
Enterococcus casseliflavus KB1733 Isolated from a Traditional Japanese Pickle Induces Interferon-Lambda Production in Human Intestinal Epithelial Cells

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. To Screen for the LAB Strain Capable of Inducing the Gene Expression of IFN-λ3
2.1.1. Strains, Growth Conditions, and Sample Preparation
2.1.2. Cells
2.1.3. Cell Culture with Samples and Reporter Assay for IFN-λ3
2.2. To Assess the Ability of Enterococcus casseliflavus KB1733 to Produce IFN-λs and Induce Expressions of IFN-λ Genes and ISGs
2.2.1. Strains, Growth Conditions, and Sample Preparation
2.2.2. Cells
2.2.3. Cell Culture with Samples
2.2.4. Assay for Production of IFN-λ1, -λ2, and -λ3
2.2.5. Assay for Expressions of IFN-λ Genes and ISGs
2.3. Statistical Analysis
3. Results
3.1. To Screen for the LAB Strain Capable of Inducing the Gene Expression of IFN-λ3
3.2. To Assess the Ability of Enterococcus casseliflavus KB1733 to Induce IFN-λ Production and Expressions of IFN-λ Genes and ISGs
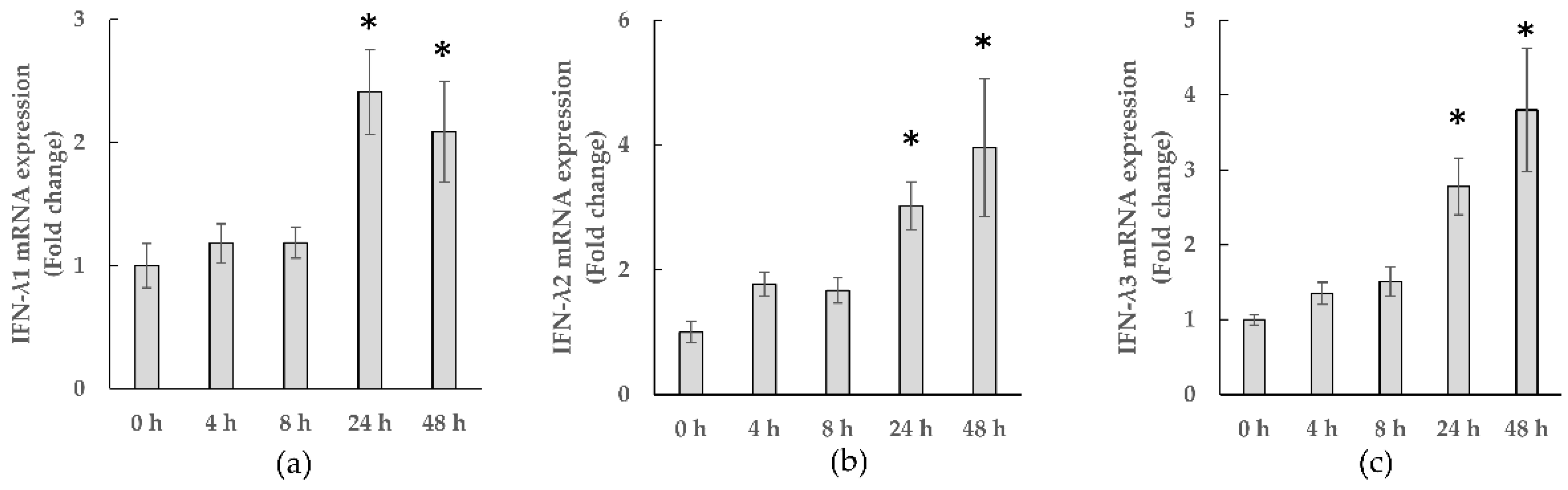
3.2.1. Gene Expressions of IFN-λs
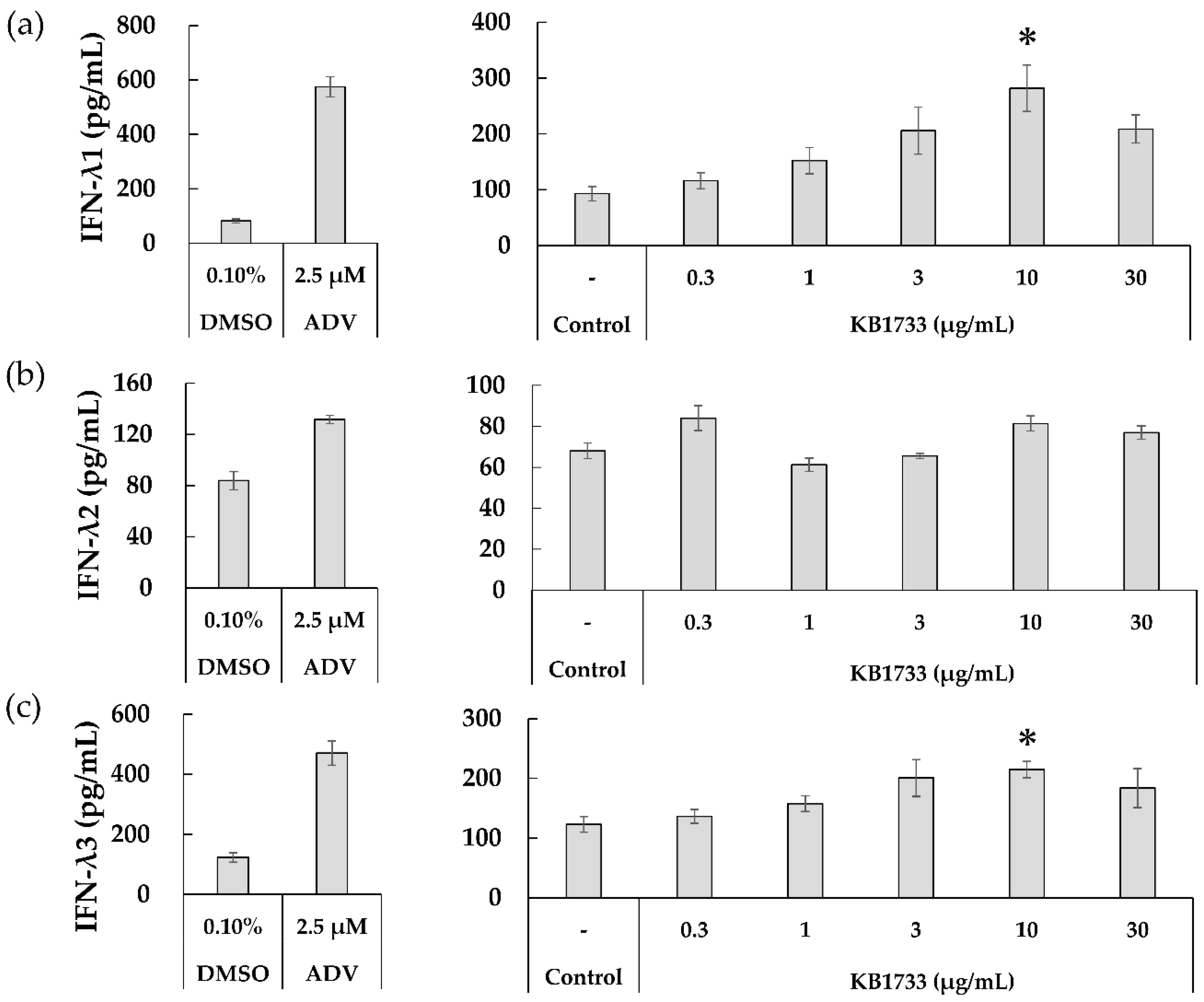
3.2.2. Production of IFN-λ1, -λ2, and -λ3
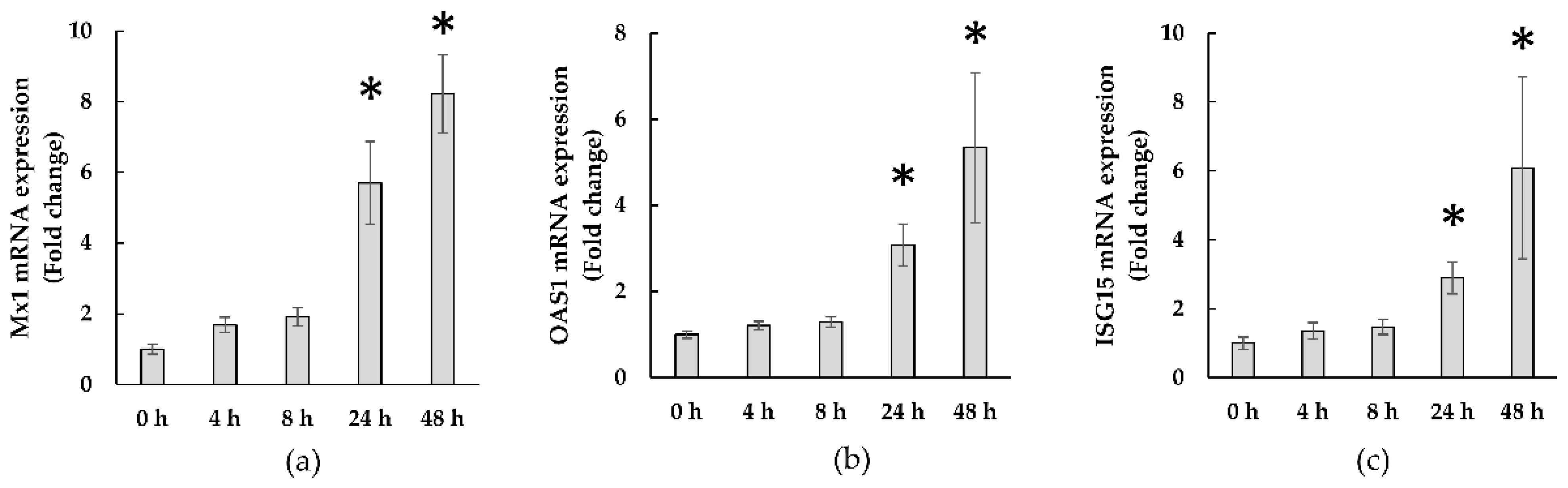
3.2.3. Expressions of ISGs
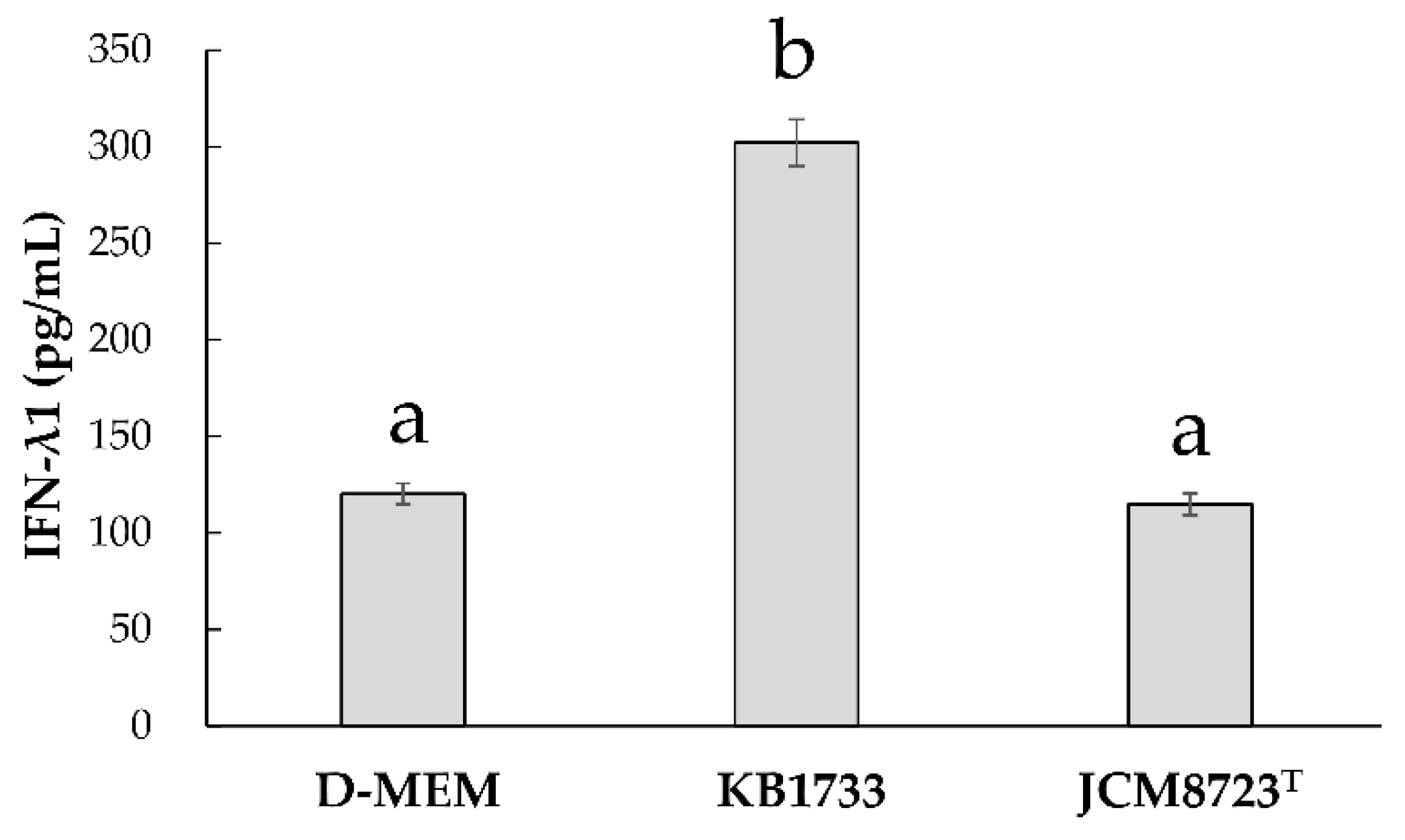
3.2.4. Strain Specificity for IFN-λ1 Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joint FAO/WHO Working Group. Guidelines for the Evaluation of Probiotics in Food: Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization of the United Nations: London, ON, Canada, 2002. [Google Scholar]
