
Comparative Genomics of Acetic Acid Bacteria within the Genus Bombella in Light of Beehive Habitat Adaptation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Ecology
2.2. Genome Sequencing, Assembly and Annotation
2.3. 16S rRNA Gene Sequence, Average Nucleotide Identity (ANI) and In Silico DNA-DNA Hybridization (DDH) Values
2.4. Comparative Genomic Analysis
2.5. Analysis of Antimicrobial Resistance
2.6. Growth Experiments
3. Results
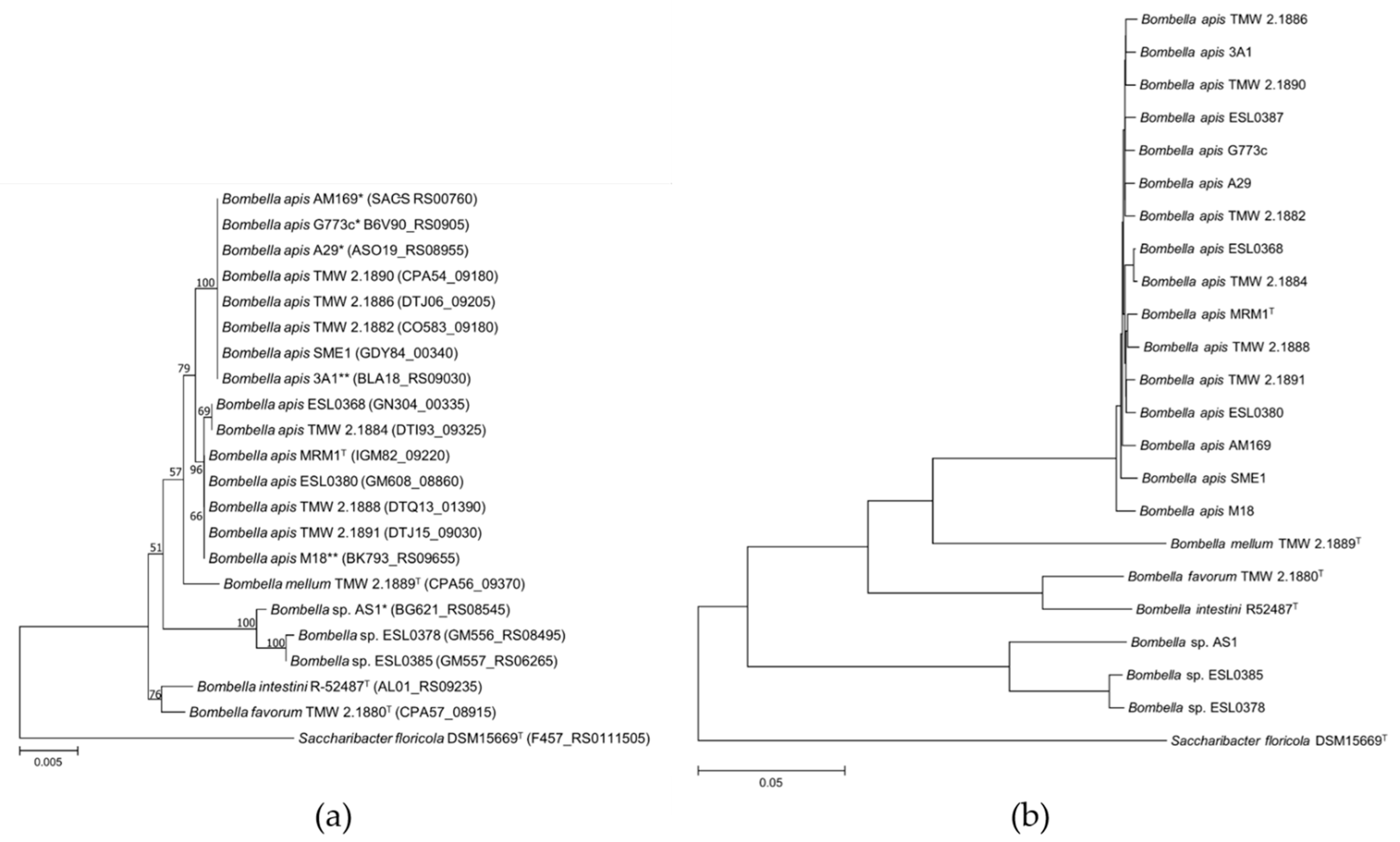
3.1. Phylogeny, Current Taxonomy and Reclassification of Parasaccharibacter Apium, Saccharibacter sp. and Bombella sp. Strains
