



Biodegradation of Oil by a Newly Isolated Strain Acinetobacter junii WCO-9 and Its Comparative Pan-Genome Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Screening of Strains
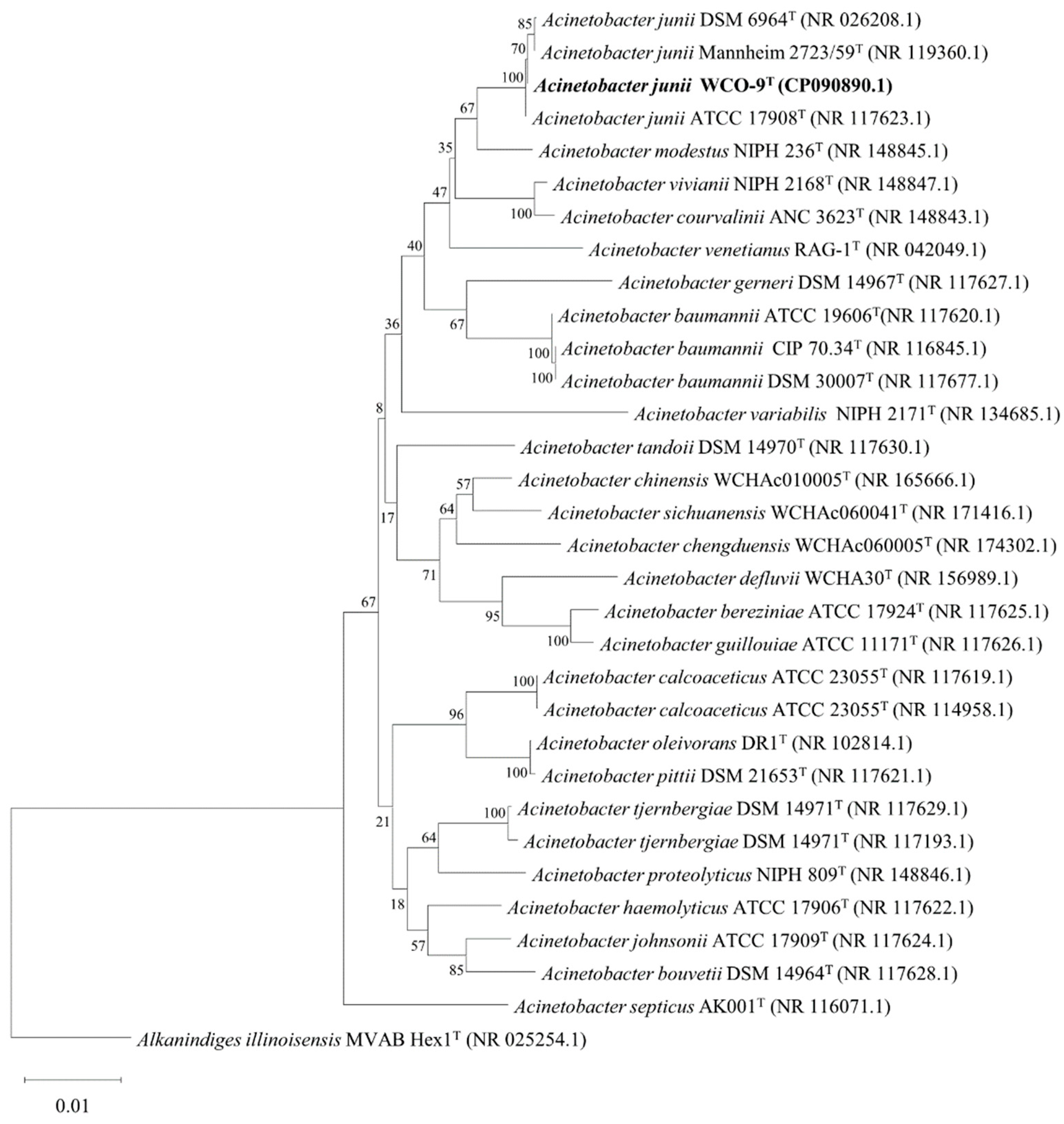
2.2. Phylogenetic Analysis of 16S rDNA and Housekeeping Genes
2.3. Comparison of the Growth and Oil Degradation Capabilities of Strain WCO-9
2.4. Whole Genome Sequencing and Pan-Genome Analysis
2.5. Analysis of Lipase Genes and Triacylglycerol Degradation Pathway
2.6. Statistical Analysis
3. Results
3.1. Screening and Identification of Strain WCO-9
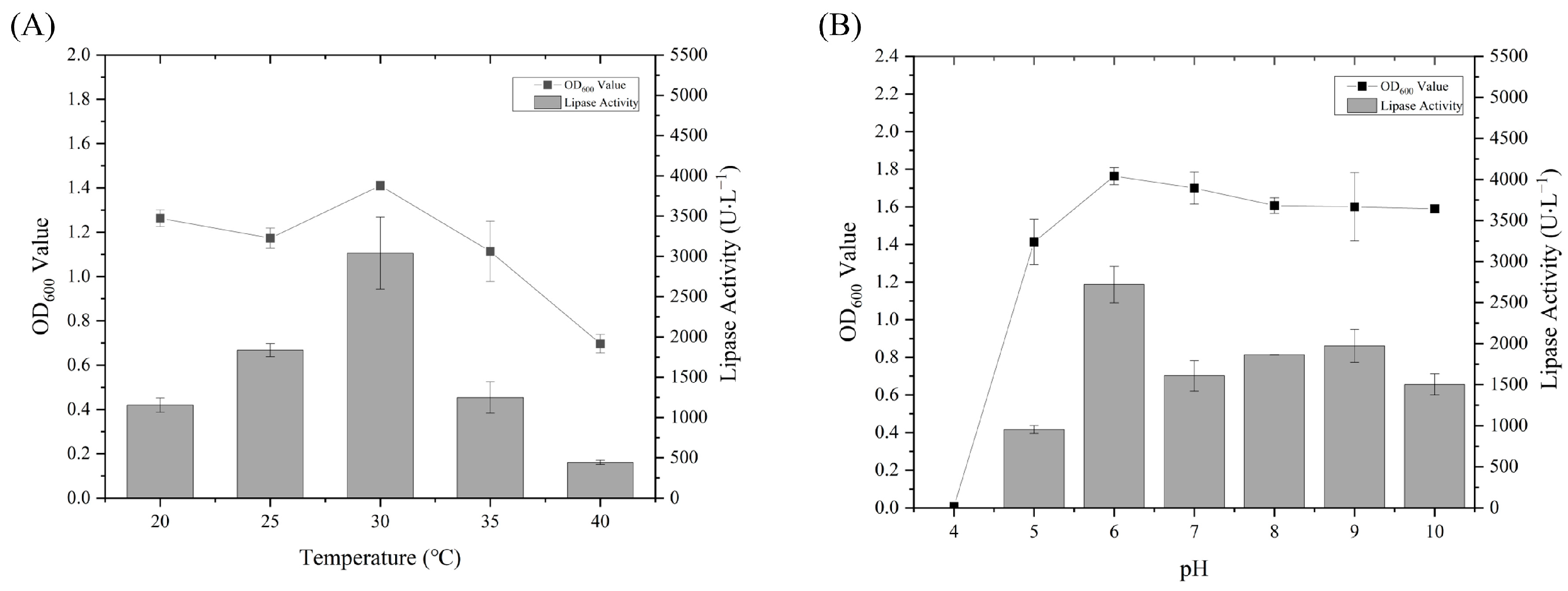
3.2. Growth Characteristics and Oil Degradation Capabilities of Strain WCO-9
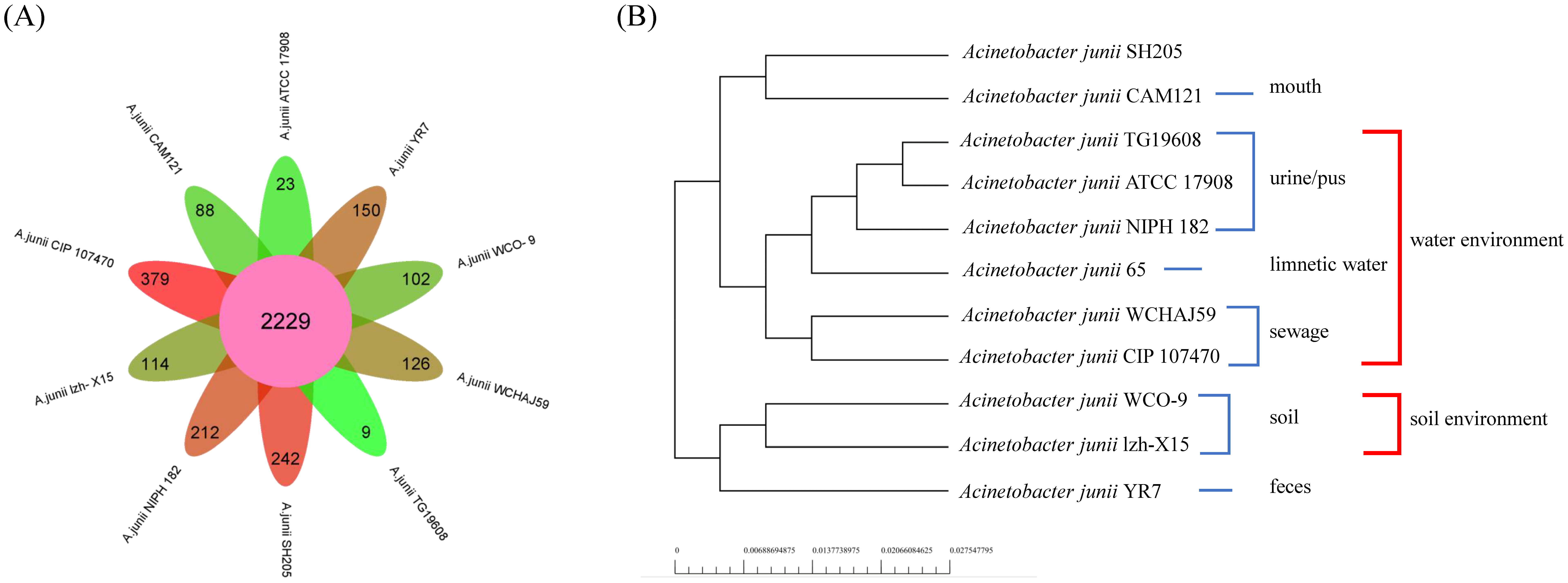
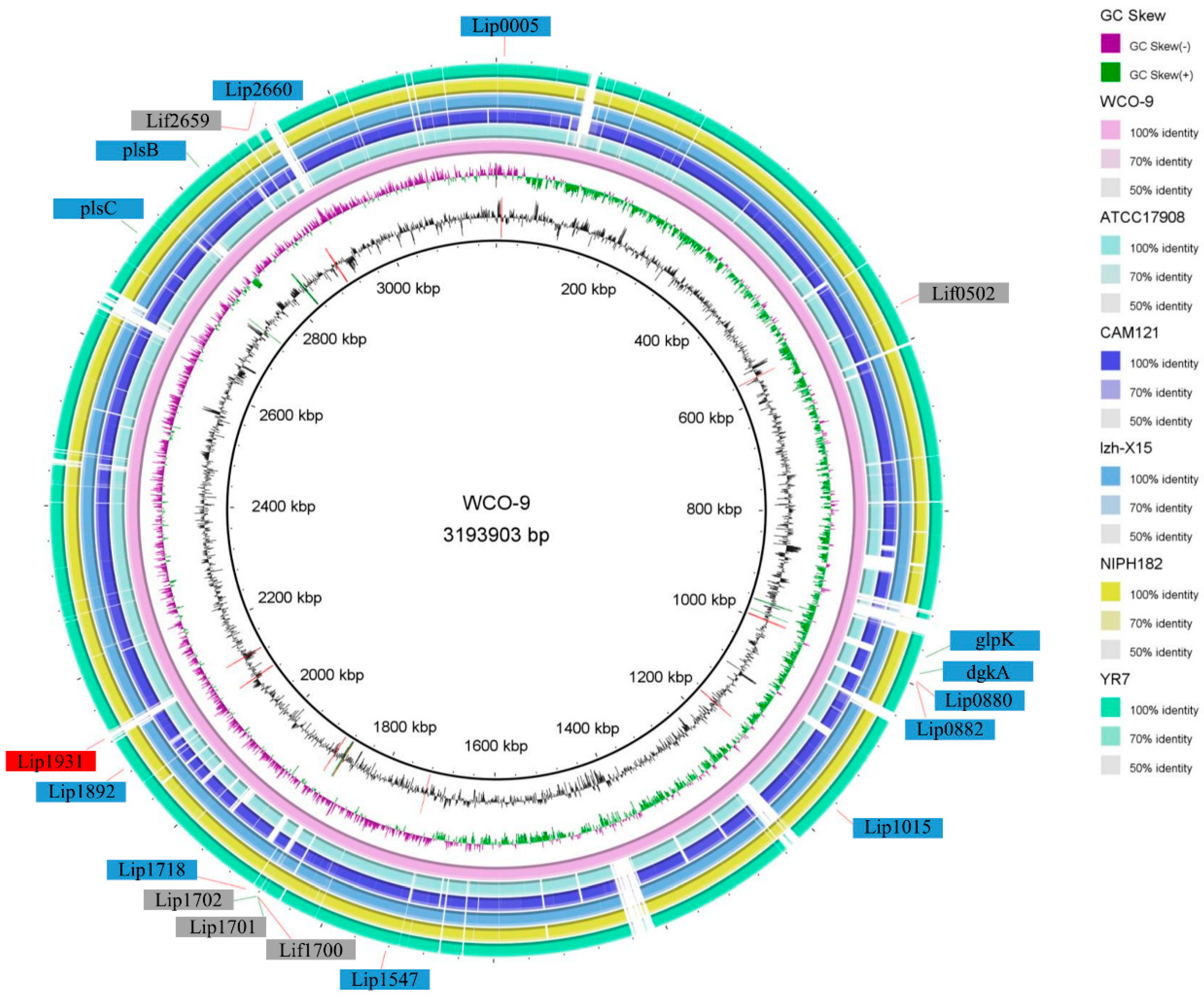
3.3. Genome Comparison and Pan-Genome Analysis of WCO-9
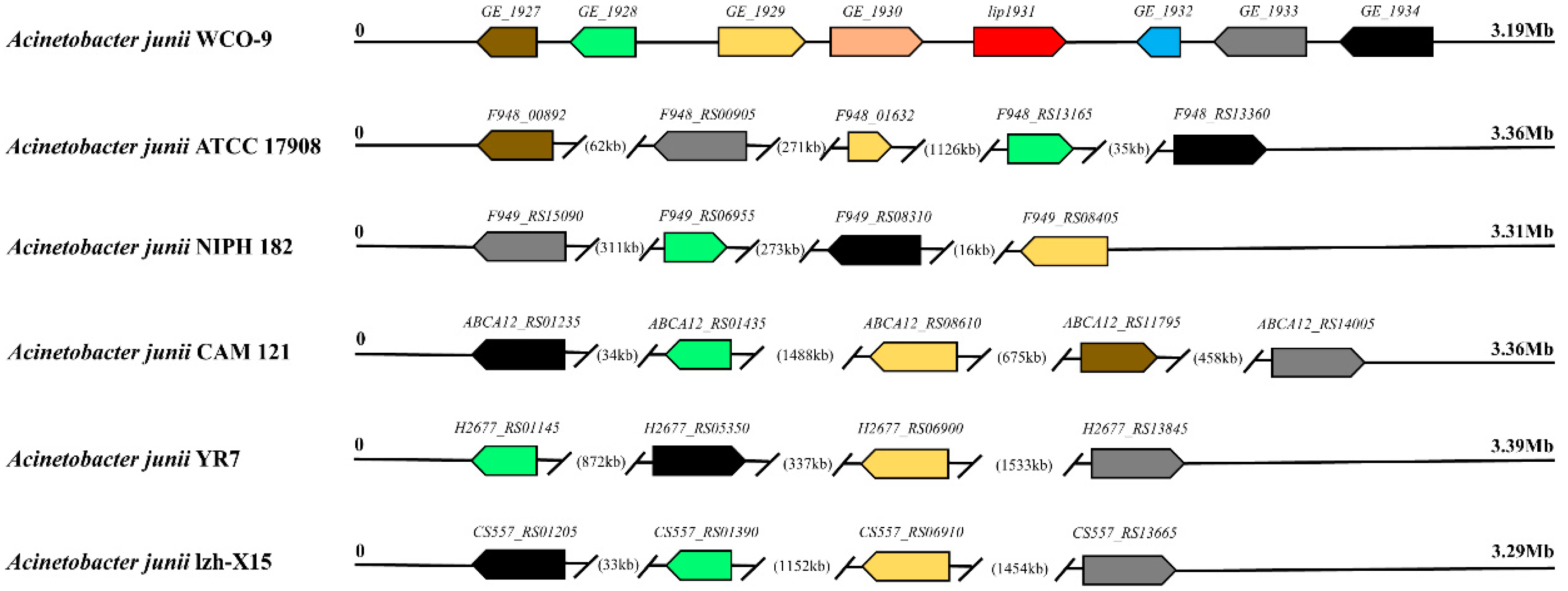
3.4. Comparison of Lipase Genes and Triacylglycerol Metabolic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giwa, A.S.; Ali, N.; Vakili, M.; Guo, X.; Liu, D.; Wang, K. Opportunities for holistic waste stream valorization from food waste treatment facilities: A review. Rev. Chem. Eng. 2022, 38, 35–53. [Google Scholar] [CrossRef]
- Zeng, J.; Zeng, H.; Wang, Z. Review on technology of making biofuel from food waste. Int. J. Energ. Res. 2022, 46, 10301–10319. [Google Scholar] [CrossRef]
- Kim, K.; Jung, J.Y.; Kwon, J.H. Potential of Bacterial Strains Isolated from Coastal Water for Wastewater Treatment and as Aqua-Feed Additives. Microorganisms 2021, 9, 2441. [Google Scholar] [CrossRef] [PubMed]
- Wahi, R.; Chuah, L.A.; Choong, T.S.Y.; Ngaini, Z.; Nourouzi, M.M. Oil removal from aqueous state by natural fibrous sorbent: An overview. Sep. Purif. Technol. 2013, 113, 51–63. [Google Scholar] [CrossRef]
- Zhou, S.P.; Ke, X.; Zhou, H.Y.; Zou, S.P.; Xue, Y.P.; Zheng, Y.G. Community scale in-situ rapid biological reduction and resource recovery of food waste. Bioresour. Technol. 2022, 346, 126603. [Google Scholar] [CrossRef]
- Liu, P.; Ji, J.; Wu, Q.; Ren, J.; Wu, G.; Yu, Z.; Xiong, J.; Tian, F.; Zafar, Y.; Li, X. Klebsiella pneumoniae sp. LZU10 degrades oil in food waste and enhances methane production from co-digestion of food waste and straw. Int. Biodeter. Biodegr. 2018, 126, 28–36. [Google Scholar] [CrossRef]
