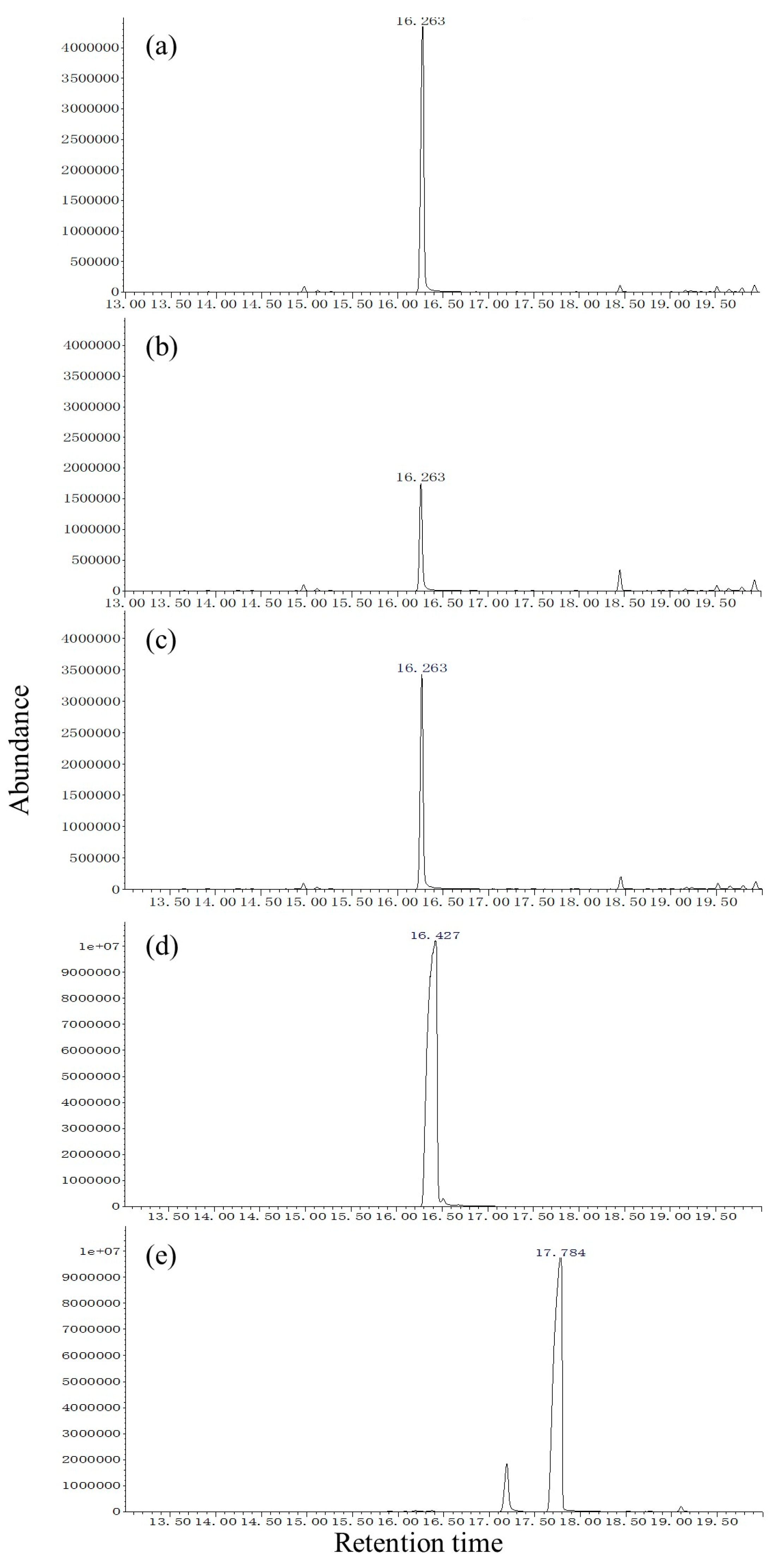
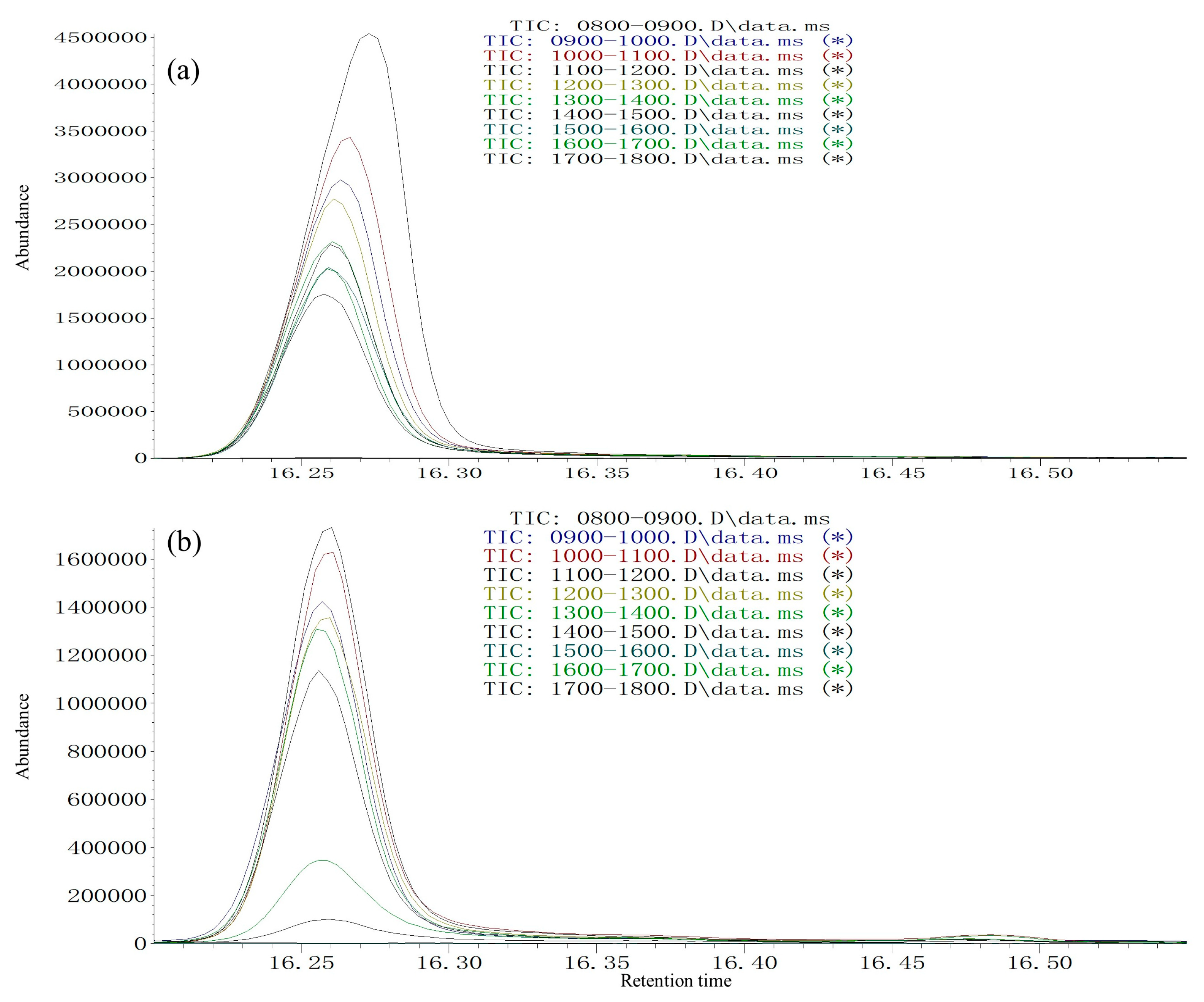
Volatiles were collected by headspace solid-phase microextraction (SPME) using a polydimethylsiloxane fiber (film thickness 100 µm; Supelco Inc., Bellefonte, PA, USA). A group of 15 virgin males (2-day-old) with average body weight was placed inside a sterile 500 mL tissue culture flask with a ventilation cap (Sarstedt, Leicester, UK). The SPME fiber was conditioned before use by baking it for 2 min at the injection port of a gas chromatograph (GC) at 270 °C, after which it was inserted through the cap and exposed to the sample headspace. Direct contact between the fiber and the wasps was prevented by a fine net surrounding the needle. Volatiles were collected from 8:00 to 18:00 at 1 h intervals (15 males). Three experimental replicates with different individuals (15 × 3 males) were used for each of the ten sampling time-points (once per hour from 8:00 to 18:00).
