Diversity of Methanogens in Animals’ Gut



Abstract
:1. Introduction
2. Materials and Methods
2.1. Feces Samples
2.2. DNA Extraction and PCR Assays
2.3. Multispacer Sequence Typing
2.4. Phylogenetic Analyses
2.5. Statistical Analyses
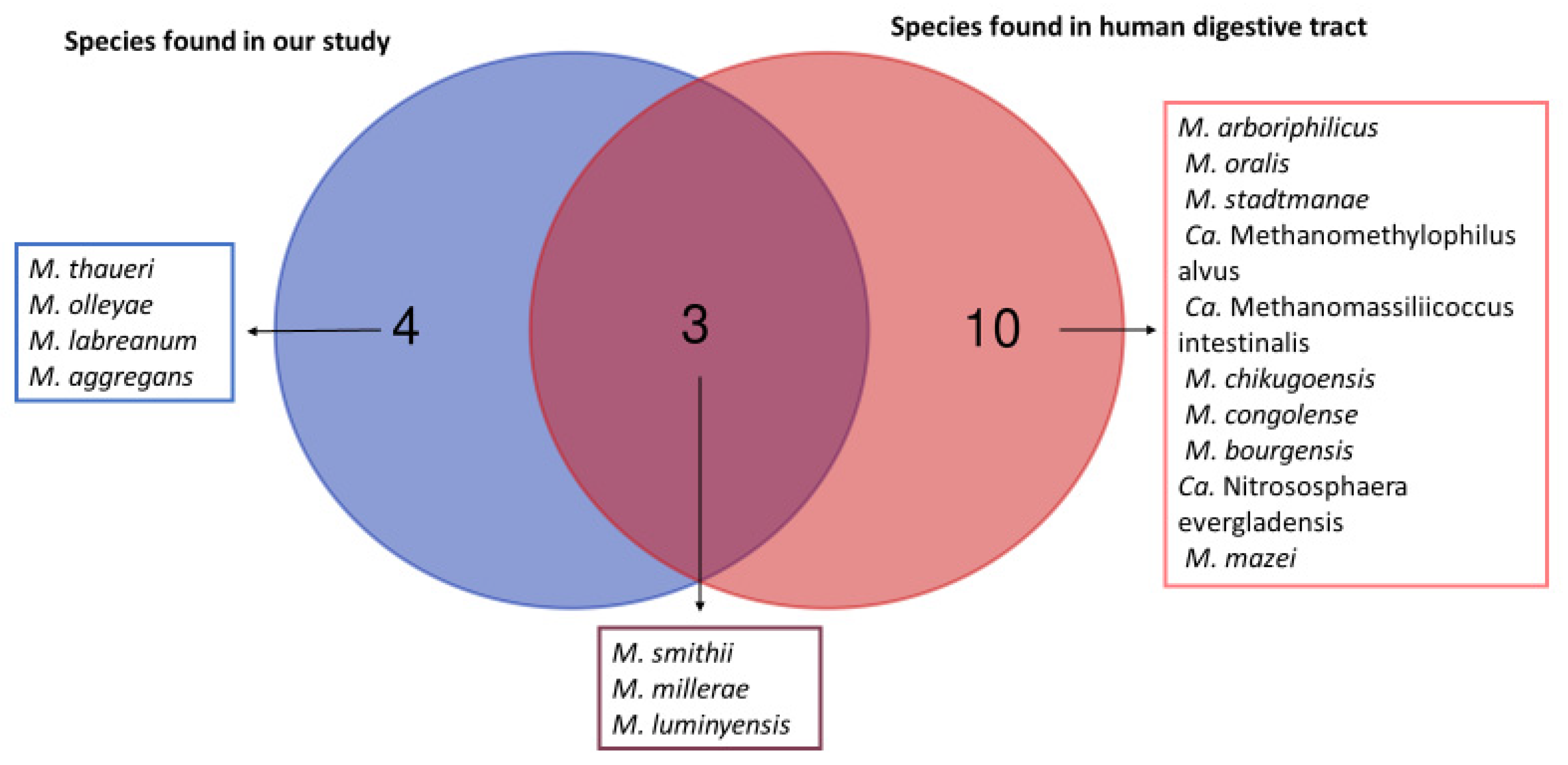
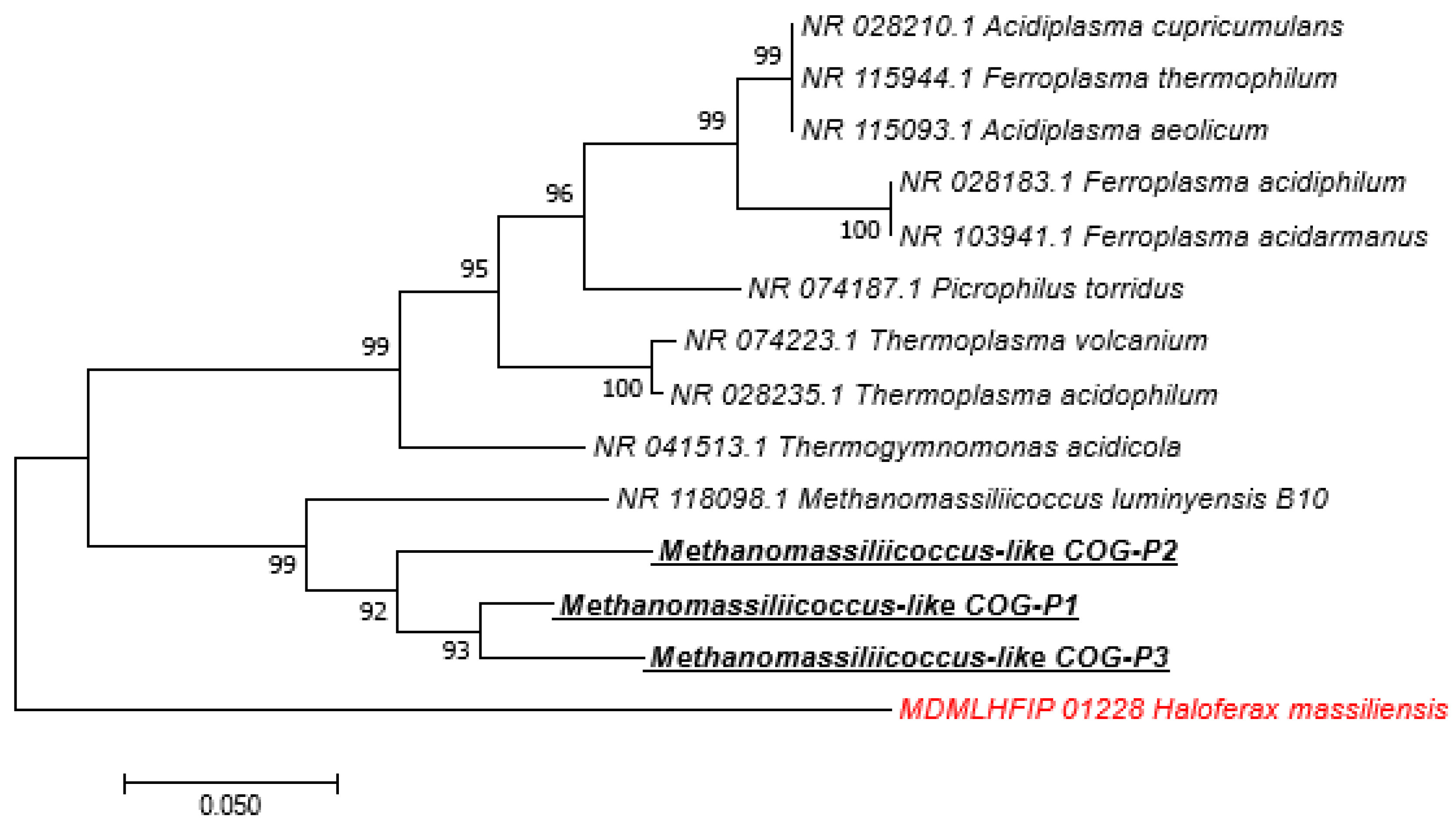
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Michelland, R.J.; Monteils, V.; Combes, S.; Cauquil, L.; Gidenne, T.; Fortun-Lamothe, L. Comparison of the archaeal community in the fermentative compartment and faeces of the cow and the rabbit. Anaerobe 2010, 16, 396–401. [Google Scholar] [CrossRef]
- Lyu, Z.; Lu, Y. Metabolic shift at the class level sheds light on adaptation of methanogens to oxidative environments. ISME J. 2018, 12, 411–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grine, G.; Terrer, E.; Boualam, M.A.; Aboudharam, G.; Chaudet, H.; Ruimy, R.; Drancourt, M. Tobacco-smoking-related prevalence of methanogens in the oral fluid microbiota. Sci. Rep. 2018, 8, 9197. [Google Scholar] [CrossRef] [PubMed]
- Sogodogo, E.; Drancourt, M.; Grine, G. Methanogens as emerging pathogens in anaerobic abscesses. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Ruaud, A.; Esquivel-Elizondo, S.; de la Cuesta-Zuluaga, J.; Waters, J.L.; Angenent, L.T.; Youngblut, N.D.; Ley, R.E. Syntrophy via interspecies H2 transfer between Christensenella and Methanobrevibacter underlies their global cooccurrence in the human gut. mBio 2020, 11, e03235-19. [Google Scholar] [CrossRef] [Green Version]
- Nkamga, V.D.; Henrissat, B.; Drancourt, M. Archaea: Essential inhabitants of the human digestive microbiota. Hum. Microbiome J. 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Guindo, C.O.; Drancourt, M.; Grine, G. Digestive tract methanodrome: Physiological roles of human microbiota-associated methanogens. Microb. Pathog. 2020, 149, 104425. [Google Scholar] [CrossRef]