- Ke, X.; Hua, X.; Sun, J.C.; Zheng, R.C.; Zheng, Y.G. Synergetic degradation of waste oil by constructed bacterial consortium for rapid in-situ reduction of kitchen waste. J. Biosci. Bioeng. 2021, 131, 412–419. [Google Scholar] [CrossRef]
- Liu, D.; Ma, X.; Huang, J.; Shu, Z.; Chu, X.; Li, Y.; Jin, Y. Study on personalized microbial formulation during high-temperature aerobic fermentation of different types of food wastes. Sci. Total Environ. 2022, 814, 152561. [Google Scholar] [CrossRef]
- Tsegaye, B.; Jaiswal, S.; Jaiswal, A.K. Food Waste Biorefinery: Pathway towards Circular Bioeconomy. Foods 2021, 10, 1174. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Wan, Y.; Zhang, S.; Luo, J.; Li, Y.Y.; Liu, J. Valorization of food waste to multiple bio-energies based on enzymatic pretreatment: A critical review and blueprint for the future. J. Clean. Prod. 2020, 277, 124091. [Google Scholar] [CrossRef]
- Strazzera, G.; Battista, F.; Garcia, N.H.; Frison, N.; Bolzonella, D. Volatile fatty acids production from food wastes for biorefinery platforms: A review. J. Environ. Manag. 2018, 226, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Hafid, H.S.; Rahman, N.A.A.; Shah, U.K.M.; Baharuddin, A.S.; Ariff, A.B. Feasibility of using kitchen waste as future substrate for bioethanol production: A review. Renew. Sust. Energ. Rev. 2017, 74, 671–686. [Google Scholar] [CrossRef]
- Kumari, A.; Ahmad, R.; Negi, S.; Khare, S.K. Biodegradation of waste grease by Penicillium chrysogenum for production of fatty acid. Bioresour. Technol. 2017, 226, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Yang, L.M.; Lv, P.M.; Wang, Z.Y.; Fu, J.Y.; Miao, C.L.; Li, Z.B.; Li, L.H.; Liu, T.; Du, W.Y.; et al. Improvement of methanol tolerance and catalytic activity of Rhizomucor miehei lipase for one-step synthesis of biodiesel by semi-rational design. Bioresour. Technol. 2022, 348, 126769. [Google Scholar] [CrossRef]
- Adewoyin, M.A.; Okoh, A.I. The natural environment as a reservoir of pathogenic and non-pathogenic Acinetobacter species. Rev. Environ. Health 2018, 33, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Haseley, S.R.; Pantophlet, R.; Brade, L.; Holst, O.; Brade, H. Structural and serological characterisation of the O-antigenic polysaccharide of the lipopolysaccharide from Acinetobacter junii strain 65. Eur. J. Biochem. 1997, 245, 477–481. [Google Scholar] [CrossRef]