- Mulaw, G.; Tessema, T.S.; Muleta, D.; Tesfaye, A. In Vitro Evaluation of Probiotic Properties of Lactic Acid Bacteria Isolated from Some Traditionally Fermented Ethiopian Food Products. Int. J. Microbiol. 2019, 2019, 7179514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, C.L.; Thorsen, L.; Schwan, R.F.; Jespersen, L. Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiol. 2013, 36, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, E.; Suzuki, S.; Fukui, Y.; Yajima, N. Cell-bound exopolysaccharides of Lactobacillus brevis KB290 enhance cytotoxic activity of mouse splenocytes. J. Appl. Microbiol. 2015, 118, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Fukui, Y.; Sasaki, E.; Fuke, N.; Nakai, Y.; Ishijima, T.; Abe, K.; Yajima, N. Effect of Lactobacillus brevis KB290 on the cell-mediated cytotoxic activity of mouse splenocytes: A DNA microarray analysis. Br. J. Nutr. 2013, 110, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- Villena, L.; Aso, H.; Rutten, V.P.M.G.; Takahashi, H.; van Eden, W.; Kitazawa, H. Immunobiotics for the Bovine Host: Their Interaction with Intestinal Epithelial Cells and Their Effect on Antiviral Immunity. Front. Immunol. 2018, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Waki, N.; Yajima, N.; Suganuma, H.; Buddle, B.M.; Luo, D.; Heiser, A.; Zheng, T. Oral administration of Lactobacillus brevis KB290 to mice alleviates clinical symptoms following influenza virus infection. Lett. Appl. Microbiol. 2014, 58, 87–93. [Google Scholar] [CrossRef]
- Waki, N.; Matsumoto, M.; Fukui, Y.; Suganuma, H. Effects of probiotic Lactobacillus brevis KB290 on incidence of influenza infection among schoolchildren: An open-label pilot study. Lett. Appl. Microbiol. 2014, 59, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Russell, W.M.; Douglas-Escobar, M.; Hauser, N.; Lopez, M.; Neu, J. Live and Heat-Killed Lactobacillus rhamnosus GG: Effects on Proinflammatory and Anti-Inflammatory Cytokines/Chemokines in Gastrostomy-Fed Infant Rats. Pediatr. Res. 2009, 66, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Sang, L.X.; Chang, B.; Wang, B.Y.; Liu, W.X.; Jiang, M. Live and heat-killed probiotic: Effects on chronic experimental colitis induced by dextran sulfate sodium (DSS) in rats. Int. J. Clin. Exp. Med. 2015, 8, 20072–20078. [Google Scholar]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [Green Version]
- Satomi, S.; Waki, N.; Arakawa, C.; Fujisawa, K.; Suzuki, S.; Suganuma, H. Effects of Heat-Killed Levilactobacillus brevis KB290 in Combination with β-Carotene on Influenza Virus Infection in Healthy Adults: A Randomized Controlled Trial. Nutrients 2021, 13, 3039. [Google Scholar] [CrossRef] [PubMed]
- Hermant, P.; Michiels, T. Interferon-λ in the context of viral infections: Production, response and therapeutic implications. J. Innate Immun. 2014, 6, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Schnepf, D.; Staeheli, P. Interferon-lambda orchestrates innate and adaptive mucosal immune responses. Nat. Rev. Immunol. 2019, 19, 614–625. [Google Scholar] [CrossRef]
- Sommereyns, C.; Paul, S.; Staeheli, P.; Michiels, T. IFN-lambda (IFN-lambda) is expressed in a tissue-dependent fashion and primarily acts on epithelial cells in vivo. PLoS Pathog. 2008, 4, e10000017. [Google Scholar] [CrossRef]
- Kotenko, S.V.; Gallagher, G.; Baurin, V.V.; Lewis-Antes, A.; Shen, M.; Shah, N.K.; Langer, J.A.; Sheikh, F.; Dickensheets, H.; Donnelly, R.P. IFN-lambdas mediate antiviral protection through a distinct class II cytokine receptor complex. Nat. Immunol. 2003, 4, 69–77. [Google Scholar] [CrossRef]
- Sheppard, P.; Kindsvogel, W.; Xu, W.; Henderson, K.; Schlutsmeyer, S.; Whitmore, T.E.; Kuestner, R.; Garrigues, U.; Birks, C.; Roraback, J.; et al. IL-28, IL-29 and their class II cytokine receptor IL-28R. Nat. Immunol. 2003, 4, 63–68. [Google Scholar] [CrossRef]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef]