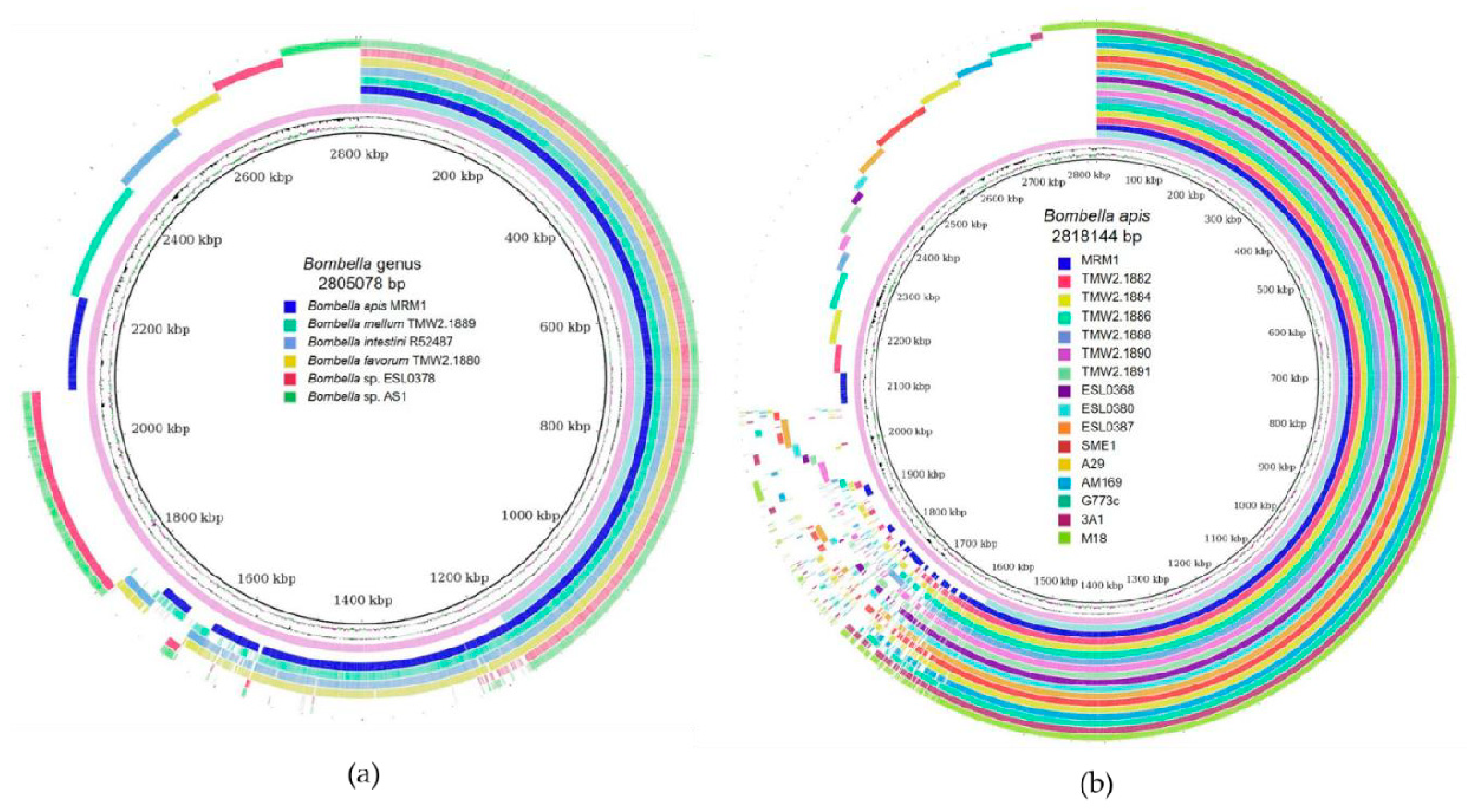
3.2. Origin and General Genomic Features of Bombella sp. Isolates
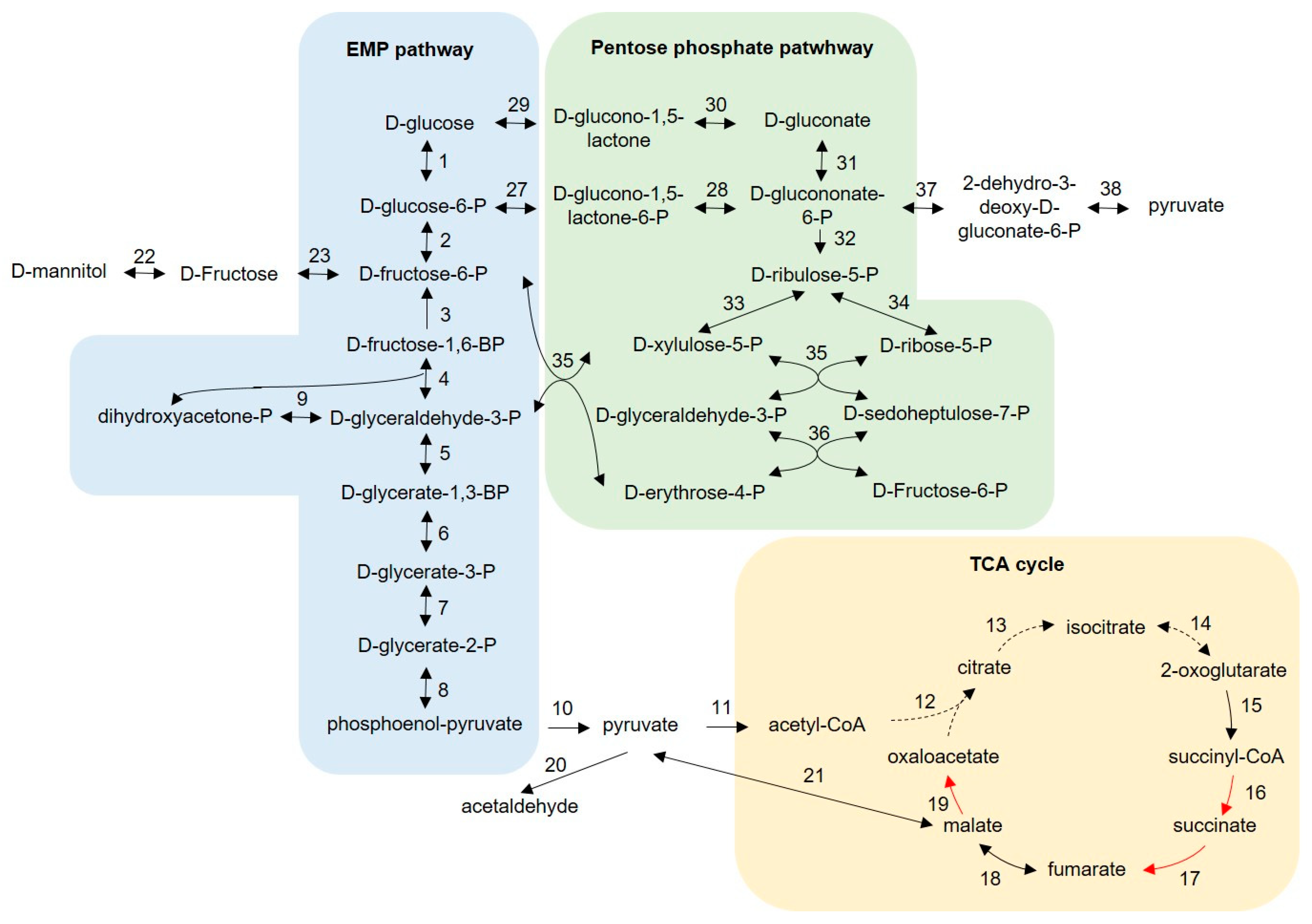
3.3. Main Cytoplasmic Carbohydrate Metabolism
3.4. Membrane-Bound Dehydrogenases and Respiratory Enzymes
3.5. Presence of Prophages and Bacteriocins