- Togo, A.H.; Grine, G.; Khelaifia, S.; des Robert, C.; Brevaut, V.; Caputo, A.; Baptiste, E.; Bonnet, M.; Levasseur, A.; Drancourt, M.; et al. Culture of methanogenic archaea from human colostrum and milk. Sci. Rep. 2019, 9, 18653. [Google Scholar] [CrossRef] [Green Version]
- Grine, G.; Drouet, H.; Fenollar, F.; Bretelle, F.; Raoult, D.; Drancourt, M. Detection of Methanobrevibacter smithii in vaginal samples collected from women diagnosed with bacterial vaginosis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1643–1649. [Google Scholar] [CrossRef]
- Grine, G.; Lotte, R.; Chirio, D.; Chevalier, A.; Raoult, D.; Drancourt, M.; Ruimy, R. Co-culture of Methanobrevibacter smithii with enterobacteria during urinary infection. EBioMedicine 2019, 43, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Drancourt, M.; Nkamga, V.D.; Lakhe, N.A.; Régis, J.-M.; Dufour, H.; Fournier, P.-E.; Bechah, Y.; Michael Scheld, W.; Raoult, D. Evidence of archaeal methanogens in brain abscess. Clin. Infect. Dis. 2017, 65, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Nkamga, V.D.; Lotte, R.; Chirio, D.; Lonjon, M.; Roger, P.-M.; Drancourt, M.; Ruimy, R. Methanobrevibacter oralis detected along with Aggregatibacter actinomycetemcomitans in a series of community-acquired brain abscesses. Clin. Microbiol. Infect. 2018, 24, 207–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkamga, V.D.; Lotte, R.; Roger, P.-M.; Drancourt, M.; Ruimy, R. Methanobrevibacter smithii and Bacteroides thetaiotaomicron cultivated from a chronic paravertebral muscle abscess. Clin. Microbiol. Infect. 2016, 22, 1008–1009. [Google Scholar] [CrossRef]