- Maslunka, C.; Guertler, V.; Seviour, R.J. The impact of horizontal gene transfer on targeting the internal transcribed spacer region (ITS) to identify Acinetobacter junii strains. J. Appl. Microbiol. 2015, 118, 1435–1443. [Google Scholar] [CrossRef]
- Linde, H.J.; Hahn, J.; Holler, E.; Reischl, U.; Lehn, N. Septicemia due to Acinetobacter junii. J. Clin. Microbiol. 2002, 40, 2696–2697. [Google Scholar] [CrossRef]
- Hansen, W.; Schoutens, E.; Yourassowsky, E. Taxonomy of Acinetobacter: The usefulness of beta-D-xyloside xylohydrolase for strain differentiation. J. Clin. Pathol. 1977, 30, 838–841. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.K.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Tabaraki, R.; Ahmady-Asbchin, S.; Abdi, O. Biosorption of Zn (II) from aqueous solutions by Acinetobacter sp. isolated from petroleum spilled soil. J. Environ. Chem. Eng. 2013, 1, 604–608. [Google Scholar] [CrossRef]
- Zheng, J.; Feng, J.Q.; Zhou, L.; Mbadinga, S.M.; Gu, J.-D.; Mu, B.Z. Characterization of bacterial composition and diversity in a long-term petroleum contaminated soil and isolation of high-efficiency alkane-degrading strains using an improved medium. World J. Microb. Biot. 2018, 34, 34. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Wang, R.; Rao, P.; Wu, B.; Yan, L.; Hu, L.; Park, S.; Ryu, M.; Zhou, X. Bioremediation of Petroleum Hydrocarbons Using Acinetobacter sp. SCYY-5 Isolated from Contaminated Oil Sludge: Strategy and Effectiveness Study. Int. J. Environ. Res. Public Health 2021, 18, 819. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.M.; Perez, F.; Pan, Q.; Jiang, Y.; Evans, S.R.; Bonomo, R.A. Acinetobacter Infections. Clin. Infect. Dis. 2020, 71, 1357–1358. [Google Scholar] [CrossRef] [PubMed]
- Weber, B.S.; Kinsella, R.L.; Harding, C.M.; Feldman, M.F. The Secrets of Acinetobacter Secretion. Trends Microbiol. 2017, 25, 533–546. [Google Scholar] [CrossRef]
- Gao, L.L.; Lu, Y.C.; Zhang, J.L.; Li, J.; Zhang, J.D. Biotreatment of restaurant wastewater with an oily high concentration by newly isolated bacteria from oily sludge. World J. Microb. Biot. 2019, 35, 179. [Google Scholar] [CrossRef]
- Ke, X.; Sun, J.C.; Liu, C.; Ying, J.M.; Zou, S.P.; Xue, Y.P.; Zheng, Y.G. Fed-in-situ biological reduction treatment of food waste via high-temperature-resistant oil degrading microbial consortium. Bioresour. Technol. 2021, 340, 125635. [Google Scholar] [CrossRef]