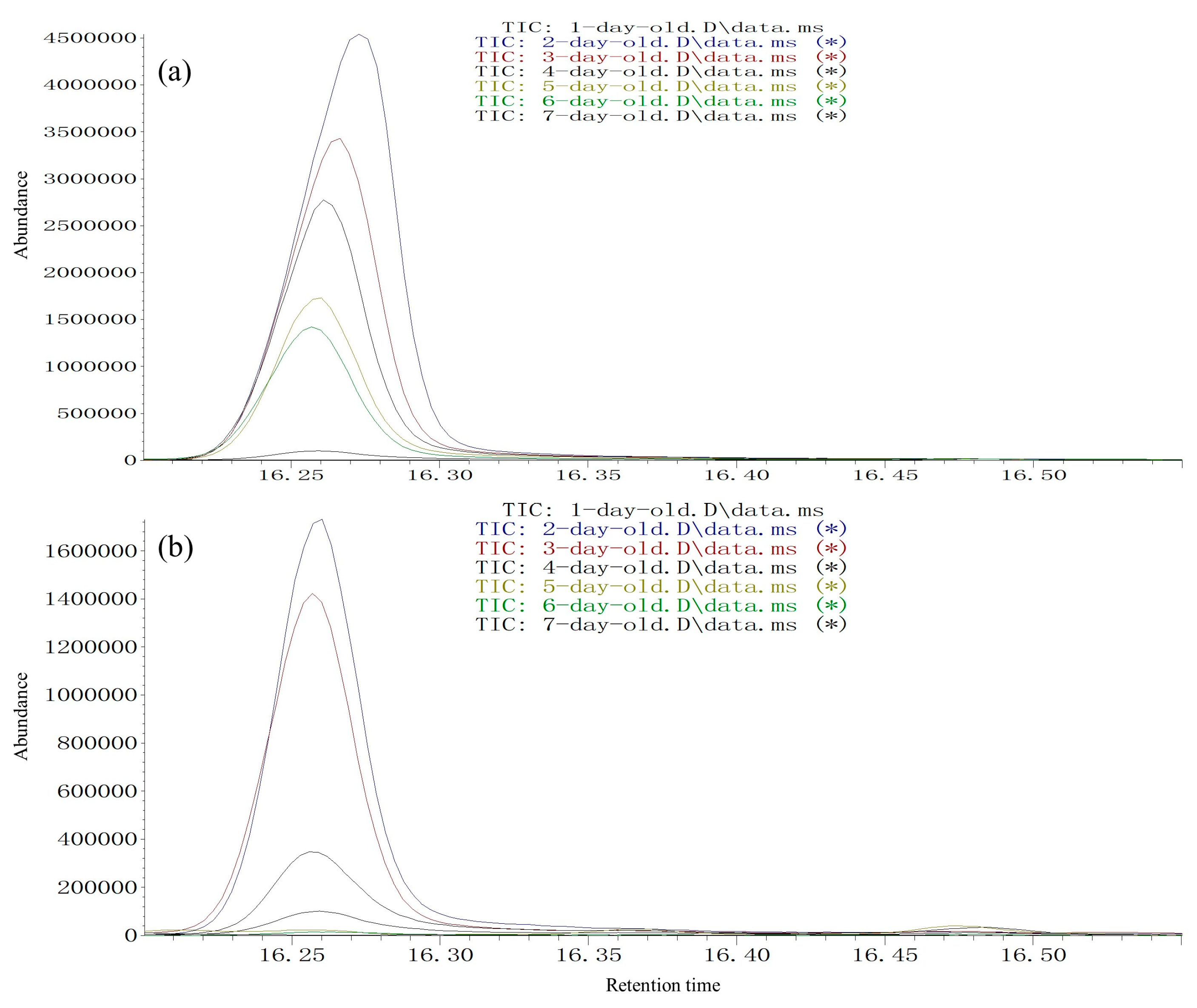
Age-dependent pheromone release was measured using virgin males aged from 1 to 7 days. For each test day, a group of 15 virgin males with average body weight was placed inside a sterile 500 mL tissue culture flask with a ventilation cap (Sarstedt, Leicester, UK). Volatiles were collected from 11:00 to 12:00 (15 males). For each of the seven groups (1, 2, 3, 4, 5, 6, or 7 days old), three experimental replicates (15 × 3 males) with different individuals were used for each sampling time point.