- Hornung, V.; Hartmann, R.; Ablasser, A.; Hopfner, K.P. OAS proteins and cGAS: Unifying concepts in sensing and responding to cytosolic nucleic acids. Nat. Rev. Immunol. 2014, 14, 521–528. [Google Scholar] [CrossRef]
- Cardoso, N.P.; Mansilla, F.C.; Benedetti, E.; Turco, C.S.; Barone, L.J.; Iserte, J.A.; Soria, I.; Baumeister, E.; Capozzo, A.V. Bovine Interferon Lambda Is a Potent Antiviral Against SARS-CoV-2 Infection in vitro. Front. Vet. Sci. 2020, 7, 603622. [Google Scholar] [CrossRef]
- Nice, T.J.; Baldridge, M.T.; McCune, B.T.; Norman, J.M.; Lazear, H.M.; Artyomov, M.; Diamond, M.S.; Virgin, H.W. Interferon-λ cures persistent murine norovirus infection in the absence of adaptive immunity. Science 2015, 347, 269–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, K.; Asano, M.; Matsumoto, A.; Sugiyama, M.; Nishida, N.; Tanaka, E.; Inoue, T.; Sakamoto, M.; Enomoto, N.; Shirasaki, T.; et al. Induction of IFN-λ3 as an additional effect of nucleotide, not nucleoside, analogues: A new potential target for HBV infection. Gut 2018, 67, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Murayama, A.; Shiina, M.; Aly, H.H.; Iwamoto, M.; Tsukuda, S.; Watashi, K.; Tanaka, T.; Moriishi, K.; Nishitsuji, H.; et al. Anti-viral effects of interferon-λ3 on hepatitis B virus infection in cell culture. Hepatol. Res. 2020, 50, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Kang, J.H.; Nagashima, S.; Matsui, T.; Karino, Y.; Yamamoto, Y.; Atarashi, T.; Oohara, M.; Uebayashi, M.; Sakata, H.; et al. IFN-λ3 as a host immune response in acute hepatitis E virus infection. Cytokine 2020, 125, 154816. [Google Scholar] [CrossRef] [PubMed]