3.6. Polyketide Synthase Gene Cluster
3.7. Tetracycline Resistance
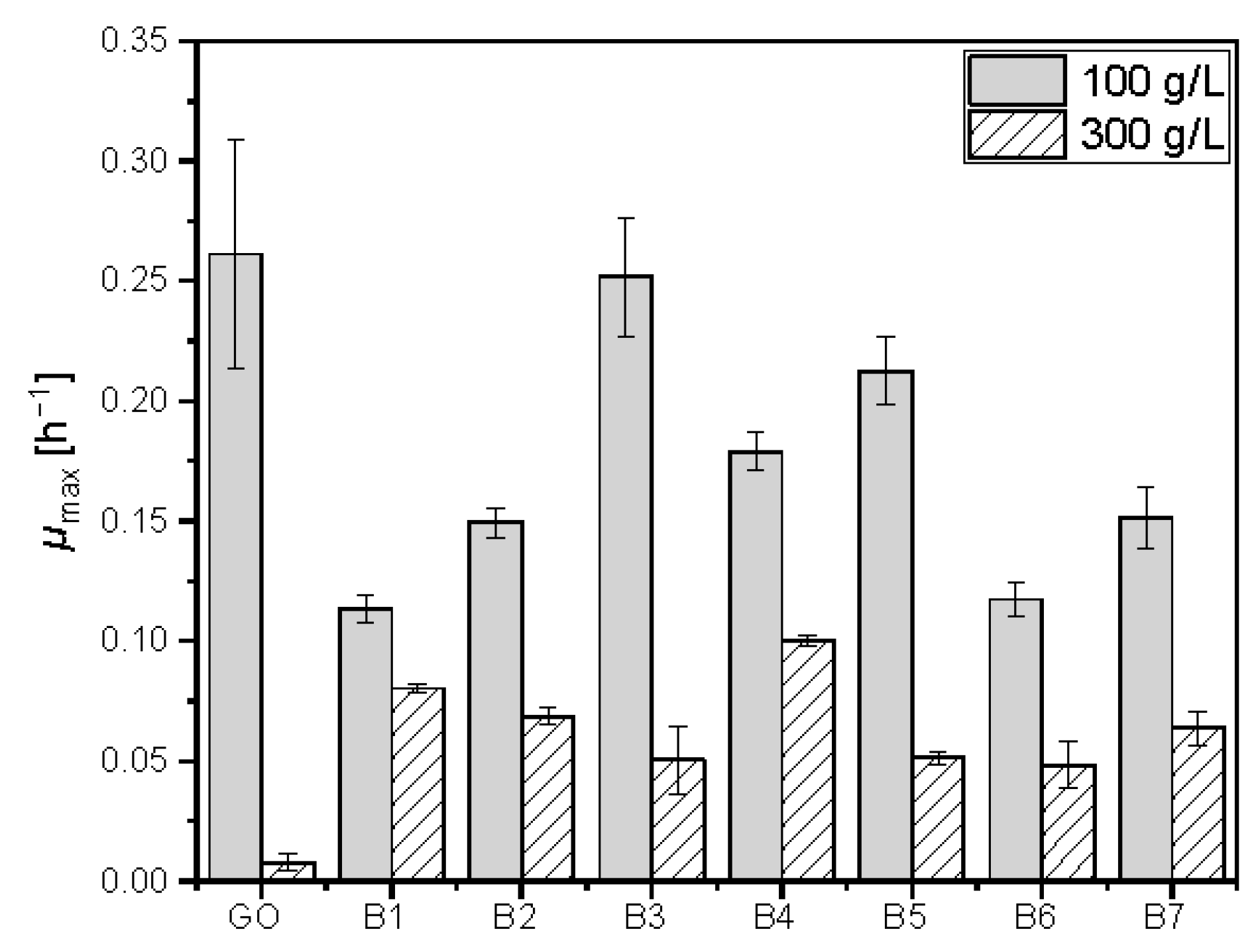
3.8. Adaptation to High Glucose Concentration
4. Discussion
4.1. Phylogeny, Current Taxonomy and Reclassification of Parasaccharibacter apium, Saccharibacter sp. and Bombella sp. Strains