- Cai, L.; Guo, H.T.; Zheng, G.D.; Wang, X.Y.; Wang, K. Metagenomic analysis reveals the microbial degradation mechanism during kitchen waste biodrying. Chemosphere 2022, 307, 135862. [Google Scholar] [CrossRef]
- Adrio, J.L.; Demain, A.L. Recombinant organisms for production of industrial products. Bioeng. Bugs 2010, 1, 116–131. [Google Scholar] [CrossRef]
- Melani, N.B.; Tambourgi, E.B.; Silveira, E. Lipases: From Production to Applications. Sep. Purif. Rev. 2020, 49, 143–158. [Google Scholar] [CrossRef]
- Marchut-Mikolajczyk, O.; Drozdzynski, P.; Struszczyk-Swita, K. Biodegradation of slop oil by endophytic Bacillus cereus EN18 coupled with lipase from Rhizomucor miehei (Palatase®). Chemosphere 2020, 250, 126203. [Google Scholar] [CrossRef]
- Cavalcante, F.T.T.; Neto, F.S.; Falca, I.R.D.; Souza, J.E.D.; de Moura Junior, L.S.; Sousa, P.D.; Rocha, T.G.; de Sousa, I.G.; Gomes, P.H.D.; de Souza, M.C.M.; et al. Opportunities for improving biodiesel production via lipase catalysis. Fuel 2021, 288, 119577. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J.H.; Kim, D.S.; Yoo, H.Y.; Park, C.; Kim, S.W. Biodiesel production by lipases co-immobilized on the functionalized activated carbon. Bioresour. Technol. Rep. 2019, 7, 100248. [Google Scholar] [CrossRef]
- Zuo, K.; Zhang, L.; Yao, H.; Wang, J. Isolation and functional expression of a novel lipase gene isolated directly from oil-contaminated soil. Acta Biochim. Pol. 2010, 57, 305–311. [Google Scholar] [CrossRef]
- Tan, Z.; Chen, G.; Zhao, Y.; Shi, H.; Wang, S.; Bilal, M.; Li, X. Digging and identification of novel microorganisms from the soil environments with high methanol-tolerant lipase production for biodiesel preparation. Environ. Res. 2022, 212, 113570. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, W.; Shah, S.B.; Zanaroli, G.; Xu, P.; Tang, H. Phenol biodegradation by Acinetobacter radioresistens APH1 and its application in soil bioremediation. Appl. Microbiol. Biot. 2020, 104, 427–437. [Google Scholar] [CrossRef]
- Zhou, H.; Huang, X.; Liang, Y.; Li, Y.; Xie, Q.; Zhang, C.; You, S. Enhanced bioremediation of hydraulic fracturing flowback and produced water using an indigenous biosurfactant-producing bacteria Acinetobacter sp. Y2. Chem. Eng. J. 2020, 397, 125348. [Google Scholar] [CrossRef]