2.7. Gas Chromatography/Mass Spectrometry (GC/MS) Analysis
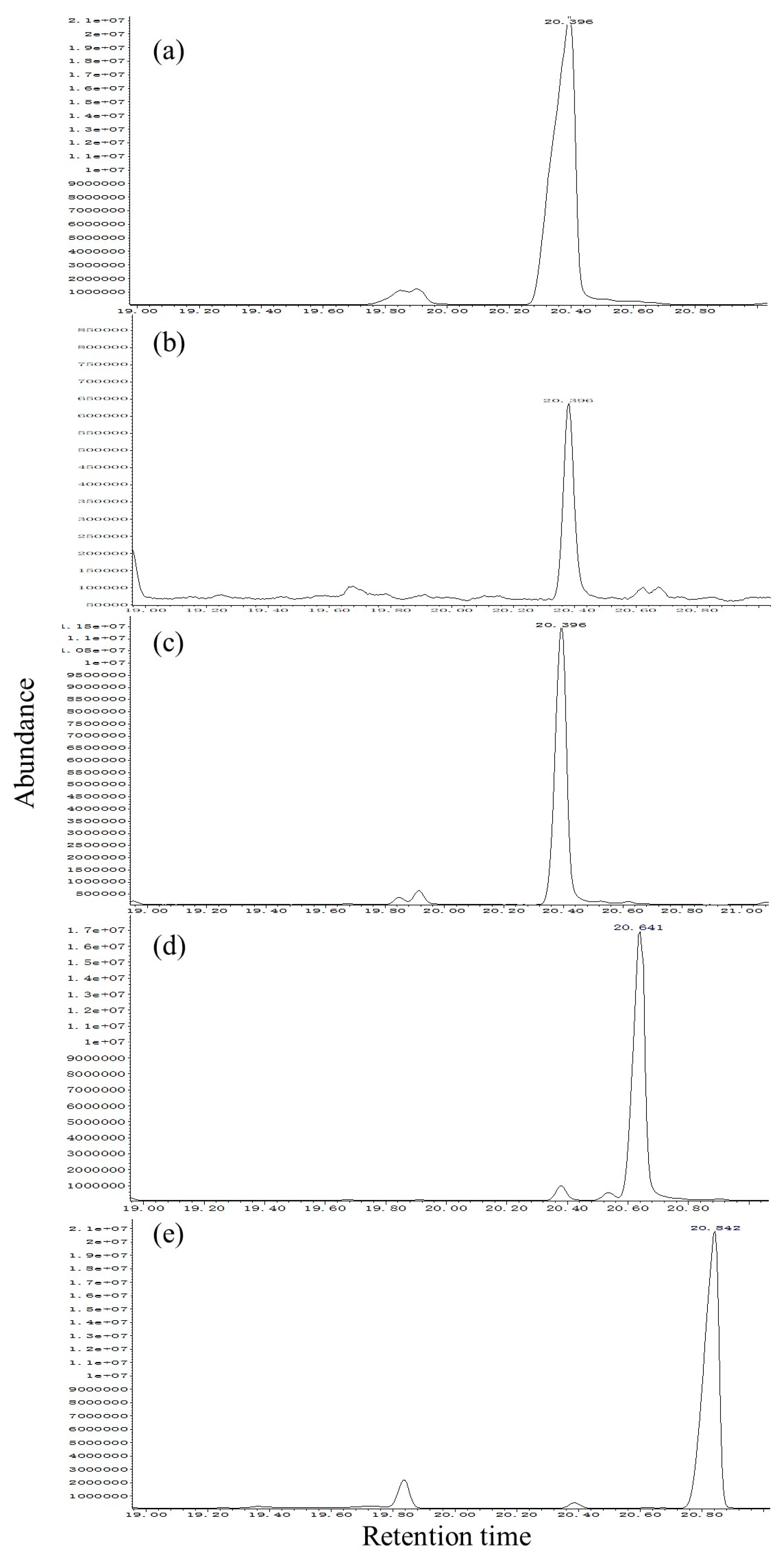
The SPME fibers were thermally desorbed into a splitless GC injector at 270 °C for 2 min. The GC/MS system consisted of an Agilent Technologies 5977A MS (Agilent, Santa Clara, CA, USA) coupled to an Agilent Technologies 7890 B GC (Agilent). For the HP-5 fused-silica column (30 m × 0.25 mm ID, 0.25 µm film, J&W Scientific Inc., Folsom, CA, USA), the initial oven temperature was maintained at 40 °C for 2 min, then increased at 6 °C/min to 180 °C, then increased at 15 °C/min to 270 °C. For the DB-WAX column (30 m × 0.25 mm ID, 0.25 µm film, J&W Scientific Inc., Folsom, CA, USA), the initial oven temperature was maintained at 50 °C for 1 min, then increased at 5 °C/min to 240 °C and maintained for 10 min. The NIST Mass Spectral Search Program (version 1.7) was used for data analysis. Injections were made in splitless mode. Helium was used as the carrier gas (1.0 mL/min). For electron impact (EI) mass spectra, the ionization voltage was 70 eV, the temperature of the ion source was 250 °C, and the temperature of the interface was 250 °C. Assignment of chemical structures to chromatographic peaks was based on comparison of their mass spectra fragmentation patterns and retention indexes with those of authentic standards. Circadian and age-dependent pheromone release was achieved by quantities expressed relative to the most abundant (Z)-3-decen-1-ol release period (set to a value of 100) during 11:00–12:00 in 2-day-old individuals for the two species.