4.2. Predictive Carbohydrate Metabolism of Bombella spp.
4.3. Potential Role of Extracellular Invertase in Melezitose Degradation
4.4. Potential Antifungal Properties
4.5. Tetracycline Resistance
4.6. Sugar Tolerance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breeze, T.D.; Bailey, A.P.; Balcombe, K.G.; Potts, S.G. Pollination services in the UK: How important are honeybees? Agric. Ecosyst. Environ. 2011, 142, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Free, J.B. Insect Pollination of Crops, 2nd ed.; Academic Press: London, UK, 1993; ISBN 978-01-2266-651-3. [Google Scholar]
- Brodschneider, R.; Gray, A.; Adjlane, N.; Ballis, A.; Brusbardis, V.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Dahle, B.; de Graaf, D.C.; et al. Multi-country loss rates of honey bee colonies during winter 2016/2017 from the COLOSS survey. J. Apic. Res. 2018, 57, 452–457. [Google Scholar] [CrossRef] [Green Version]
- van Engelsdorp, D.; Hayes, J.; Underwood, R.M.; Pettis, J. A survey of honey bee colony losses in the U.S., fall 2007 to spring 2008. PLoS ONE 2008, 3, e4071. [Google Scholar]
- Paxton, R.J. Does infection by Nosema ceranae cause “Colony Collapse Disorder” in honey bees (Apis mellifera)? J. Apic. Res. 2010, 49, 80–84. [Google Scholar] [CrossRef]
- Johnson, R.M.; Ellis, M.D.; Mullin, C.A.; Frazier, M. Pesticides and honey bee toxicity—USA. Apidologie 2010, 41, 312–331. [Google Scholar] [CrossRef] [Green Version]
- Kaltenpoth, M.; Göttler, W.; Herzner, G.; Strohm, E. Symbiotic bacteria protect wasp larvae from fungal infestation. Curr. Biol. 2005, 15, 475–479. [Google Scholar] [CrossRef] [Green Version]
- van Arnam, E.B.; Currie, C.R.; Clardy, J. Defense contracts: Molecular protection in insect-microbe symbioses. Chem. Soc. Rev. 2018, 47, 1638–1651. [Google Scholar] [CrossRef]
- Moran, N.A. Genomics of the honey bee microbiome. Curr. Opin. Insect Sci. 2015, 10, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.-H.; Lee, J.-Y.; Hyun, D.-W.; Jung, M.-J.; Bae, J.-W. Bombella apis sp. nov., an acetic acid bacterium isolated from the midgut of a honey bee. Int. J. Syst. Evol. Microbiol. 2017, 67, 2184–2188. [Google Scholar] [CrossRef]
- Li, L.; Praet, J.; Borremans, W.; Nunes, O.C.; Manaia, C.M.; Cleenwerck, I.; Meeus, I.; Smagghe, G.; de Vuyst, L.; Vandamme, P. Bombella intestini gen. nov., sp. nov., an acetic acid bacterium isolated from bumble bee crop. Int. J. Syst. Evol. Microbiol. 2015, 65, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Hilgarth, M.; Redwitz, J.; Ehrmann, M.A.; Vogel, R.F.; Jakob, F. Bombella favorum sp. nov. and Bombella mellum sp. nov., two novel species isolated from the honeycombs of Apis mellifera. Int. J. Syst. Evol. Microbiol. 2021, 71, 004633. [Google Scholar] [CrossRef] [PubMed]
- Corby-Harris, V.; Snyder, L.A.; Schwan, M.R.; Maes, P.; McFrederick, Q.S.; Anderson, K.E. Origin and effect of Alpha 2.2 Acetobacteraceae in honey bee larvae and description of Parasaccharibacter apium gen. nov., sp. nov. Appl. Environ. Microbiol. 2014, 80, 7460–7472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corby-Harris, V.; Snyder, L.; Meador, C.A.D.; Naldo, R.; Mott, B.; Anderson, K.E. Parasaccharibacter apium, gen. nov., sp. nov., Improves Honey Bee (Hymenoptera: Apidae) Resistance to Nosema. J. Econ. Entomol. 2016, 109, 537–543. [Google Scholar] [PubMed] [Green Version]
- Miller, D.L.; Smith, E.A.; Newton, I.L.G. A Bacterial Symbiont Protects Honey Bees from Fungal Disease. mBio 2021, 12, e0050321. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Sheehan, T.H.; Mott, B.M.; Maes, P.; Snyder, L.; Schwan, M.R.; Walton, A.; Jones, B.M.; Corby-Harris, V. Microbial ecology of the hive and pollination landscape: Bacterial associates from floral nectar, the alimentary tract and stored food of honey bees (Apis mellifera). PLoS ONE 2013, 8, e83125. [Google Scholar] [CrossRef] [Green Version]