- Jounai, K.; Ikado, K.; Sugimura, T.; Ano, Y.; Braun, J.; Fujiwara, D. Spherical lactic acid bacteria activate plasmacytoid dendritic cells immunomodulatory function via TLR9-dependent crosstalk with myeloid dendritic cells. PLoS ONE 2012, 7, e32588. [Google Scholar] [CrossRef]
- Indo, Y.; Kitahara, S.; Tomokiyo, M.; Araki, S.; Islam, M.A.; Zhou, B.; Albarracin, L.; Miyazaki, A.; Ikeda-Ohtsubo, W.; Nochi, T.; et al. Ligilactobacillus salivarius Strains Isolated From the Porcine Gut Modulate Innate Immune Responses in Epithelial Cells and Improve Protection Against Intestinal Viral-Bacterial Superinfection. Front. Immunol. 2021, 12, 652923. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Kimoto-Nira, H.; Suganuma, H.; Suzuki, C.; Saito, T.; Yajima, N. Cellular fatty acid composition and exopolysaccharide contribute to bile tolerance in Lactobacillus brevis strains isolated from fermented Japanese pickles. Can. J. Microbiol. 2014, 60, 183–191. [Google Scholar] [CrossRef]
- Sugiyama, M.; Tanaka, Y.; Wakita, T.; Nakanishi, M.; Mizokami, M. Genetic Variation of the IL-28B Promoter Affecting Gene Expression. PLoS ONE 2011, 6, e26620. [Google Scholar] [CrossRef]
- Sugiyama, M.; Kimura, T.; Naito, S.; Mukaide, M.; Shinauchi, T.; Ueno, M.; Ito, K.; Murata, K.; Mizokami, M. Development of specific and quantitative real-time detection PCR and immunoassays for λ3-interferon. Hepatol. Res. 2012, 42, 1089–1099. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented Foods as a Dietary Source of Live Organisms. Front. Microbiol. 2018, 24, 1785. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, Y.; Izumo, T.; Izumi, F.; Maekawa, T.; Shibata, H.; Nakano, A.; Kishi, A.; Akatani, K.; Kiso, Y. Antiallergic Effects of Lactobacillus pentosus strain S-PT84 mediated by modulation of Th1/Th2 immunobalance and induction of IL-10 production. Int. Arch. Allergy Immunol. 