
Antifungal Potential of Capsaicinoids and Capsinoids from the Capsicum Genus for the Safeguarding of Agrifood Production: Advantages and Limitations for Environmental Health


 , , ,
, , ,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
1.1. Synthetic Fungicides and the One Health Concept
1.2. Capsicum spp. as a Source of Compounds with Natural Antifungal Activity
2. Materials and Methods
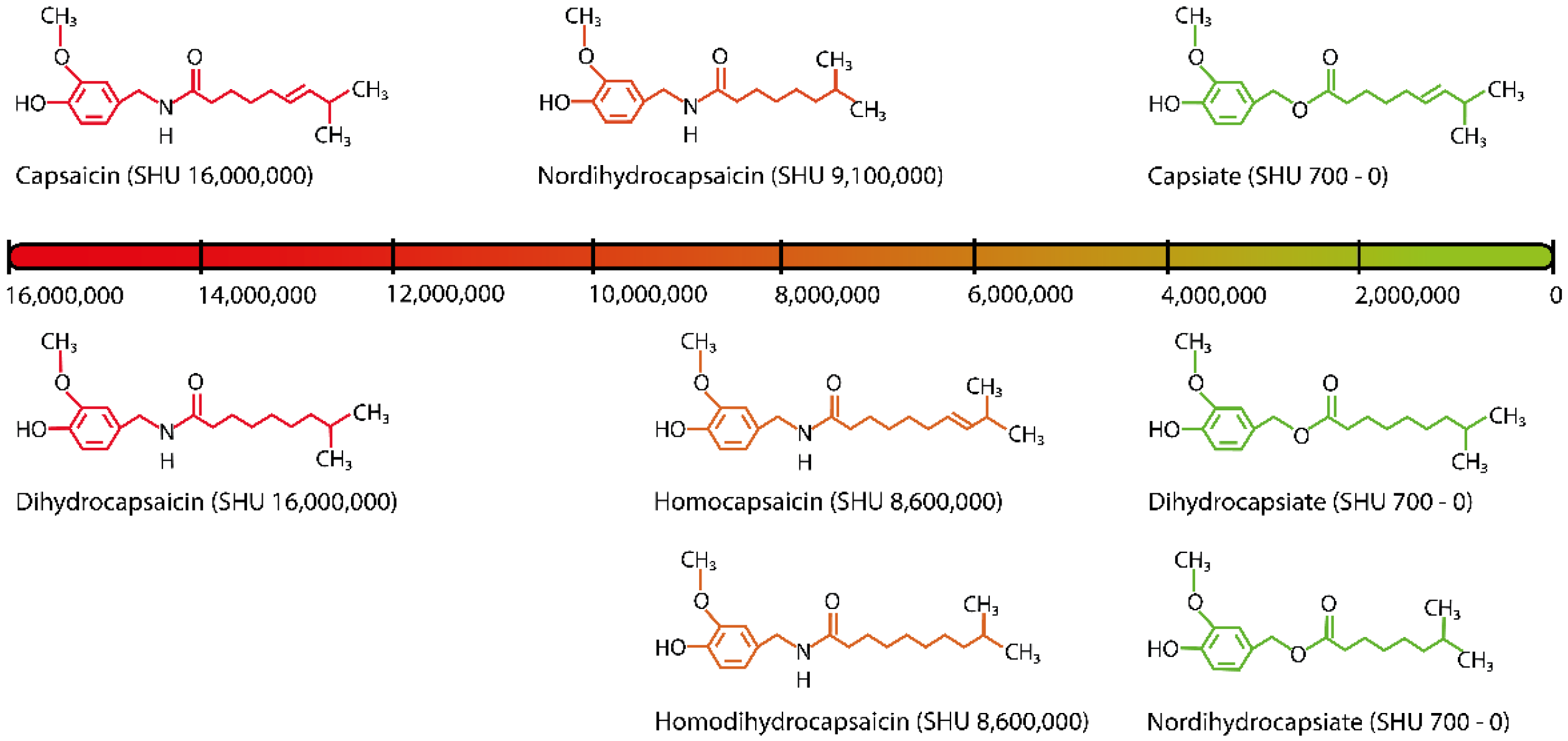
3. Characteristics of Capsaicinoids and Capsinoids
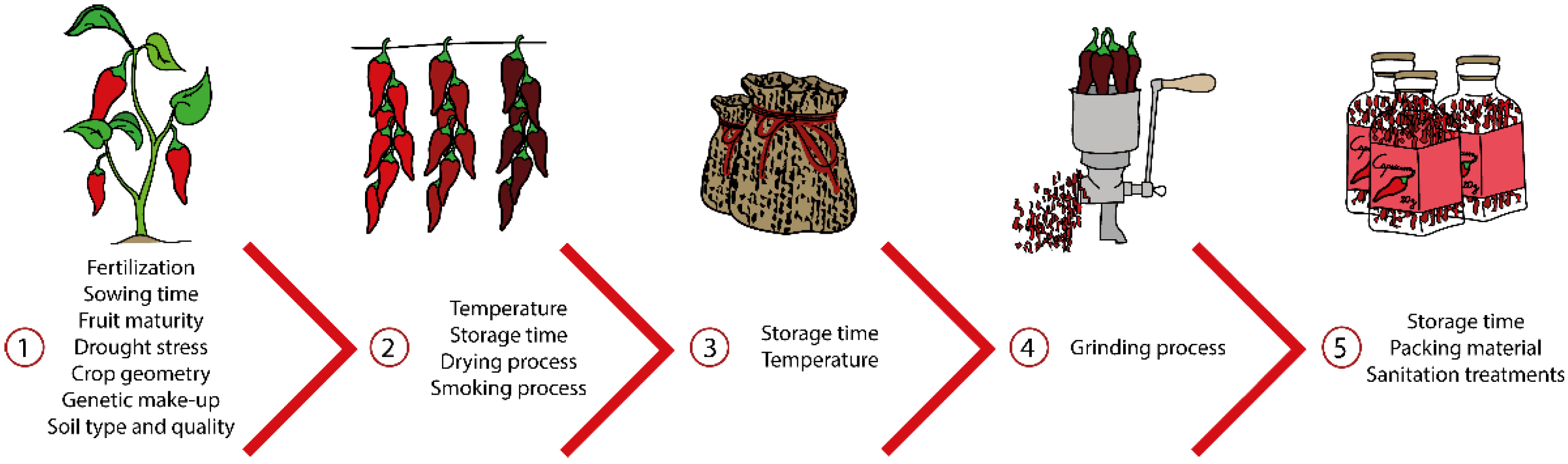
4. External Factors That Affect Capsaicinoids and Capsinoids
5. Capsaicinoids Antifungal Activity

{kind=link}
{kind=link}
| Plant Species | Pure Molecule (PM) or RAW Extract (RE) | Plant Substrate | Compound Name | Concentration | Fungal Taxa | Inhibition (%) | Reference |
|---|---|---|---|---|---|---|---|
| Capsicum annuum | RE | Seed | NI | 5 mg mL−1 | Colletotrichum gloeosporioides | 46.4 | [121] |
| Capsicum annuum | RE | Seed | NI | 10 mg mL−1 | Colletotrichum gloeosporioides | 54.6 | [121] |
| Capsicum annuum | RE | Seed | NI | 5 mg mL−1 | Colletotrichum gloeosporioides | 25.0 | [121] |
| Capsicum annuum | RE | Seed | NI | 10 mg mL−1 | Colletotrichum gloeosporioides | 38.1 | [121] |
| Capsicum annuum | RE | Pericarp | NI | 5 mg mL−1 | Colletotrichum gloeosporioides | 20.7 | [121] |
| Capsicum annuum | RE | Pericarp | NI | 10 mg mL−1 | Colletotrichum gloeosporioides | 43.6 | [121] |
| Capsicum annuum | RE | Pericarp | NI | 5 mg mL−1 | Colletotrichum gloeosporioides | 21.4 | [121] |
| Capsicum chinense | RE | Fruit | NI | 50 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 23.5 | [111] |
| Capsicum chinense | RE | Fruit | NI | 75 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 39.0 | [111] |
| Capsicum chinense | RE | Fruit | NI | 150 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 50.0 | [111] |
| Capsicum chinense | RE | Fruit | NI | 250 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 65.0 | [111] |
| Capsicum chinense | RE | Fruit | NI | 300 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 76.0 | [111] |
| Capsicum chinense | PM | Unknown | Capsaicin | 50 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 50.0 | [111] |
| Capsicum chinense | PM | Unknown | Capsaicin | 75 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 58.0 | [111] |
| Capsicum chinense | PM | Unknown | Capsaicin | 150 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 60.0 | [111] |
| Capsicum chinense | PM | Unknown | Capsaicin | 200 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 67.0 | [111] |
| Capsicum chinense | PM | Unknown | Capsaicin | 250 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 77.0 | [111] |
| Capsicum chinense | PM | Unknown | Capsaicin | 300 mg mL−1 | Aspergillus parasiticus ATCC 16992 | 80.0 | [111] |
| Capsicum frutescens | RE | Leaf | NI | 10 mg mL−1 | Aspergillus flavus | 88.1 | [122] |
| Capsicum frutescens | RE | Leaf | NI | 20 mg mL−1 | Aspergillus niger | 79.3 | [122] |
| Capsicum frutescens | RE | Leaf | NI | 5 mg mL−1 | Penicillium sp. | 20.5 | [122] |
| Capsicum frutescens | RE | Leaf | NI | 5 mg mL−1 | Rhizopus sp. | 69.0 | [122] |
| Capsicum frutescens | RE | Leaf | NI | 5 mg mL−1 | Aspergillus flavus | 79.2 | [122] |
| Capsicum frutescens | RE | Leaf | NI | 10 mg mL−1 | Aspergillus niger | 88.3 | [122] |
| Capsicum frutescens | RE | Leaf | NI | 1.25 mg mL−1 | Penicillium sp. | 32.9 | [122] |
| Capsicum frutescens | RE | Leaf | NI | 5 mg mL−1 | Rhizopus sp. | 77.2 | [122] |
| Capsicum frutescens | RE | Whole plant | NI | 3 µg mL−1 | Aspergillus niger | 91.4 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 3 µg mL−1 | Penicillium digitatum | 83.1 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 3 µg mL−1 | Fusarium sp. | 87.6 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 2 µg mL−1 | Aspergillus niger | 78.9 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 2 µg mL−1 | Penicillium digitatum | 74.0 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 2 µg mL−1 | Fusarium sp. | 69.3 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 1 µg mL−1 | Aspergillus niger | 73.2 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 1 µg mL−1 | Penicillium digitatum | 69.2 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 1 µg mL−1 | Fusarium sp. | 61.3 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 0.5 µg mL−1 | Aspergillus niger | 55.7 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 0.5 µg mL−1 | Penicillium digitatum | 51.5 | [123] |
| Capsicum frutescens | RE | Whole plant | NI | 0.5 µg mL−1 | Fusarium sp. | 49.0 | [123] |
| Capsicum sp. | RE | Fruit | NI | 500 mg mL−1 | Sphaeropsis sapinea | 100.0 | [124] |
| Capsicum sp. | RE | Fruit | NI | 500 mg mL−1 | Sphaeropsis sapinea | 100.0 | [124] |
| Capsicum sp. | RE | Fruit | NI | 350 mg mL−1 | Sphaeropsis sapinea | 40.0 | [124] |
| Capsicum sp. | RE | Fruit | NI | 350 mg mL−1 | Sphaeropsis sapinea | 100.0 | [124] |
| Capsicum sp. | PM | Unknown | Capsaicin | 25 mg mL−1 | Colletotrichum truncatum | 15.0 | [109] |
| Capsicum sp. | PM | Unknown | Capsaicin | 50 mg mL−1 | Colletotrichum truncatum | 35.0 | [109] |
| Capsicum sp. | PM | Unknown | Capsaicin | 100 mg mL−1 | Colletotrichum truncatum | 41.0 | [109] |
| Capsicum sp. | PM | Unknown | Capsaicin | 200 mg mL−1 | Colletotrichum truncatum | 59.0 | [109] |