- Tarpy, D.R.; Mattila, H.R.; Newton, I.L.G. Development of the honey bee gut microbiome throughout the queen-rearing process. Appl. Environ. Microbiol. 2015, 81, 3182–3191. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Illeghems, K.; van Kerrebroeck, S.; Borremans, W.; Cleenwerck, I.; Smagghe, G.; de Vuyst, L.; Vandamme, P. Whole-Genome Sequence Analysis of Bombella intestini LMG 28161T, a Novel Acetic Acid Bacterium Isolated from the Crop of a Red-Tailed Bumble Bee, Bombus lapidarius. PLoS ONE 2016, 11, e0165611. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.A.; Newton, I.L.G. Genomic Signatures of Honey Bee Association in an Acetic Acid Symbiont. Genome Biol. Evol. 2020, 12, 1882–1894. [Google Scholar] [CrossRef]
- Huptas, C.; Scherer, S.; Wenning, M. Optimized Illumina PCR-free library preparation for bacterial whole genome sequencing and analysis of factors influencing de novo assembly. BMC Res. Notes 2016, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.; Chalita, M.; Ha, S.-M.; Na, S.-I.; Yoon, S.-H.; Chun, J. ContEst16S: An algorithm that identifies contaminated prokaryotic genomes using 16S RNA gene sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2053–2057. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinf. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Werum, V.; Ehrmann, M.; Vogel, R.; Hilgarth, M. Comparative genome analysis, predicted lifestyle and antimicrobial strategies of Lactococcus carnosus and Lactococcus paracarnosus isolated from meat. Microbiol. Res. 2022, 258, 126982. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Moore, W.E.C.; Stackebrandt, E.; Kandler, O.; Colwell, R.R.; Krichevsky, M.I.; Truper, H.G.; Murray, R.G.E.; Wayne, L.G.; Grimont, P.A.D.; Brenner, D.J.; et al. Report of the Ad Hoc Committee on Reconciliation of Approaches to Bacterial Systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Behr, J.; Geissler, A.J.; Schmid, J.; Zehe, A.; Vogel, R.F. The Identification of Novel Diagnostic Marker Genes for the Detection of Beer Spoiling Pediococcus damnosus Strains Using the BlAst Diagnostic Gene findEr. PLoS ONE 2016, 11, e0152747. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Kahm, M.; Hasenbrink, G.; Lichtenberg-Fraté, H.; Ludwig, J.; Kschischo, M. Grofit: Fitting Biological Growth Curves. Nat. Proc. 2010, 33, 1–21. [Google Scholar] [CrossRef]
- Bonilla-Rosso, G.; Paredes Juan, C.; Das, S.; Ellegaard, K.M.; Emery, O.; Garcia-Garcera, M.; Glover, N.; Hadadi, N.; van der Meer, J.R.; Tagini, F.; et al. Acetobacteraceae in the honey bee gut comprise two distant clades with diverging metabolism and ecological niches. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Chouaia, B.; Gaiarsa, S.; Crotti, E.; Comandatore, F.; Degli Esposti, M.; Ricci, I.; Alma, A.; Favia, G.; Bandi, C.; Daffonchio, D. Acetic acid bacteria genomes reveal functional traits for adaptation to life in insect guts. Genome Biol. Evol. 2014, 6, 912–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corby-Harris, V.; Anderson, K.E. Draft Genome Sequences of Four Parasaccharibacter apium Strains Isolated from Honey Bees. Genome Announc. 2018, 6, e00165-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veress, A.; Wilk, T.; Kiss, J.; Olasz, F.; Papp, P.P. Draft Genome Sequences of Saccharibacter sp. Strains 3.A.1 and M18 Isolated from Honey and a Honey Bee (Apis mellifera) Stomach. Genome Announc. 2017, 5, e00744-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prust, C.; Hoffmeister, M.; Liesegang, H.; Wiezer, A.; Fricke, W.F.; Ehrenreich, A.; Gottschalk, G.; Deppenmeier, U. Complete genome sequence of the acetic acid bacterium Gluconobacter oxydans. Nat. Biotechnol. 2005, 23, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Anderson, K.E.; Corby-Harris, V.; McFrederick, Q.S.; Parish, A.J.; Rice, D.W.; Newton, I.L.G. Reclassification of seven honey bee symbiont strains as Bombella apis. Int. J. Syst. Evol. Microbiol. 2021, 71, 4950. [Google Scholar] [CrossRef]