2008, 145, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Dellgren, C.; Gad, H.H.; Hamming, O.J.; Melchjorsen, J.; Hartmann, R. Human interferon-lambda3 is a potent member of the type III interferon family. Genes Immun. 2009, 10, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Nagino, T.; Hoshino, G.; Ushida, K. Nucleic acids of Enterococcus faecalis strain EC-12 are potent Toll-like receptor 7 and 9 ligands inducing interleukin-12 production from murine splenocytes and murine macrophage cell line J774. 1. FEMS Immunol. Med. Microbiol. 2011, 61, 94–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macho Fernandez, E.; Valenti, V.; Rockel, C.; Hermann, C.; Pot, B.; Boneca, I.G.; Grangette, C. Anti-inflammatory capacity of selected lactobacilli in experimental colitis is driven by NOD2-mediated recognition of a specific peptidoglycan-derived muropeptide. Gut 2011, 60, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- EC. Directive 2000/54/EC of the European Parliament and of the council of 18 September 2000 on the protection of workers from risks related to exposure to biological agents at work. Off. J. Eur. Union L. 2000, 262, 21–45. [Google Scholar]
- Jahan, M.; Zhanel, G.G.; Sparling, R.; Holley, R.A. Horizontal transfer of antibiotic resistance from Enterococcus faecium of fermented meat origin to clinical isolates of E. faecium and Enterococcus faecalis. Int. J. Food Microbiol. 2015, 199, 78–85. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′→3′) | |
|---|---|---|
| Human IL-29 (IFN-λ1) | F | GCCTCCTCACGCGAGACCTC |
| R | GGAGTAGGGCTCAGCGCATA | |
| Human IL-28A (IFN-λ2) | F | TCTGGAGGCCACCGCTGACA |
| R | TGGGCTGAGGCTGGATACAG | |
| Human IL-28B (IFN-λ3) | F | TGGCCCTGACGCTGAAGGTT |
| R | CGTGGGCTGAGGCTGGATAC | |