| Capsicum sp. | PM | Unknown | Capsaicin | 122.16 µg mL–1 | Penicillium expansum | 75.0 | [125] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Verticillium dahliae VDL | 22.0 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Verticillium dahliae UDC53Vd | 35.3 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Verticillium dahliae 2694 | 0.9 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Verticillium dahliae 2884 | 19.9 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Verticillium tricorpus 2695 | 2.7 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Botrytis cinerea 2850 | 20.6 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Rhizoctonia solani 2815 | 32.0 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Fusarium oxysporum 2715 | 18.7 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Pythium ultimum 2364 | 29.2 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici P12M | 25.2 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici P15M | 44.4 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici UDC1Pc | 40.5 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici UDC141Pc | 46.8 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici UDC288Pc | 24.9 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici UDC299Pc | 73.2 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici UDC265Pc | 59.9 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Phytophthora capsici UDC384Pc | 55.8 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Verticillium dahliae VDL | 31.7 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Verticillium dahliae UDC53Vd | 46.1 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Verticillium dahliae 2694 | 3.7 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Verticillium dahliae 2884 | 29.5 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Verticillium tricorpus 2695 | 10.3 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 76.4 µg mL−1 | Botrytis cinerea 2850 | 35.9 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Rhizoctonia solani 2815 | 45.6 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Fusarium oxysporum 2715 | 33.3 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Pythium ultimum 2364 | 49.1 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici P12M | 38.7 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici P15M | 59.5 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici UDC1Pc | 62.4 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici UDC141Pc | 60.6 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici UDC288Pc | 65.5 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici UDC299Pc | 79.1 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici UDC265Pc | 69.6 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 152.7 µg mL−1 | Phytophthora capsici UDC384Pc | 68.3 | [100] |
| Capsicum sp. | PM | Unknown | Dihydrocapsaicin | 153.7 µg mL−1 | Verticillium dahliae | 37.4 | [100] |
| Capsicum sp. | PM | Unknown | Dihydrocapsaicin | 153.7 µg mL−1 | Verticillium dahliae | 62.6 | [100] |
| Capsicum sp. | PM | Unknown | Capsaicin | 60 mg mL−1 | Aspergillus carbonarius ATHUM 2854 | 89.7 | [110] |
| Capsicum sp. | PM | Unknown | Capsaicin | 60 mg mL−1 | Aspergillus section Nigri | 54.5 | [110] |
| Capsicum sp. | PM | Unknown | Capsaicin | 60 mg mL−1 | Aspergillus section Nigri ATHUM 6998 | 78.9 | [110] |
| Capsicum sp. | PM | Unknown | Capsaicin | 60 mg mL−1 | Aspergillus section Nigri ATHUM 6999 | 79.0 | [110] |
| Capsicum sp. | PM | Unknown | Capsaicin | 60 mg mL−1 | Aspergillus section Nigri ATHUM 7000 | 70.0 | [110] |
| Capsicum sp. | PM | Unknown | ABX-I | 115.6 mg mL−1 | Botrytis cinerea | 2.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 231.2 mg mL−1 | Botrytis cinerea | 9.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 462.4 mg mL−1 | Botrytis cinerea | 33.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 115.6 mg mL−1 | Colletotrichum gloeosporioides | 18.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 231.2 mg mL−1 | Colletotrichum gloeosporioides | 28.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 462.4 mg mL−1 | Colletotrichum gloeosporioides | 65.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 115.6 mg mL−1 | Rhizoctonia solani | 44.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 231.2 mg mL−1 | Rhizoctonia solani | 62.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 462.4 mg mL−1 | Rhizoctonia solani | 87.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 115.6 mg mL−1 | Fusarium sp. | 41.0 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 231.2 mg mL−1 | Fusarium sp. | 51 | [116] |
| Capsicum sp. | PM | Unknown | ABX-I | 462.4 mg mL−1 | Fusarium sp. | 67.0 | [116] |
| Capsicum sp. | PM | Unknown | Capsaicin | 244.3 µg mL−1 | Colletotrichum gloeosporioides | 60.0 | [116] |
| Capsicum sp. | PM | Unknown | Capsaicin | 244.3 µg mL−1 | Colletotrichum acutatum | 59.0 | [116] |
6. Impact of Using Capsaicinoids and Capsinoids on Environmental and Human Health
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO); Regional Office for Europe. A Health Perspective on the Role of the Environment in One Health; WHO/EURO:2022-5290-45054-64214; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- United Nations (UN). Global Perspective Human Stories. Available online: https://news.un.org/en/story/2022/07/1122272 (accessed on 10 September 2022).