- Faveri, M.; Gonçalves, L.F.H.; Feres, M.; Figueiredo, L.C.; Gouveia, L.A.; Shibli, J.A.; Mayer, M.P.A. Prevalence and microbiological diversity of Archaea in peri-implantitis subjects by 16S ribosomal RNA clonal analysis: Archaea in peri-implantitis subjects. J. Periodontal Res. 2011, 46, 338–344. [Google Scholar] [CrossRef]
- Nguyen-Hieu, T.; Khelaifia, S.; Aboudharam, G.; Drancourt, M. Methanogenic archaea in subgingival sites: A review. APMIS 2013, 121, 467–477. [Google Scholar] [CrossRef]
- Sogodogo, E.; Fellag, M.; Loukil, A.; Nkamga, V.D.; Michel, J.; Dessi, P.; Fournier, P.-E.; Drancourt, M. Nine cases of methanogenic archaea in refractory sinusitis, an emerging clinical entity. Front. Public Health 2019, 7, 38. [Google Scholar] [CrossRef]
- Drancourt, M.; Djemai, K.; Gouriet, F.; Grine, G.; Loukil, A.; Bedotto, M.; Levasseur, A.; Lepidi, H.; Bou-Khalil, J.; Khelaifia, S.; et al. Methanobrevibacter smithii archaemia in febrile patients with bacteremia, including those with endocarditis. Clin. Infect. Dis. 2020, ciaa998. [Google Scholar] [CrossRef]
- Guindo, C.O.; Terrer, E.; Chabrière, E.; Aboudharam, G.; Drancourt, M.; Grine, G. Culture of salivary methanogens assisted by chemically produced hydrogen. Anaerobe 2020, 61, 102128. [Google Scholar] [CrossRef]
- Miller, T.L. Description of Methanobrevibacter gottschalkii sp. nov., Methanobrevibacter thaueri sp. nov., Methanobrevibacter woesei sp. nov. and Methanobrevibacter wolinii sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 819–822. [Google Scholar]
- Pol, A.; Demeyer, D.I. Fermentation of methanol in the sheep rumen. Appl. Environ. Microbiol. 1988, 54, 832–834. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.H.; Kirs, M. Structure of the archaeal community of the rumen. Appl. Environ. Microbiol. 2008, 74, 3619–3625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, G.N.; Strömpl, C.; Burgess, D.M.; Skillman, L.C.; Moore, E.R.B.; Joblin, K.N. Isolation and identification of ruminal methanogens from grazing cattle. Curr. Microbiol. 