| Homo sapiens MX dynamin like GTPase 1 (MX1) | F | TACCAGACTCCGACACGAGTTCC |
| R | GATTTGCTGTTTCACGATTGTCTCA | |
| Homo sapiens 2′-5′-oligoadenylate synthetase 1 (OAS1) | F | AGAGCCTCATCCGCCTAGTCAA |
| R | GCTCCCAAGCATAGACCGTCA | |
| Interferon stimulated gene 15 (ISG15) | F | TGGACAAATGCGACGAACCTC |
| R | CTGCGGCCCTTGTTATTCCTC | |
| Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) | F | TGGTGAAGACGCCAGTGGA |
| R | GCACCGTCAAGGCTGAGAAC | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satomi, S.; Kokubu, D.; Inoue, T.; Sugiyama, M.; Mizokami, M.; Suzuki, S.; Murata, K. Enterococcus casseliflavus KB1733 Isolated from a Traditional Japanese Pickle Induces Interferon-Lambda Production in Human Intestinal Epithelial Cells. Microorganisms 2022, 10, 827. https://doi.org/10.3390/microorganisms10040827
Satomi S, Kokubu D, Inoue T, Sugiyama M, Mizokami M, Suzuki S, Murata K. Enterococcus casseliflavus KB1733 Isolated from a Traditional Japanese Pickle Induces Interferon-Lambda Production in Human Intestinal Epithelial Cells. Microorganisms. 2022; 10(4):827. https://doi.org/10.3390/microorganisms10040827
Chicago/Turabian StyleSatomi, Shohei, Daichi Kokubu, Takuro Inoue, Masaya Sugiyama, Masashi Mizokami, Shigenori Suzuki, and Kazumoto Murata. 2022. "Enterococcus casseliflavus KB1733 Isolated from a Traditional Japanese Pickle Induces Interferon-Lambda Production in Human Intestinal Epithelial Cells" Microorganisms 10, no. 4: 827. https://doi.org/10.3390/microorganisms10040827
APA StyleSatomi, S., Kokubu, D., Inoue, T., Sugiyama, M., Mizokami, M., Suzuki, S., & Murata, K. (2022). Enterococcus casseliflavus KB1733 Isolated from a Traditional Japanese Pickle Induces Interferon-Lambda Production in Human Intestinal Epithelial Cells. Microorganisms, 10(4), 827. https://doi.org/10.3390/microorganisms10040827