- Food and Agriculture Organization (FAO). Food Outlook—Biannual Report on Global Food Markets; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal diseases as neglected pathogens: A wake-up call to public health officials. PLoS Negl. Trop. Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eurostat. 2022. Available online: https://ec.europa.eu/eurostat (accessed on 10 September 2022).
- Sáenz, V.; Alvarez-Moreno, C.; Pape, P.L.; Restrepo, S.; Guarro, J.; Ramírez, A.M.C. A one health perspective to recognize Fusarium as important in clinical practice. J. Fungi 2020, 6, 235. [Google Scholar] [CrossRef]
- Sun, S.; Lui, Q.; Han, L.; Ma, Q.; He, S.; Li, X.; Zhang, H.; Zhang, J.; Liu, X.; Wang, L. Identification and Characterization of Fusarium proliferatum, a New Species of Fungi that Cause Fungal Keratitis. Sci. Rep. 2018, 8, 4859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christy, J.S.; Balraj, A.; Agarwal, A. A rare case of Colletotrichum truncatum keratitis in a young boy with complete healing after medical treatment. Indian J. Med. Microbiol. 2020, 38, 475–477. [Google Scholar] [CrossRef] [PubMed]
- Iturrieta-González, I.; Giacaman, A.; Godoy-Martínez, P.; Vega, F.; Sepúlveda, M.; Santos, C.; Toledo, V.; Rivera, G.; Ortega, L.; San Martín, A.; et al. Penicillium digitatum, First Clinical Report in Chile: Fungal Co-Infection in COVID-19 Patient. J. Fungi 2022, 8, 961. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.N.; Heick, T.M. Azole Use in Agriculture, Horticulture, and Wood Preservation—Is It Indispensable? Front. Cell. Infect. Microbiol. 2021, 11, 730297. [Google Scholar] [CrossRef]
- Sallach, J.B.; Thirkell, T.J.; Field, K.J.; Carter, J.L. The emerging threat of human-use antifungals in sustainable and circular agriculture schemes. Plants People Planet 2021, 3, 685–693. [Google Scholar] [CrossRef]
- Atashgahi, S.; Shetty, S.A.; Smidt, H.; de Vos, W.M. Flux, Impact, and Fate of Halogenated Xenobiotic Compounds in the Gut. Front. Physiol. 2018, 9, 888. [Google Scholar] [CrossRef]
- Sarkar, S.; Gil, J.D.B.; Keeley, J.; Möhring, N.; Jansen, K. The Use of Pesticides in Developing Countries and Their Impact on Health and the Right to Food; European Parliament: Brussels, Belgium, 2021; pp. 1–56. [Google Scholar] [CrossRef]
- Belsky, J.; Joshi, N.K. Effects of Fungicide and Herbicide Chemical Exposure on Apis and Non-Apis Bees in Agricultural Landscape. Front. Environ. Sci. 2020, 8, 81. [Google Scholar] [CrossRef]
- Govindarajan, V.S. Capsicum-production, technology, chemistry and quality part1: History, botany, cultivation and primary processing. Crit. Rev. Food Sci. Nutr. 1985, 22, 109–176. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, R.H.; Thorpe, P.A. The antimicrobial properties of chile peppers (Capsicum species) and their uses in Mayan medicine. J. Ethnopharmacol. 1996, 52, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Lima, N.; Santos, C. Chilean pepper: Spoilage fungi and mycotoxins contamination risk in Capsicum products. In Conhecimento, Conservação e uso de Fungos, 1st ed.; Oliveira, L.A., Jesus, M.A., Jackisch-Matsuura, A.B., Gasparotto, L., Oliveira, L.G.S., Lima-Neto, R.G., Rocha, L.C., Eds.; Editora INPA: Manaus, Brazil, 2019; pp. 29–39. [Google Scholar]
- Vázquez-Espinosa, M.; Fayos, O.; González-De-Peredo, A.V.; Espada-Bellido, E.; Ferreiro-González, M.; Palma, M.; Garcés-Claver, A.; Barbero, G.F. Changes in capsiate content in four chili pepper genotypes (Capsicum spp.) at different ripening stages. Agronomy 2020, 10, 1337. [Google Scholar] [CrossRef]
- Costa, J.; Rodríguez, R.; Garcia-Cela, E.; Medina, A.; Magan, N.; Lima, N.; Battilani, P.; Santos, C. Overview of fungi and mycotoxin contamination in Capsicum pepper and in its derivatives. Toxins 2019, 11, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, R.; Abd-Elsayed, A. Capsaicin. In Pain, 2nd ed.; Abd-Elsayed, A., Ed.; Springer: Cham, Switzerland, 2019; pp. 339–341. [Google Scholar] [CrossRef]
- Scoville, W.L. Note on Capsicums. J. Am. Pharm. Assn. 1912, 1, 453–454. [Google Scholar] [CrossRef] [Green Version]
- Douventzidis, A.; Landquist, E. Logarithms Are Hot Stuff: A New Rating Scale for Chili Peppers. PRIMUS 2021, 32, 650–660. [Google Scholar] [CrossRef]
- Naves, E.R.; Silva, A.L.; Sulpice, R.; Araújo, W.L.; Nunes-Nesi, A.; Peres, L.E.P.; Zsögön, A. Capsaicinoids: Pungency beyond Capsicum. Trends Plant Sci. 2019, 24, 109–120. [Google Scholar] [CrossRef]
- Idrees, I.; Hanif, M.A.; Ayub, M.A.; Hanif, A.; Ansari, T.M. Chili Pepper. In Medicinal Plants of South Asia Novel Sources for Drug Discovery, 1st ed.; Hanif, M., Nawaz, H., Khan, M., Byrne, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 113–124. [Google Scholar] [CrossRef]
- Conforti, F.; Statti, G.A.; Menichini, F. Chemical and biological variability of hot pepper fruits (Capsicum annuum var. acuminatum L.) in relation to maturity stage. Food Chem. 2007, 102, 1096–1104. [Google Scholar] [CrossRef]
- Kosuge, S.; Furuta, M. Studies on the pungent principle of Capsicum. Agric. Biol. Chem. 1970, 34, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Al Othman, Z.A.; Ahmed, Y.B.; Habila, M.A.; Ghafar, A.A. Determination of capsaicin and dihydrocapsaicin in Capsicum fruit samples using high performance liquid chromatography. Molecules 2011, 16, 8919–8929. [Google Scholar] [CrossRef] [Green Version]
- Gudeva, L.K.; Mitrev, S.; Maksimova, V.; Spasov, D. Content of capsaicin extracted from hot pepper (Capsicum annuum spp. microcarpum L.) and its use as an ecopesticide. Hem. Ind. 2013, 67, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Antonio, A.S.; Wiedemann, L.S.M.; Veiga Junior, V.F. The genus: Capsicum: A phytochemical review of bioactive secondary metabolites. RSC Adv. 2018, 8, 25767–25784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, C.A.; Zimmerman, K.; Fenstermacher, K.; Thompson, M.G.; Skyrud, W.; Behie, S.; Pringle, A. Fungal seed pathogens of wild chili peppers possess multiple mechanisms to tolerate capsaicinoids. Appl. Environ. Microbiol. 2020, 86, e01697-19. [Google Scholar] [CrossRef]
- Vidal, C.; González, F.; Santander, C.; Pérez, R.; Gallardo, V.; Santos, C.; Aponte, H.; Ruiz, A.; Cornejo, P. Management of Rhizosphere Microbiota and Plant Production under Drought Stress: A Comprehensive Review. Plants 2022, 11, 2437. [Google Scholar] [CrossRef]