2.8. Chemicals
The sex pheromone compound (Z)-3-decen-1-ol was purchased from Shuiguang Technology Co., Ltd. (Beijing, China). (Z)-4-decen-1-ol and (E, E)-2,4-decadienal was purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). n-Hexane was redistilled before use. The compounds were found to be 98% pure by GC analysis.
2.9. Statistical Analysis
The effect of the time of day on mating frequency, the effects of environmental factors at different times of the day, the effect of adult age on mating frequency, the effect of the sex ratio on mating, circadian and age-dependent pheromone release were analyzed by ANOVA. Tukey’s multiple range test was used to identify significant differences between means. Student’s paired-sample t-test was used to identify significant differences between body size of mated and unmated siricids. Correlational analysis was performed to assess the strength of the relationships between environmental parameters and mating behavior. Significant differences in mating circadian rhythms, effects of age and sex ratio on mating frequency between two siricids were analyzed by Mann–Whitney U-tests. All data were analyzed with SPSS v. 16.0 (IBM Co., Armonk, NY, USA).
4. Discussion
Observation of the mating behavior of woodwasps in their native forests is difficult because mating always occurs on the upper branches of trees. Here, the mating behavior of S. nitobei and S. nitobei was observed in outdoor cages. Compared with lepidopteron insects, these siricids did not show distinct calling behavior. In general, the mating behavior of S. nitobei was similar to that of S. noctilio: males approached females, touched antennae to find a suitable mating partner, and mated successfully. However, differences between these species were also observed. Compared with native species S. nitobei, the invasive species S. noctilio showed much stronger mating ability. Female and male S. noctilio cumulatively mated up to 19 and 13 times per day, respectively, and the average mating duration was 51.25 ± 25.54 s. However, female S. nitobei cumulatively mated up to 3 times per day, and males mated at most 4 times per day, while the average mating duration of the species was only 32.19 ± 16.02 s. The mating frequency of S. noctilio was much higher than that of S. nitobei regardless of the experimental context (circadian rhythm in a day, differences according to day, sex ratio).
Similar circadian rhythms were observed for both species, in which mating activity mainly occurred between 9:00 and 17:00 each day and peaked at 11:00 and 12:00. Caetano and Hajek [
14] reported peak mating activity from 13:00 to 15:35. Another woodwasp, horntail
Sirex rufi-abdomins, damages
P. massoniana in Anhui province in southern China [
21], and this species was found to mate mainly from 11:00 to 12:00. Taken together with these studies, our findings indicate that woodwasps generally mate around midday.
The mating behavior of both species was related to light intensity. Madden [
1] reported that male
S. noctilio swarmed near the bright tops of trees. Hurley et al. (2015) also reported that a clear intercept trap with light caught more female and male
S. noctilio in comparison with a clear intercept without light. In our study, males gathered quickly in the upper corner of the cage, towards the sun, once they had been put into the cage. These findings suggest that
S. noctilio and
S. nitobei are photopositive. Temperature also influences siricid mating. Caetano and Hajek [
13] reported that mating of
S. noctilio mainly occurred in the brightest part of the cage, and the number of mating sessions was positively associated with temperature, which was in accordance with our results. Madden [
1] found that the oviposition behavior of
S. noctilio was positively correlated with temperature and peaked at 20–22 °C. Morgan [
2] reported that the emergence behavior of adult
S. noctilio was affected by weather conditions; most insects emerged on bright and sunny days, whereas fewer insects emerged on cloudy, humid days, and no adults emerged on rainy days. In summary, environmental conditions with appropriate light intensity and temperature are beneficial for siricid reproductive behavior.