- Nandi, L.; Panigrahi, A.K.; Maitra, N.; Chattopadhyay, A.P.; Manna, S.K. Isolation, characterization and growth kinetics of phenol hyper-tolerant bacteria from sewage-fed aquaculture system. J. Environ. Sci. Health A 2020, 55, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Abo-Zed, A.; Yassin, M.; Phan, T. Acinetobacter junii as a rare pathogen of urinary tract infection. Urol. Case Rep. 2020, 32, 101209. [Google Scholar] [CrossRef] [PubMed]
- Kollimuttathuillam, S.; Bethel, N.; Shaaban, H. A Case of Acinetobacter junii Cavitary Pneumonia With Bacteremia in a Patient With Systemic Lupus Erythematosus. Cureus 2021, 13, e19711. [Google Scholar] [CrossRef] [PubMed]
- Hrenovic, J.; Kovacevic, D.; Ivankovic, T.; Tibljas, D. Selective immobilization of Acinetobacter junii on the natural zeolitized tuff in municipal wastewater. Colloid. Surface. B 2011, 88, 208–214. [Google Scholar] [CrossRef]
- Yang, L.; Ren, Y.X.; Liang, X.; Zhao, S.Q.; Wanga, J.P.; Xia, Z.H. Nitrogen removal characteristics of a heterotrophic nitrifier Acinetobacter junii YB and its potential application for the treatment of high-strength nitrogenous wastewater. Bioresour. Technol. 2015, 193, 227–233. [Google Scholar] [CrossRef]
- Bar-Niv, N.; Azaizeh, H.; Kuc, M.E.; Azerrad, S.; Haj-Zaroubi, M.; Menashe, O.; Kurzbaum, E. Advanced oxidation process UV-H2O2 combined with biological treatment for the removal and detoxification of phenol. J. Water Process Eng. 2022, 48, 102923. [Google Scholar] [CrossRef]
- Rehman, K.; Imran, A.; Amin, I.; Afzal, M. Enhancement of oil field-produced wastewater remediation by bacterially-augmented floating treatment wetlands. Chemosphere 2019, 217, 576–583. [Google Scholar] [CrossRef]
- Sarma, H.; Sonowal, S.; Prasad, M.N.V. Plant-microbiome assisted and biochar-amended remediation of heavy metals and polyaromatic compounds-a microcosmic study. Ecotox. Environ. Safe. 2019, 176, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wu, N.; Fan, Y. Characterization of a novel lipase and its specific foldase from Acinetobacter sp. XMZ-26. Process Biochem. 2012, 47, 643–650. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, M.; Chen, X.; Lv, P.; Luo, W.; Wang, Z.; Xu, J.; Wang, Z. Co-Expression of a Thermally Stable and Methanol-Resistant Lipase and Its Chaperone from Burkholderia cepacia G63 in Escherichia coli. Appl. Biochem. Biotech. 2021, 193, 717–729. [Google Scholar] [CrossRef]