- Yazawa, S.; Suetom, N.; Okamoto, K.; Namiki, T. Content of Capsaicinoids and Capsaicinoid-like substances in fruit of pepper (Capsicum annuum L.) hybrids made with “CH-19 Sweet” as a parent. J. Jpn. Soc. Hort. Sci. 1989, 58, 601–607. [Google Scholar] [CrossRef]
- Kobata, K.; Todo, T.; Yazawa, S.; Iwai, K.; Watanabe, T. Novel Capsaicinoid-like Substances, Capsiate and Dihydrocapsiate, from the Fruits of a Nonpungent Cultivar, CH-19 Sweet, of Pepper (Capsicum annuum L.). J. Agric. Food Chem. 1998, 46, 1695–1697. [Google Scholar] [CrossRef]
- Nelson, E.K. The constitution of capsaicin, the pungent principle of Capsicum. J. Am. Chem. Soc. 1919, 41, 1115–1121. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Hosokawa, M.; Otsu, K.; Watanabe, T.; Yazawa, S. Assessment of capsiconinoid composition, nonpungent capsaicinoid analogues, in capsicum cultivars. J. Agric. Food Chem. 2009, 57, 5407–5412. [Google Scholar] [CrossRef]
- Haak, D.C.; McGinnis, L.A.; Levey, D.J.; Tewksbury, J.J. Why are not all chillies hot? A trade-off limits pungency. Proc. Biol. Sci. 2012, 279, 2012–2017. [Google Scholar] [CrossRef] [Green Version]
- Uarrota, V.G.; Maraschin, M.; de Bairros, Â.d.F.M.; Pedreschi, R. Factors affecting the capsaicinoid profile of hot peppers and biological activity of their non-pungent analogs (Capsinoids) present in sweet peppers. Crit. Rev. Food Sci. Nutr. 2021, 61, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Gour, J.K.; Rizvi, S.I. Capsaicin has potent anti-oxidative effects in vivo through a mechanism which is non-receptor mediated. Arch. Physiol. Biochem. 2019, 128, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Yu, X.; Zheng, Z.; Huang, J.; Yang, X.; Shi, H. Capsaicin suppressed activity of prostate cancer stem cells by inhibition of Wnt/β-catenin pathway. Phytother. Res. 2020, 34, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Zhang, X.; Guo, Y. Identification of the dietary supplement capsaicin as an inhibitor of Lassa virus entry. Acta Pharm. Sin. B 2020, 10, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Ayariga, J.A.; Abugri, D.A.; Amrutha, B.; Villafane, R. Capsaicin Potently Blocks Salmonella typhimurium Invasion of Vero Cells. Antibiotics 2022, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the emerging threat of antifungal resistance to human health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef]
- Kobata, K.; Kawaguchi, M.; Watanabe, T. Enzymatic synthesis of a capsinoid by the acylation of vanillyl alcohol with fatty acid derivatives catalyzed by lipases. Biosci. Biotechnol. Biochem. 2002, 66, 319–327. [Google Scholar] [CrossRef]
- Kobata, K.; Sutoh, K.; Todo, T.; Yazawa, S.; Iway, K.; Watanabe, T. Nordihydrocapsiate, a new capsinoid from the fruits of a nonpungent pepper, Capsicum annuum. J. Nat. Prod. 1999, 62, 335–336. [Google Scholar] [CrossRef]
- Zewdie, Y.; Bosland, P.W. Evaluation of genotype, environment, and genotype-by-environment interaction for capsaicinoids in Capsicum annuum L. Euphytica 2000, 111, 185–190. [Google Scholar] [CrossRef]
- Park, M.; Lee, J.H.; Han, K.; Jang, S.; Han, J.; Lim, J.H.; Jung, J.W.; Kang, B.C. A major QTL and candidate genes for capsaicinoid biosynthesis in the pericarp of Capsicum chinense revealed using QTL-seq and RNA-seq. Theor. Appl. Genet. 2019, 132, 515–529. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Nabhan, G.P. Seed dispersal: Directed deterrence by capsaicin in chillies. Nature 2001, 412, 403–404. [Google Scholar] [CrossRef] [PubMed]
- Levey, D.J.; Tewksbury, J.J.; Cipollini, M.L.; Carlo, T.A. A field test of the directed deterrence hypothesis in two species of wild chili. Oecologia 2006, 150, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Shilpi, J.A.; Uddin, S.J. Analgesic and antipyretic natural products. Annu. Rep. Med. Chem. 2020, 55, 435–458. [Google Scholar] [CrossRef]
- Aza-González, C.; Núñez-Palenius, H.G.; Ochoa-Alejo, N. Molecular biology of capsaicinoid biosynthesis in chili pepper (Capsicum spp.). Plant Cell Rep. 2011, 30, 695–706. [Google Scholar] [CrossRef]
- Reyes-Escogido, M.L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and Pharmacological Aspects of Capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef] [Green Version]
- Vera-Guzmán, A.M.; Chávez-Servia, J.L.; Carrillo-Rodríguez, J.C.; López, M.G. Phytochemical Evaluation of Wild and Cultivated Pepper (Capsicum annuum L. and C. pubescens Ruiz & Pav.) from Oaxaca, Mexico. Chil. J. Agric. Res. 2011, 71, 578–585. [Google Scholar] [CrossRef]
- Aguiar, A.C.; Viganó, J.; Anthero, A.G.S.; Dias, A.L.B.; Hubinger, M.D.; Martínez, J. Supercritical fluids and fluid mixtures to obtain high-value compounds from Capsicum peppers. Food Chem. 2022, 13, 100228. [Google Scholar] [CrossRef]
- Oğuzkan, S.B. Extraction of Capsinoid and its Analogs from Pepper Waste of Different Genotypes. Nat. Prod. Commun. 2019, 14, 1–5. [Google Scholar] [CrossRef]
- Hursel, R.; Westerterp-Plantenga, M. Thermogenic ingredients and body weight regulation. Int. J. Obes. 2010, 34, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Rodríguez, R.; Santos, C.; Soares, C.; Lima, N.; Santos, C. Mycobiota in Chilean chilli Capsicum annuum L. used for production of Merkén. Int. J. Food Microbiol. 2020, 334, 108833. [Google Scholar] [CrossRef]
- Costa, J.; Santos, C.; Soares, C.; Rodríguez, R.; Lima, N.; Santos, C. Occurrence of Aflatoxins and Ochratoxin A during Merkén pepper powder production in Chile. Foods 2022. accepted for publication. [Google Scholar]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; G Wasef, L.; Elewa, Y.; A Al-Sagan, A.; Abd El-Hack, M.E.; Taha, A.E.; M Abd-Elhakim, Y.; Prasad Devkota, H. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeeatid, N.; Techawongstien, S.; Suriharn, B.; Chanthai, S.; Bosland, P.W.; Techawongstien, S. Influence of water stresses on capsaicinoid production in hot pepper (Capsicum chinense Jacq.) cultivars with different pungency levels. Food Chem. 2018, 245, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, R. Stable capsaicinoid biosynthesis during the fruit development stage of Capsicum baccatum. Cytologia 2019, 84, 309–312. [Google Scholar] [CrossRef]