Mating behavior was found to be age-dependent for both tested woodwasp species.
S. noctilio and
S. nitobei can mate soon after emergence; most females and males of both species mated at 2-days-old. Cooperband et al. [
13] reported that male
S. noctilio were only attracted to odors from 2–5-day-old males and not to those from 0–1-day-old males, which suggested that 2-day-old males released more pheromones. Many studies have reported that appropriate adult age is associated with stronger reproductive characteristics, including female fertility, egg hatching rate, and longevity [
22,
23,
24,
25].
The mating behavior of the tested species was also found to be related to the sex ratio. In our study, a female-to-male ratio of 5:15 was the most conducive to mating for both species, but mating success decreased when the number of males was increased to 20. These results are indicative of a male-biased species, which tend to follow particular reproductive strategies. Firstly, these species generally mate multiple times. In our study, 41.9% of
S. nitobei females and 36.8% of
S. nitobei males mated more than once in their lives. Campbell [
26] indicated that the benefits of multiple mating by females exceeded the costs of refusing males, which was a waste of time and energy that also increased the risk of predation [
27]. After multiple mating sessions, females received more sperm. Furthermore, females who mated multiple times lived longer and showed greater fertility [
28]. Secondly, male-biased species generally have a relatively short mating duration. In our study,
S. nitobei spent 8–73 s mating, with an average of 32.19 ± 16.24 s. Average mating duration in
S. noctilio was 51.25 ± 25.54 s. Both mating durations were shorter than those of most insects, including hymenopterous parasitoid wasp
Campoletis chlorideae (162.45 ± 9.4 s) [
29], coleopterous
Monochamus alternatus (63.49 min) [
23], lepidopterous
Maruca vitrata (75 min) (Lu et al. 2008). A short mating duration can increase mating success and reduce the risk of predation.
Mating behavior was also found to be related to body size. Fukuda et al. [
30] showed that larger female adults were more likely to have reproductive success. Fukuda and Hijii [
30] indicated that longevity and the number of eggs laid by female
S. nitobei were positively correlated with fresh body weight, whereas male longevity was negatively correlated with fresh body weight. In our study, mated
S. noctilio had longer bodies, longer proala, longer ovipositors, and wider thoraxes in comparison with unmated individuals.
In our study, (
Z)-3-decenol was present in the SPME extracts of
S. noctilio and
S. nitobei populations. The morphology of
S. noctilio and
S. nitobei is very similar and they were reported to attack the same host,
P. sylvestris var.
mongolica, successively in Jinbaotun town, Inner Mongolia Autonomous Region, China. For the two sympatric species with similar pheromone components, what is the reproductive isolation mechanism? In Jinbaotun town, Inner Mongolia Autonomous Region,
S. noctilio adults started to emerge in the field from late June to early September; subsequently,
S. nitobei emerged from the same trees, with a peak emergence rate from late August through late September. Although the presence of the siricids in the region overlapped by a week or ten days at most, most of their development stage cannot meet in the field and they do not required special chemical cues to locate their own mates. However, several suspected minor components reported by Cooperband et al. [
13], including (
Z)-4-decen-1-ol) and
(E, E)-2,4-decadienal, were not identified by GC-MS in extracts from
S. nitobei or
S. noctilio. Further study should be conducted to elucidate the crucial and minor pheromone components in both species.
Mating activity and male pheromone (
Z)-3-decen-1-ol release were synchronized at different ages and times of the day in
S. noctilio. In general, reproduction behavior coincides with pheromone release in insect species [
24,
31,
32,
33]. In many lepidopterous moths, sex pheromones are always released by females, and changes in male responsiveness and locomotor activity are observed accordingly [
32,
33,
34]. The pheromone plume makes a powerful external stimulus to the receiver and could hypothetically function to synchronize rhythms in reproductive behaviors, which are primarily determined by rhythmic pheromone biosynthesis and endocrine system control [
35]. This kind of synchronization may arise for both sexes through reliance on the same external zeitgebers, such as the photoperiod, temperature, and other environmental factors, as indicated by our finding that rhythmic mating frequency, pheromone release, and corresponding environmental factors are closely associated with different ages and times of the day.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}