- Jia, J.; Liu, M.; Feng, L.; Wang, Z. Comparative genomic analysis reveals the evolution and environmental adaptation of Acinetobacter johnsonii. Gene 2022, 808, 145985. [Google Scholar] [CrossRef]
- Power, J.J.; Pinheiro, F.; Pompei, S.; Kovacova, V.; Yueksel, M.; Rathmann, I.; Foerster, M.; Laessig, M.; Maier, B. Adaptive evolution of hybrid bacteria by horizontal gene transfer. Proc. Natl. Acad. Sci. USA 2021, 118, e2007873118. [Google Scholar] [CrossRef]
- Arnold, B.J.; Huang, I.T.; Hanage, W.P. Horizontal gene transfer and adaptive evolution in bacteria. Nat. Rev. Microbiol. 2022, 20, 206–218. [Google Scholar] [CrossRef]
- Chandra, P.; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef] [PubMed]
- Snellman, E.A.; Colwell, R.R. Acinetobacter lipases: Molecular biology, biochemical properties and biotechnological potential. J. Ind. Microbiol. Biot. 2004, 31, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.K.; Jagtap, S.; Priya, R.; Ramadas, K. Purification, Characterization of Alkaline Cold Active Lipase from Acinetobacter radioresistens PR8 and Development of a New Zymography Method for Lipase Detection. Protein Pept. Lett. 2018, 25, 897–907. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Investigation of Lipolytic-Secreting Bacteria from an Artificially Polluted Soil Using a Modified Culture Method and Optimization of Their Lipase Production. Microorganisms 2021, 9, 2590. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Activity of WCO-9 | Activity of ATCC 17908 | WCO-9’s Diameter of Degradation Circle | ATCC 17908’s Diameter of Degradation Circle |
|---|---|---|---|---|
| Natural oils | (U/L) | (U/L) | (mm) | (mm) |
| Corn oil | 3358 ± 156 | ND | 23 ± 0.5 | ND |
| Peanut oil | 4219 ± 183 | ND | 25 ± 0.5 | ND |
| Canola oil | 3013 ± 134 | ND | 22 ± 0.5 | ND |
| Soybean oil | 3145 ± 173 | ND | 21 ± 0.5 | ND |
| Olive oil | 2833 ± 166 | ND | 21 ± 0.5 | ND |
| ρ-NP ester | (U/L) | (U/L) | ||
| ρ-nitrophenyl decanoate (C10) | 2435 ± 177 | ND | − | − |
| ρ-nitrophenyl laurate (C12) | 2012 ± 90 | 177 ± 20 | − | − |
| ρ-nitrophenyl myristate (C14) | 1388 ± 50 | 119 ± 9 | − | − |
| ρ-nitrophenyl palmitate (C16) | 288 ± 14 | 46 ± 7 | − | − |
| ρ-nitrophenyl stearate (C18) | 161 ± 7 | 25 ± 13 | − | − |
| Strain | Assembly No. | Level | Size (Mb) | GC (%) | No. of Genes | No. of Proteins | Isolation Source |
|---|---|---|---|---|---|---|---|
| A. junii WCO-9 | CP090890.1 | complete | 3.19 | 38.6 | 3012 | 2890 | oil-contaminated soil |