- Mahmood, T.; Rana, R.M.; Ahmar, S.; Saeed, S.; Gulzar, A.; Khan, M.A.; Wattoo, F.M.; Wang, X.; Branca, F.; Mora-Poblete, F.; et al. Effect of drought stress on capsaicin and antioxidant contents in pepper genotypes at reproductive stage. Plants 2021, 10, 1286. [Google Scholar] [CrossRef] [PubMed]
- Darko, E.; Hamow, K.A.; Marček, T.; Dernovics, M.; Ahres, M.; Galiba, G. Modulated Light Dependence of Growth, Flowering, and the Accumulation of Secondary Metabolites in Chilli. Front. Plant Sci. 2022, 13, 801656. [Google Scholar] [CrossRef]
- Stan, T.; Munteanu, N.; Teliban, G.-C.; Cojocaru, A.; Stoleru, V. Fertilization Management Improves the Yield and Capsaicinoid Content of Chili Peppers. Agriculture 2021, 11, 181. [Google Scholar] [CrossRef]
- Das, S.; Sarkar, S.; Das, M.; Banik, P.; Bhattacharya, S.S. Influence of soil quality factors on capsaicin biosynthesis, pungency, yield, and produce quality of chili: An insight on Csy1, Pun1, and Pun12 signaling responses. Plant Physiol. Biochem. 2021, 166, 427–436. [Google Scholar] [CrossRef]
- Das, S.; Teja, K.C.; Duary, B.; Agrawal, P.K.; Bhattacharya, S.S. Impact of nutrient management, soil type and location on the accumulation of capsaicin in Capsicum chinense (Jacq.): One of the hottest chili in the world. Sci. Hortic. 2016, 213, 354–366. [Google Scholar] [CrossRef]
- Estrada, B.; Pomar, F.; Díaz, J.; Merino, F.; Bernal, M.A. Pungency Level in Fruits of the Padron Pepper with Different Water Supply. Sci. Hortic. 1999, 81, 385–396. [Google Scholar] [CrossRef]
- Gurung, T.; Techawongstien, S.; Suriharn, B.; Techawongstien, S. Stability analysis of yield and capsaicinoids content in chilli (Capsicum spp.) grown across six environments. Euphytica 2012, 187, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Fayos, O.; Ochoa-Alejo, N.; De La Vega, O.M.; Savirón, M.; Orduna, J.; Mallor, C.; Barbero, G.F.; Garcés-Claver, A. Assessment of Capsaicinoid and Capsinoid Accumulation Patterns during Fruit Development in Three Chili Pepper Genotypes (Capsicum spp.) Carrying Pun1 and pAMT Alleles Related to Pungency. J. Agric. Food Chem. 2019, 67, 12219–12227. [Google Scholar] [CrossRef] [PubMed]
- Olguín-Rojas, J.A.; Fayos, O.; Vázquez-León, L.A.; Ferreiro-González, M.; Rodríguez-Jimenes, G.C.; Palma, M.; Garc’s-Claver, A.; Barbero, G.F. Progression of the total and individual capsaicinoids content in the fruits of three different cultivars of capsicum chinense Jacq. Agronomy 2019, 9, 141. [Google Scholar] [CrossRef] [Green Version]
- Saga, K.; Tamura, T. Studies on the changes of some constituents in the fruits in relation to the growth of pepper fruits, especially on development of capsaicin in the fruits. Mem. Fac. Agric. Hokkaido Univ. 1970, 7, 294–300. [Google Scholar]
- Sukrasno, N.; Yeoman, M.M. Phenylpropanoid metabolism during growth and development of Capsicum frutescens fruits. Phytochemistry 1993, 32, 839–844. [Google Scholar] [CrossRef]
- Stewart, C.; Kang, B.C.; Liu, K.; Mazourek, M.; Moore, S.L.; Yoo, E.Y.; Kim, B.D.; Paran, I.; Jahn, M.M. The Pun1 gene for pungency in pepper encodes a putative acyltransferase. Plant J. 2005, 42, 675–688. [Google Scholar] [CrossRef]
- Suzuki, T.; Fujiwake, H.; Iwai, K. Intracellular localization of capsaicin and its analogues, capsaicinoid, in Capsicum fruit 1. Microscopic investigation of the structure of the placenta of Capsicum annuum var. annuum cv. Karayatsubusa. Plant Cell. Physiol. 1980, 21, 839–853. [Google Scholar] [CrossRef]
- Contreras-Padilla, M.; Yahia, E.M. Changes in capsaicinoids during development, maturation, and senescence of chile peppers and relation with peroxidase activity. J. Agric. Food Chem. 1998, 46, 2075–2079. [Google Scholar] [CrossRef]
- Moirangthem, S.S.; Gogoi, S.; Thongbam, P.D.; Ramya, K.T.; Fiyaz, R.A.; Pandey, D.S. Effect of sowing time and crop geometry on the Capsaicinoid content in Bhoot Jolokia (Capsicum chinense Jacq.). J. Food Sci. Technol. 2014, 51, 1974–1981. [Google Scholar] [CrossRef] [Green Version]
- Nandeshwar, V.N.; Bharad, S.G. Effect of Planting Geometry and Fertigation Levels on Growth, Yield and Quality of Chilli. J. Krishi Vigyan 2019, 8, 63–69. [Google Scholar] [CrossRef]
- Topuz, A.; Dincer, C.; Özdemir, K.S.; Feng, H.; Kushad, M. Influence of different drying methods on carotenoids and capsaicinoids of paprika (Cv.; Jalapeno). Food Chem. 2011, 129, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Maurya, V.K.; Gothandam, K.M.; Ranjan, V.; Shakya, A.; Pareek, S. Effect of drying methods (microwave vacuum, freeze, hot air and sun drying) on physical, chemical and nutritional attributes of five pepper (Capsicum annuum var. annuum) cultivars. J. Sci. Food Agric. 2018, 98, 3492–3500. [Google Scholar] [CrossRef] [PubMed]
- Sanatombi, K.; Rajkumari, S. Effect of Processing on Quality of Pepper: A Review. Food Rev. Int. 2019, 36, 626–643. [Google Scholar] [CrossRef]
- Ornelas-Paz, J.J.; Martinez-Burrola, J.M.; Ruiz-Cruz, S.; Santana-Rodriguez, V.; Ibarra-Junquera, V.; Olivas, G.I.; Perez-Martinez, J.D. Effect of cooking on the capsaicinoids and phenolics contents of Mexican peppers. Food Chem. 2010, 119, 1619–1625. [Google Scholar] [CrossRef]
- Moreno-Escamilla, J.O.; Rosa, L.A.; López-Díaz, J.A.; Rodrigo-García, J.; Núñez-Gastélum, J.A.; Alvarez-Parrilla, E. Effect of the smoking process and firewood type in the phytochemical content and antioxidant capacity of red Jalapeño pepper during its transformation to chipotle pepper. Int. Food Res. J. 2015, 76, 654–660. [Google Scholar] [CrossRef]
- Bernal, M.A.; Calderon, A.A.; Pedreño, M.A.; Muñoz, R.; Ros Barceló, A.; Caceres, F.M. Capsaicin oxidation by peroxidase from Capsicum annuum (variety annuum) fruits. J. Agric. Food Chem. 1993, 41, 1041–1044. [Google Scholar] [CrossRef]
- Phimchan, P.; Chanthai, S.; Bosland, P.W.; Techawongstien, S. Impact of drought stress on the accumulation of capsaicinoids in Capsicum cultivars with different initial capsaicinoid levels. HortScience 2012, 47, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Gurung, T.; Techawongstien, S.; Suriharn, B.; Techawongstien, S. Growth, yield and capsaicinoid contents of 14 cultivars of hot pepper (Capsicum spp.) at two elevations of Thailand. Sabrao J. Breed Genet. 2011, 43, 130–143. [Google Scholar]