- Azuma, Y.; Hosoyama, A.; Matsutani, M.; Furuya, N.; Horikawa, H.; Harada, T.; Hirakawa, H.; Kuhara, S.; Matsushita, K.; Fujita, N.; et al. Whole-genome analyses reveal genetic instability of Acetobacter pasteurianus. Nucleic Acids Res. 2009, 37, 5768–5783. [Google Scholar] [CrossRef] [Green Version]
- Illeghems, K.; de Vuyst, L.; Weckx, S. Complete genome sequence and comparative analysis of Acetobacter pasteurianus 386B, a strain well-adapted to the cocoa bean fermentation ecosystem. BMC Genom. 2013, 14, 526. [Google Scholar] [CrossRef] [Green Version]
- Hanke, T.; Nöh, K.; Noack, S.; Polen, T.; Bringer, S.; Sahm, H.; Wiechert, W.; Bott, M. Combined fluxomics and transcriptomics analysis of glucose catabolism via a partially cyclic pentose phosphate pathway in Gluconobacter oxydans 621H. Appl. Environ. Microbiol. 2013, 79, 2336–2348. [Google Scholar] [CrossRef] [Green Version]
- Mullins, E.A.; Francois, J.A.; Kappock, T.J. A specialized citric acid cycle requiring succinyl-coenzyme A (CoA): Acetate CoA-transferase (AarC) confers acetic acid resistance on the acidophile Acetobacter aceti. J. Bacteriol. 2008, 190, 4933–4940. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.P.; Wernegreen, J.J. Genomic erosion and extensive horizontal gene transfer in gut-associated Acetobacteraceae. BMC Genom. 2019, 20, 472. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, K.; Toyama, H.; Adachi, O. Respiratory Chains and Bioenergetics of Acetic Acid Bacteria. Adv. Microb. Physiol. 1994, 36, 247–301. [Google Scholar] [PubMed]
- Adachi, O.; Moonmangmee, D.; Toyama, H.; Yamada, M.; Shinagawa, E.; Matsushita, K. New developments in oxidative fermentation. Appl. Microbiol. Biotechnol. 2003, 60, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; Fujii, Y.; Ano, Y.; Toyama, H.; Shinjoh, M.; Tomiyama, N.; Miyazaki, T.; Sugisawa, T.; Hoshino, T.; Adachi, O. 5-keto-D-gluconate production is catalyzed by a quinoprotein glycerol dehydrogenase, major polyol dehydrogenase, in Gluconobacter species. Appl. Environ. Microbiol. 2003, 69, 1959–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richhardt, J.; Luchterhand, B.; Bringer, S.; Büchs, J.; Bott, M. Evidence for a key role of cytochrome bo3 oxidase in respiratory energy metabolism of Gluconobacter oxydans. J. Bacteriol. 2013, 195, 4210–4220. [Google Scholar] [CrossRef] [Green Version]
- Simo, K.; Christensen, G.M. Quantitative analysis of sugars in royal jelly. Nature 1962, 196, 1208–1209. [Google Scholar] [CrossRef]
- Siddiqui, I.R. The Sugars of Honey. Adv. Carbohydr. Chem. Biochem. 1970, 25, 285–309. [Google Scholar]
- Seeburger, V.C.; D’Alvise, P.; Shaaban, B.; Schweikert, K.; Lohaus, G.; Schroeder, A.; Hasselmann, M. The trisaccharide melezitose impacts honey bees and their intestinal microbiota. PLoS ONE 2020, 15, e0230871. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D. Diverse origins of tetracycline resistance in the honey bee bacterial pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2003, 83, 46–50. [Google Scholar] [CrossRef]
- Raymann, K.; Shaffer, Z.; Moran, N.A. Antibiotic exposure perturbs the gut microbiota and elevates mortality in honeybees. PLoS Biol. 2017, 15, e2001861. [Google Scholar] [CrossRef]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [Green Version]
- Geng, S.; Pan, X.-C.; Mei, R.; Wang, Y.-N.; Sun, J.-Q.; Liu, X.-Y.; Tang, Y.-Q.; Wu, X.-L. Paradevosia shaoguanensis gen. nov., sp. nov., isolated from a coking wastewater. Curr. Microbiol. 2015, 70, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Empadinhas, N.; da Costa, M.S. Osmoadaptation mechanisms in prokaryotes: Distribution of compatible solutes. Int. Microbiol. 2008, 11, 151–161. [Google Scholar] [PubMed]
- Zahid, N.; Deppenmeier, U. Role of mannitol dehydrogenases in osmoprotection of Gluconobacter oxydans. Appl. Microbiol. Biotechnol. 2016, 100, 9967–9978. [Google Scholar] [CrossRef] [PubMed]