| A. junii ATCC 17908 | APPX00000000.1 | scaffold | 3.36 | 38.9 | 3185 | 2947 | urine |
| A. junii CAM121 | CP068253.1 | complete | 3.36 | 38.8 | 3224 | 2898 | mouth |
| A. junii 65 | CP019041.1 | complete | 3.38 | 38.6 | 3211 | 2956 | limnetic water |
| A. junii CIP 107470 | APPS00000000.1 | scaffold | 3.78 | 38.6 | 3601 | 3302 | activated sludge plant |
| A. junii lzh-X15 | CP024632.1 | complete | 3.29 | 38.8 | 3069 | 2954 | soil |
| A. junii NIPH 182 | APPW00000000.1 | scaffold | 3.31 | 38.5 | 3174 | 3047 | phlegmon pus |
| A. junii SH205 | ACPM00000000.1 | scaffold | 3.46 | 39.0 | 3279 | 3079 | missing |
| A. junii TG19608 | AMJF00000000.1 | contig | 3.26 | 38.7 | 3108 | 2921 | urine |
| A. junii WCHAJ59 | CP028800.2 | complete | 3.35 | 38.9 | 3202 | 2968 | sewage |
| A. junii YR7 | CP059558.1 | complete | 3.39 | 38.6 | 3150 | 3023 | feces |
| Strain | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ANIb | ||||||||||||
| 1 | A. junii WCO-9 | * | 97.18 | 97.23 | 97.23 | 98.18 | 97.97 | 97.09 | 94.38 | 97.06 | 97.18 | 97.95 |
| 2 | A. junii ATCC17908 | 97.03 | * | 97.39 | 97.13 | 96.99 | 97.16 | 97.57 | 97.11 | 97.76 | 98.21 | 97.17 |
| 3 | A. junii CAM121 | 97.12 | 97.30 | * | 97.00 | 97.07 | 97.14 | 97.04 | 94.35 | 97.63 | 97.56 | 97.05 |
| 4 | A. junii CIP107470 | 96.92 | 96.76 | 96.88 | * | 96.82 | 97.13 | 96.80 | 93.89 | 96.95 | 96.96 | 96.92 |
| 5 | A. junii lzh-X15 | 98.16 | 97.06 | 97.20 | 97.17 | * | 98.13 | 97.15 | 94.80 | 97.11 | 97.21 | 98.01 |
| 6 | A. junii NIPH182 | 97.95 | 97.17 | 97.18 | 97.26 | 98.10 | * | 97.05 | 94.70 | 97.11 | 97.14 | 97.89 |
| 7 | A. junii SH205 | 97.01 | 97.46 | 97.07 | 96.95 | 97.08 | 96.98 | * | 94.12 | 97.55 | 97.43 | 96.96 |
| 8 | A. junii TG19608 | 97.00 | 100 | 96.86 | 97.38 | 96.52 | 97.51 | 96.55 | * | 96.63 | 97.45 | 96.95 |
| 9 | A. junii WCHAJ59 | 97.05 | 97.82 | 97.70 | 97.17 | 97.07 | 97.17 | 97.51 | 93.44 | * | 97.84 | 97.09 |
| 10 | A. junii 65 | 97.08 | 98.17 | 97.67 | 97.08 | 97.12 | 97.12 | 97.38 | 94.63 | 97.80 | * | 97.11 |
| 11 | A. junii YR7 | 97.94 | 97.19 | 97.15 | 97.10 | 98.01 | 97.93 | 96.94 | 94.90 | 97.05 | 97.16 | * |
| ANIm | ||||||||||||
| 1 | A. junii WCO-9 | * | 97.84 | 97.87 | 97.88 | 98.37 | 98.24 | 97.93 | 97.50 | 97.93 | 97.82 | 98.24 |
| 2 | A. junii ATCC 17908 | 97.85 | * | 98.12 | 98.04 | 97.84 | 97.82 | 98.27 | 100 | 98.35 | 98.51 | 97.87 |