- Giuffrida, D.; Dugo, P.; Torre, G.; Bignardi, C.; Cavazza, A.; Corradini, C.; Dugo, G. Evaluation of carotenoid and capsaicinoid contents in powder of red chili peppers during one year of storage. Food Res. Int. 2014, 65, 163–170. [Google Scholar] [CrossRef]
- Topuz, A.; Ozdemir, F. Influences of gamma irradiation and storage on the capsaicinoids of sun-dried and dehydrated paprika. Food Chem. 2004, 86, 509–515. [Google Scholar] [CrossRef]
- Iqbal, Q.; Amjad, M.; Asi, M.R.; Ariño, A.; Ziaf, K.; Nawaz, A.; Ahmad, T. Stability of capsaicinoids and antioxidants in dry hot peppers under different packaging and storage temperatures. Foods 2015, 4, 51–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, Q.; Amjad, M.; Asi, M.R.; Nawaz, A.; Khan, S.M.; Ariño, A.; Ahmad, T. Irradiation maintains functional components of dry hot peppers (Capsicum annuum l.) under ambient storage. Foods 2016, 5, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odai, B.T.; Tano-Debrah, K.; Addo, K.K.; Saalia, F.K.; Akyeh, L.M. Effect of gamma radiation and storage at 4 °C on the inactivation of Listeria monocytogenes, Escherichia coli and Salmonella enterica Typhimurium in Legon-18 pepper (Capsicum annuum) powder. Food Qual. Saf. 2019, 3, 265–272. [Google Scholar] [CrossRef]
- Molnár, H.; Bata-Vidács, I.; Baka, E.; Cserhalmi, Z.; Ferenczi, S.; Tömösközi-Farkas, R.; Adànyi, N.; Székács, A. The effect of different decontamination methods on the microbial load, bioactive components, aroma and colour of spice paprika. Food Control 2018, 83, 131–140. [Google Scholar] [CrossRef]
- Duah, A.S.; Silva e Souza, C.; Daood, H.G.; Pék, Z.; Neményi, A.; Helyes, L. Content and response to ɣ-irradiation before over-ripening of capsaicinoid, carotenoid, and tocopherol in new hybrids of spice chili peppers. LWT 2021, 147, 111555. [Google Scholar] [CrossRef]
- Kyung, H.K.; Ramakrishnan, S.R.; Kwon, J.H. Evaluation of capsaicinoid profile and antioxidant properties in dried Korean red pepper (Capsicum annuum L.) as affected by variable dose rates of electron beam and gamma-ray irradiation. J. Food Sci. Technol. 2018, 55, 3902–3910. [Google Scholar] [CrossRef]
- Lee, J.H.; Sung, T.H.; Lee, K.T.; Kim, M.R. Effect of gamma-irradiation on color, pungency, and volatiles of Korean red pepper powder. J. Food Sci. 2004, 69, C585–C592. [Google Scholar] [CrossRef]
- Kwon, J.H.; Byun, M.W.; Cho, H.O. Effect of gamma irradiation on the sterilization of red pepper powder. J. Korean Soc. Food Sci. Nutr. 1984, 13, 188–192. [Google Scholar]
- Balakrishnan, N.; Yusop, S.M.; Rahman, I.A.; Dauqan, E.; Abdullah, A. Efficacy of Gamma Irradiation in Improving the Microbial and Physical Quality Properties of Dried Chillies (Capsicum annuum L.): A Review. Foods 2022, 11, 91. [Google Scholar] [CrossRef]
- Jung, K.; Song, B.S.; Kim, M.J.; Moon, B.-G.; Go, S.M.; Kim, J.K.; Lee, Y.J.; Park, J.H. Effect of X-ray, gamma ray, and electron beam irradiation on the hygienic and physicochemical qualities of red pepper powder. LWT 2015, 63, 846–851. [Google Scholar] [CrossRef]
- Cheok, C.Y.; Sobhi, B.; Mohd Adzahan, N.; Bakar, J.; Abdul Rahman, R.; Ab Karim, M.S.; Ghazali, Z. Physicochemical properties and volatile profile of chili shrimp paste as affected by irradiation and heat. Food Chem. 2017, 216, 10–18. [Google Scholar] [CrossRef]
- Veloso, J.; Prego, C.; Varela, M.M.; Carballeira, R.; Bernal, A.; Merino, F.; Díaz, J. Properties of capsaicinoids for the control of fungi and oomycetes pathogenic to pepper. Plant Biol. 2014, 16, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Aranda, F.J.; Villalín, J.; Gómez-Fernández, J.C. Capsaicin affects the structure and phase organization of phospholipid membranes. Biochim. Biophys. Acta 1995, 1234, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, H. Biphasic membrane effects of capsaicin, an active component in Capsicum species. J. Ethnopharmacol. 2001, 75, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Kurita, S.; Kitagawa, E.; Kim, C.H.; Momose, Y.; Iwahashi, H. Studies on the antimicrobial mechanisms of capsaicin using yeast DNA microarray. Biosci. Biotechnol. Biochem. 2002, 66, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Jamaica, L.M.; Villordo-Pineda, E.; González-Chavira, M.M.; Guevara-González, R.G.; Medina-Ramos, G. Effect of Fragmented DNA From Plant Pathogens the Protection Against Wilt and Root Rot of Capsicum annuum L. plants. Front. Plant Sci. 2021, 11, 581891. [Google Scholar] [CrossRef] [PubMed]
- Oljira, T.; Berta, S. Isolation and characterization of wilt-causing pathogens of local growing pepper (Capsicum annuum L.) in Gurage Zone, Ethiopia. Int. J. Agron. 2020, 2020, 6638683. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Reagan, K.M.; Machnicki, N.J.; Carlo, T.A.; Haak, D.C.; Peñaloza, A.L.C.; Levey, D.J. Evolutionary ecology of pungency in wild chilies. Proc. Natl. Acad. Sci. USA 2008, 105, 11808–11811. [Google Scholar] [CrossRef] [Green Version]
- Tahboub, M.B.; Sanogo, S.; Bosland, P.W.; Murray, L. Heat Level in Chile Pepper in Relation to Root and Fruit Infection by Phytophthora capsici. HortScience 2008, 43, 1846–1851. [Google Scholar] [CrossRef] [Green Version]
- Padilha, H.K.M.; Madruga, N.A.; Aranha, B.C.; Hoffmann, J.F.; Crizel, R.L.; Barbieri, R.L.; Chaves, F.C. Defense responses of Capsicum spp. genotypes to post-harvest Colletotrichum sp. inoculation. Phytoparasitica 2019, 47, 557–573. [Google Scholar] [CrossRef]
- Saini, T.J.; Tiwari, A.; Yeole, M.; Gupta, S. Effect of pungency levels of Capsicum spp. fruits on tolerance to anthracnose. Physiol. Mol. Plant Pathol. 2021, 116, 101720. [Google Scholar] [CrossRef]
- Kollia, E.; Proestos, C.; Zoumpoulakis, P.; Markaki, P. Capsaicin, an inhibitor of Ochratoxin A production by Aspergillus section Nigri strains in grapes (Vitis vinifera L.). Food Addit. Contam. 2019, 36, 1709–1721. [Google Scholar] [CrossRef] [PubMed]