- Leandro, M.; Andrade, L.; Vespoli, L.; Moreira, J.; Pimentel, V.; Soares, F.; Passamani, L.; Silveira, V.; de Souza Filho, G. Comparative proteomics reveals essential mechanisms for osmotolerance in Gluconacetobacter diazotrophicus. Res. Microbiol. 2021, 172, 103785. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Contigs | Size (mbp) | GC (%) | Density (%) | Source | Symbiotic Host | Country | Reference | BioSample |
|---|---|---|---|---|---|---|---|---|---|
| Bombella apis MRM1T | 7 | 2.03 | 59.59 | 90.55 | midgut | Apis mellifera | KOR | [11] | SAMN16262074 |
| Bombella apis ESL0368 | 1 | 1.99 | 59.6 | 89.85 | gut of adult queen | Apis mellifera | CHE | [41] | SAMN13280441 |
| Bombella apis ESL0380 | 10 | 1.98 | 59.57 | 90.43 | gut of adult queen | Apis mellifera | CHE | [41] | SAMN13280444 |
| Bombella apis ESL0387 | 10 | 2.04 | 59.47 | 90.3 | gut of adult queen | Apis mellifera | CHE | [41] | SAMN13280447 |
| Bombella apis SME1 | 11 | 2.09 | 59.56 | 90.09 | beehive | Apis mellifera | USA | - | SAMN13042715 |
| Bombella apis TMW 2.1882 | 5 | 2.02 | 59.41 | 90.33 | honey | Apis mellifera | DEU | this study | SAMN07674798 |
| Bombella apis TMW 2.1884 | 19 | 2.05 | 59.37 | 90.33 | honey | Apis mellifera | DEU | this study | SAMN09635582 |
| Bombella apis TMW 2.1886 | 39 | 2.05 | 58.86 | 89.57 | honey | Apis mellifera mellifera | AUT | this study | SAMN09635591 |
| Bombella apis TMW 2.1888 | 1 | 2.01 | 59.48 | 89.72 | royal jelly | Apis mellifera mellifera | AUT | this study | SAMN09641705 |
| Bombella apis TMW 2.1890 | 7 | 2.02 | 59.49 | 90.29 | honey | Apis mellifera | DEU | this study | SAMN07675059 |
| Bombella apis TMW 2.1891 | 7 | 2.01 | 59.44 | 90.46 | honey | Apis mellifera | DEU | this study | SAMN09635595 |
| Bombella apis A29 | 27 | 2.01 | 59.39 | 90.15 | larva | Apis mellifera | USA | [14] | SAMN04240487 |
| Bombella apis AM169 | 9 | 1.98 | 59.32 | 90 | gut adult | Apis mellifera | ITA | [42] | SAMEA3139036 |
| Bombella apis G773c | 1 | 2.01 | 59.42 | 90.02 | hindgut adult | Apis mellifera | USA | [43] | SAMN06649799 |
| Bombella apis 3A1 | 24 | 2.01 | 59.41 | 90.01 | honey | Apis mellifera | HUN | [44] | SAMN05935507 |
| Bombella apis M18 | 11 | 2.08 | 59.35 | 90.22 | stomach adult | Apis mellifera | HUN | [44] | SAMN05935506 |
| Bombella intestini R52487T | 12 | 2.02 | 54.94 | 90.16 | crop | Bombus lapidarius | BEL | [19] | SAMN02598725 |
| Bombella favorum TMW 2.1880T | 7 | 1.98 | 55.33 | 90.29 | honey | Apis mellifera | DEU | [13] | SAMN07674723 |
| Bombella mellum TMW 2.1889T | 11 | 2.07 | 60.43 | 89.93 | honey | Apis mellifera | DEU | [13] | SAMN07674951 |
| Bombella sp. ESL0378 | 15 | 1.85 | 52.88 | 91.34 | gut of adult worker | Apis mellifera | CHE | [41] | SAMN13280442 |
| Bombella sp. ESL0385 | 5 | 1.9 | 52.91 | 91.51 | gut of adult worker | Apis mellifera | CHE | [41] | SAMN13280446 |
| Bombella sp. AS1 | 13 | 1.85 | 52.64 | 91.55 | larva | Apis mellifera | USA | - | SAMN05720096 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Härer, L.; Hilgarth, M.; Ehrmann, M.A. Comparative Genomics of Acetic Acid Bacteria within the Genus Bombella in Light of Beehive Habitat Adaptation. Microorganisms 2022, 10, 1058. https://doi.org/10.3390/microorganisms10051058
Härer L, Hilgarth M, Ehrmann MA. Comparative Genomics of Acetic Acid Bacteria within the Genus Bombella in Light of Beehive Habitat Adaptation. Microorganisms. 2022; 10(5):1058. https://doi.org/10.3390/microorganisms10051058
Chicago/Turabian StyleHärer, Luca, Maik Hilgarth, and Matthias A. Ehrmann. 2022. "Comparative Genomics of Acetic Acid Bacteria within the Genus Bombella in Light of Beehive Habitat Adaptation" Microorganisms 10, no. 5: 1058. https://doi.org/10.3390/microorganisms10051058
APA StyleHärer, L., Hilgarth, M., & Ehrmann, M. A. (2022). Comparative Genomics of Acetic Acid Bacteria within the Genus Bombella in Light of Beehive Habitat Adaptation. Microorganisms, 10(5), 1058. https://doi.org/10.3390/microorganisms10051058