| 3 | A. junii CAM121 | 97.87 | 98.13 | * | 97.94 | 97.95 | 97.92 | 98.12 | 97.40 | 98.42 | 98.17 | 97.92 |
| 4 | A. junii CIP 107470 | 97.88 | 98.04 | 97.94 | * | 97.91 | 97.91 | 98.16 | 97.59 | 98.06 | 98.02 | 97.85 |
| 5 | A. junii lzh-X15 | 98.37 | 97.83 | 97.95 | 97.92 | * | 98.32 | 97.93 | 97.61 | 97.93 | 97.89 | 98.29 |
| 6 | A. junii NIPH182 | 98.24 | 97.82 | 97.92 | 97.91 | 98.32 | * | 97.89 | 97.69 | 97.91 | 97.86 | 98.30 |
| 7 | A. junii SH205 | 97.93 | 98.26 | 98.12 | 98.16 | 97.93 | 97.89 | * | 98.39 | 98.28 | 98.31 | 97.91 |
| 8 | A. junii TG19608 | 97.50 | 100 | 97.40 | 97.59 | 97.61 | 97.69 | 98.38 | * | 98.32 | 98.50 | 97.78 |
| 9 | A. junii WCHAJ59 | 97.93 | 98.36 | 98.42 | 98.06 | 97.93 | 97.91 | 98.28 | 98.32 | * | 98.40 | 97.90 |
| 10 | A. junii 65 | 97.82 | 98.51 | 98.17 | 98.02 | 97.89 | 97.86 | 98.32 | 98.50 | 98.40 | * | 97.87 |
| 11 | A. junii YR7 | 98.24 | 97.87 | 97.92 | 97.85 | 98.29 | 98.30 | 97.91 | 97.75 | 97.90 | 97.87 | * |
| DDH | ||||||||||||
| 1 | A. junii WCO-9 | * | 78.8 | 78 | 78.8 | 85.8 | 83.8 | 79.4 | 77.9 | 79.1 | 78.5 | 84.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Fan, Q.; Zhang, Z.; Deng, Y.; Wang, L.; Dai, Q.; Wang, J.; Lin, M.; Zhou, J.; Long, Z.; et al. Biodegradation of Oil by a Newly Isolated Strain Acinetobacter junii WCO-9 and Its Comparative Pan-Genome Analysis. Microorganisms 2023, 11, 407. https://doi.org/10.3390/microorganisms11020407
Jiang S, Fan Q, Zhang Z, Deng Y, Wang L, Dai Q, Wang J, Lin M, Zhou J, Long Z, et al. Biodegradation of Oil by a Newly Isolated Strain Acinetobacter junii WCO-9 and Its Comparative Pan-Genome Analysis. Microorganisms. 2023; 11(2):407. https://doi.org/10.3390/microorganisms11020407
Chicago/Turabian StyleJiang, Shijie, Qingfeng Fan, Zeying Zhang, Yunfeng Deng, Lihong Wang, Qilin Dai, Jin Wang, Min Lin, Jian Zhou, Zhijian Long, and et al. 2023. "Biodegradation of Oil by a Newly Isolated Strain Acinetobacter junii WCO-9 and Its Comparative Pan-Genome Analysis" Microorganisms 11, no. 2: 407. https://doi.org/10.3390/microorganisms11020407
APA StyleJiang, S., Fan, Q., Zhang, Z., Deng, Y., Wang, L., Dai, Q., Wang, J., Lin, M., Zhou, J., Long, Z., He, G., & Zhou, Z. (2023). Biodegradation of Oil by a Newly Isolated Strain Acinetobacter junii WCO-9 and Its Comparative Pan-Genome Analysis. Microorganisms, 11(2), 407. https://doi.org/10.3390/microorganisms11020407