- Buitimea-Cantúa, G.V.; Buitimea-Cantúa, N.E.; Rocha-Pizaña, M.d.R.; Hernández-Morales, A.; Magaña-Barajas, E.; Molina-Torres, J. Inhibitory effect of Capsicum chinense and Piper nigrum fruits, capsaicin and piperine on aflatoxins production in Aspergillus parasiticus by downregulating the expression of aflD, aflM, aflR, and aflS genes of aflatoxins biosynthetic pathway. J. Environ. Sci. Health B 2020, 55, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Hernandez, J.A.; Solano-Alvarez, N.; Feregrino-Perez, A.A.; Rico-Rodriguez, M.-A.; Torres-Pacheco, I.; Zavala-Gomez, C.-E.; Guevara-Gonzalez, R.-G. In vitro and in vivo antimicrobial activity of a synthetic capsaicinoid oleoresin against Fusarium oxysporum, Phytophthora capsici, Clavibacter michiganensis and Pseudomonas syringae. J. Plant Pathol. 2022, 104, 699–710. [Google Scholar] [CrossRef]
- Zanotto, A.W. Emprego de Compostos Amídicos para o Controle de Penicillium expansum em Maçã. Bachelor’s Thesis, Universidade Tecnológica Federal do Paraná, Francisco Beltrão, Brazil, 2016. [Google Scholar]
- Vázquez-Espinosa, M.; Fayos, O.; González-de-Peredo, A.V.; Espada-Bellido, E.; Ferreiro-González, M.; Palma, M.; Garcès-Claver, A.; Barbero, G.F. Content of Capsaicinoids and Capsiate in “Filius” Pepper Varieties as Affected by Ripening. Plants 2020, 9, 1222. [Google Scholar] [CrossRef] [PubMed]
- García, T.; Veloso, J.; Díaz, J. Properties of vanillyl nonanoate for protection of pepper plants against Phytophthora capsici and Botrytis cinerea. Eur. J. Plant Pathol. 2018, 150, 1091–1101. [Google Scholar] [CrossRef]
- Vázquez-Fuentes, S.; Pelagio-Flores, R.; López-Bucio, J.; Torres-Gavilán, A.; Campos-García, J.; Cruz, H.R.; López-Bucio, J.S. N-vanillyl-octanamide represses growth of fungal phytopathogens in vitro and confers postharvest protection in tomato and avocado fruits against fungal-induced decay. Protoplasma 2021, 258, 729–741. [Google Scholar] [CrossRef]
- Akyuz, L.; Kaya, M.; Mujtaba, M.; Ilk, S.; Sargin, I.; Salaberria, A.M.; Labidi, J.; Cakmak, Y.S.; Islek, C. Supplementing capsaicin with chitosan-based films enhanced the anti-quorum sensing, antimicrobial, antioxidant, transparency, elasticity and hydrophobicity. Int. J. Biol. Macromol. 2018, 115, 438–446. [Google Scholar] [CrossRef]
- Pagnossa, J.P.; Rocchetti, G.; de Abreu Martins, H.H.; Bezerra, J.D.; Batiha, G.E.-S.; El-Masry, E.A.; Cocconcelli, P.S.; Santos, C.; Lucini, L.; Piccoli, R.H. Morphological and metabolomics impact of sublethal doses of natural compounds and its nanoemulsions in Bacillus cereus. Food Res. Int. 2021, 149, 110658. [Google Scholar] [CrossRef]
- Pagnossa, J.P.; Rocchetti, G.; Bezerra, J.D.; Batiha, G.E.-S.; El-Masry, E.A.; Mahmoud, M.H.; Alsayegh, A.A.; Mashraqi, A.; Cocconcelli, P.S.; Santos, C.; et al. Untargeted Metabolomics Approach of Cross-Adaptation in Salmonella Enterica Induced by Major Compounds of Essential Oils. Front. Microbiol. 2022, 13, 769110. [Google Scholar] [CrossRef]
- Hernández-Téllez, C.N.; Luque-Alcaraz, A.G.; Núñez-Mexía, S.A.; Cortez-Rocha, M.O.; Lizardi-Mendoza, J.; Rosas-Burgos, E.C.; Rosas-durazo, A.d.J.; Parra-Vergara, N.V.; Plascencia-Jatomea, M. Relationship between the Antifungal Activity of Chitosan–Capsaicin Nanoparticles and the Oxidative Stress Response on Aspergillus parasiticus. Polymers 2022, 14, 2774. [Google Scholar] [CrossRef] [PubMed]
- Nidiry, E.S.J. Evaluation of Antifungal Activity of Some Plant Extractives. Indian J. Pharm. Sci. 2022, 84, 224–227. [Google Scholar] [CrossRef]
- Soumya, S.L.; Nair, B.R. Antifungal efficacy of Capsicum frutescens L. extracts against some prevalent fungal strains associated with groundnut storage. J. Agric. Technol. 2012, 8, 739–750. [Google Scholar]
- Singh, H.; Fairs, G.; Syarhabil, M. Anti-fungal activity of Capsicum frutescence and Zingiber officinale against key post-harvest pathogens in Citrus. In Proceedings of the International Conference on Biomedical Engineering and Technology, Kuala Lumpur, Malaysia, 4–5 June 2011; Volume 11, pp. 1–6. [Google Scholar]
- Singh, T.; Chittenden, C. In-vitro antifungal activity of chilli extracts in combination with Lactobacillus casei against common sapstain fungi. Int. Biodeterior. Biodegrad. 2008, 62, 364–367. [Google Scholar] [CrossRef]
- Fieira, C.; Oliveira, F.; Calegari, R.P.; Machado, A.; Coelho, A.R. In vitro and in vivo antifungal activity of natural inhibitors against Penicillium expansum. Food Sci. Technol. 2013, 33, 40–46. [Google Scholar] [CrossRef]
- Kraikruan, W.; Sangchote, S.; Sukprakarn, S. Effect of Capsaicin on Germination of Colletotrichum capsici Conidia. Nat. Sci. 2008, 42, 417–422. [Google Scholar]
- Xing, F.; Cheng, G.; Yi, K. Study on the antimicrobial activities of the capsaicin microcapsules. J. Appl. Polym. Sci. 2006, 102, 1318–1321. [Google Scholar] [CrossRef]
- Romero-Luna, H.E.; Colina, J.; Guzmán-Rodríguez, L.; Sierra-Carmona, C.G.; Farías-Campomanes, Á.M.; García-Pinilla, S.; González-Tijera, M.M.; Malagón-Alvira, K.O.; Peredo-Lovillo, A. Capsicum fruits as functional ingredients with antimicrobial activity: An emphasis on mechanisms of action. J. Food Sci. Technol. 2022, 4, 1–11. [Google Scholar] [CrossRef]
- Cuadrado, C.J.L.; Pinillos, E.O.; Tito, R.; Mirones, C.S.; Gamarra Mendoza, N.N. Insecticidal properties of capsaicinoids and glucosinolates extracted from Capsicum chinense and Tropaeolum tuberosum. Insects 2019, 10, 132. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Yang, M.; Shi, R.; Ye, M. Insecticidal activity of natural capsaicinoids against several agricultural insects. Nat. Prod. Commun. 2019, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Soujanya, P.L.; Sekhar, J.C.; Kumar, P. Efficacy of plant extracts against rice weevil Sitophilus oryzae (L.) in stored maize. Indian J. Entomol. 2016, 78, 342–345. [Google Scholar] [CrossRef]
- Chowański, S.; Adamski, Z.; Marciniak, P.; Rosiński, G.; Büyükgüzel, E.; Büyükgüzel, K.; Falabella, P.; Scrano, L.; Ventrella, E.; Lelario, F.; et al. A review of bioinsecticidal activity of Solanaceae alkaloids. Toxins 2016, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, J.; Tęgowska, E. Opposite effect of capsaicin and capsazepine on behavioral thermoregulation in insects. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2011, 197, 1021–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelson, J.V.; Duthie, J.; Roberts, W. Toxicity of biorational insecticides: Activity against the green peach aphid, Myzus persicae (Sulzer). Pest Manag. Sci. 2002, 58, 255–260. [Google Scholar] [CrossRef]
- Hans, L.; Saxena, S. Plant Bioprospecting for Biopesticides and Bioinsecticides. In Bioprospecting of Plant Biodiversity for Industrial Molecules, 1st ed.; Upadhyay, S.K., Singh, S.P., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2021; pp. 335–344. [Google Scholar]
- McCarty, M.F.; DiNicolantonio, J.J.; O’Keefe, J.H. Capsaicin may have important potential for promoting vascular and metabolic health. Open Heart 2015, 2, e000262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehie, M.; Kumaria, S.; Tandon, P.; Ramchiary, N. Biotechnological advances on in vitro capsaicinoids biosynthesis in Capsicum: A review. Phytochem. Rev. 2014, 14, 189–201. [Google Scholar] [CrossRef]
- Petrushenko, M.O.; Petrushenko, E.A.; Lukyanetz, E.A. Activation and Desensitization of TRPV1 Channels under the Influence of Capsaicin. Neurophysiology 2020, 52, 256–260. [Google Scholar] [CrossRef]
- Batiha, G.E.S.; Alqahtani, A.; Ojo, O.A.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A.; et al. Biological properties, bioactive constituents, and pharmacokinetics of some Capsicum spp. and capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179. [Google Scholar] [CrossRef]
- Lin, M.H.; Lee, Y.H.; Cheng, H.L.; Chen, H.Y.; Jhuang, F.H.; Chueh, P.J. Capsaicin Inhibits Multiple Bladder Cancer Cell Phenotypes by Inhibiting Tumor-Associated NADH Oxidase (tNOX) and Sirtuin1 (SIRT1). Molecules 2016, 21, 849. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.R.; Richbart, S.D.; Merritt, J.C.; Brown, K.C.; Denning, K.L.; Tirona, M.T.; Valentovic, M.A.; Miles, S.L.; Dasgupta, P. Capsaicinoids: Multiple effects on angiogenesis, invasion and metastasis in human cancers. Biomed. Pharmacother. 2019, 118, 109317. [Google Scholar] [CrossRef]
- Jin, T.; Wu, H.; Wang, Y.; Peng, H. Capsaicin induces immunogenic cell death in human osteosarcoma cells. Exp. Ther. Med. 2016, 2, 765–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantrell, C.L.; Jarret, R.L. Bulk process for enrichment of capsinoids from Capsicum fruit. Processes 2022, 10, 305. [Google Scholar] [CrossRef]
- Stewart, C., Jr.; Mazourek, M.; Stellari, G.M.; O’Connell, M.; Jahn, M. Genetic control of pungency in C. chinense via the Pun1 locus. J. Exp. Bot. 2007, 58, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Park, M.; Lee, D.-G.; Lim, J.-H.; Jung, J.-W.; Kang, B.-C. Breeding Capsicum chinense lines with high levels of capsaicinoids and capsinoids in the fruit. Agriculture 2021, 11, 819. [Google Scholar] [CrossRef]
- Muratovska, N.; Grey, C.; Carlquist, M. Engineering Saccharomyces cerevisiae for production of the capsaicinoid nonivamide. Microb. Cell Fact. 2022, 21, 106. [Google Scholar] [CrossRef]
- Muratovska, N.; Silva, P.; Pozdniakova, T.; Pereira, H.; Grey, C.; Johansson, B.; Carlquist, M. Towards engineered yeast as production platform for capsaicinoids. Biotechnol. Adv. 2022, 59, 107989. [Google Scholar] [CrossRef]
- Lentmaier, C. Characterization of capsaicinoid production in recombinant Saccharomyces cerevisiae. In Degree Project in Biomedical Laboratory Science; Faculty of Health and Society, Department of Biomedical Science, Malmö University: Malmö, Sweden, 2018. [Google Scholar]
- Kashani, A.H. Evaluating E. coli as a Chassis for the Production of Capsaicinoids and Capsinoids. Master’s Thesis, The University of British Columbia, Vancouver, BC, Canada, 2021. [Google Scholar]
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef]
- Van Eck, J. Genome editing and plant transformation of solanaceous food crops. Curr. Opin. Biotechnol. 2018, 49, 35–41. [Google Scholar] [CrossRef]
- Lozada, D.N.; Bosland, P.W.; Barchenger, D.W.; Haghshenas-Jaryani, M.; Sanogo, S.; Walker, S. Chile pepper (Capsicum) breeding and improvement in the “multi-omics” era. Front. Plant Sci. 2022, 13, 879182. [Google Scholar] [CrossRef]
- Park, S.-il.; Kim, H.-B.; Jeon, H.-J.; Kim, H. Agrobacterium-mediated Capsicum annuum gene editing in two cultivars, hot pepper CM334 and bell pepper Dempsey. Int. J. Mol. Sci. 2021, 22, 3921. [Google Scholar] [CrossRef]
- Kim, H.; Choi, J.; Won, K.H. A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum. BMC Plant Biol. 2020, 20, 449. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhou, Z.; Liang, L.; Song, Z.; Hu, Y.; Cui, J.; Chen, W.; Hu, K.; Cheng, J. Genome-wide identification and analysis of highly specific CRISPR/Cas9 editing sites in pepper (Capsicum annuum L.). PLoS ONE 2020, 15, e0244515. [Google Scholar] [CrossRef] [PubMed]
- Won, K.-H.; Park, S.-il.; Choi, J.; Kim, H.H.; Kang, B.-C.; Kim, H. A reliable regeneration method in genome-editable bell pepper ‘Dempsey’. Horticulturae 2021, 7, 317. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, J.; Sepúlveda, M.; Gallardo, V.; Cayún, Y.; Santander, C.; Ruíz, A.; Reyes, M.; Santos, C.; Cornejo, P.; Lima, N.; et al. Antifungal Potential of Capsaicinoids and Capsinoids from the Capsicum Genus for the Safeguarding of Agrifood Production: Advantages and Limitations for Environmental Health. Microorganisms 2022, 10, 2387. https://doi.org/10.3390/microorganisms10122387
Costa J, Sepúlveda M, Gallardo V, Cayún Y, Santander C, Ruíz A, Reyes M, Santos C, Cornejo P, Lima N, et al. Antifungal Potential of Capsaicinoids and Capsinoids from the Capsicum Genus for the Safeguarding of Agrifood Production: Advantages and Limitations for Environmental Health. Microorganisms. 2022; 10(12):2387. https://doi.org/10.3390/microorganisms10122387
Chicago/Turabian StyleCosta, Jéssica, Marcela Sepúlveda, Víctor Gallardo, Yasna Cayún, Christian Santander, Antonieta Ruíz, Marjorie Reyes, Carla Santos, Pablo Cornejo, Nelson Lima, and et al. 2022. "Antifungal Potential of Capsaicinoids and Capsinoids from the Capsicum Genus for the Safeguarding of Agrifood Production: Advantages and Limitations for Environmental Health" Microorganisms 10, no. 12: 2387. https://doi.org/10.3390/microorganisms10122387
APA StyleCosta, J., Sepúlveda, M., Gallardo, V., Cayún, Y., Santander, C., Ruíz, A., Reyes, M., Santos, C., Cornejo, P., Lima, N., & Santos, C. (2022). Antifungal Potential of Capsaicinoids and Capsinoids from the Capsicum Genus for the Safeguarding of Agrifood Production: Advantages and Limitations for Environmental Health. Microorganisms, 10(12), 2387. https://doi.org/10.3390/microorganisms10122387