2000, 40, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Grine, G.; Boualam, M.A.; Drancourt, M. Methanobrevibacter smithii, a methanogen consistently colonising the newborn stomach. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2449–2455. [Google Scholar] [CrossRef] [PubMed]
- Mihajlovski, A.; Doré, J.; Levenez, F.; Alric, M.; Brugère, J.-F. Molecular evaluation of the human gut methanogenic archaeal microbiota reveals an age-associated increase of the diversity. Environ. Microbiol. Rep. 2010, 2, 272–280. [Google Scholar] [CrossRef]
- Danielsson, R.; Schnürer, A.; Arthurson, V.; Bertilsson, J. Methanogenic population and CH4 production in swedish dairy cows fed different levels of forage. Appl. Environ. Microbiol. 2012, 78, 6172–6179. [Google Scholar] [CrossRef] [Green Version]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the human infant intestinal microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.L.; Wolin, M.J. Methanogens in human and animal intestinal Tracts. Syst. Appl. Microbiol. 1986, 7, 223–229. [Google Scholar] [CrossRef]
- Miller, T.L.; Wolin, M.J.; Zhao, H.X.; Bryant, M.P. Characteristics of methanogens isolated from bovine rumen. Appl. Environ. Microbiol. 1986, 51, 201–202. [Google Scholar] [CrossRef] [Green Version]
- Skillman, L.C.; Evans, P.N.; Strömpl, C.; Joblin, K.N. 16S rDNA directed PCR primers and detection of methanogens in the bovine rumen. Lett. Appl. Microbiol. 2006, 42, 222–228. [Google Scholar] [CrossRef]
- Zhang, Y.; Zamudio Cañas, E.M.; Zhu, Z.; Linville, J.L.; Chen, S.; He, Q. Robustness of archaeal populations in anaerobic co-digestion of dairy and poultry wastes. Bioresour. Technol. 2011, 102, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Su, Y.; Wright, A.-D.G.; Zhang, L.; Smidt, H.; Zhu, W. Lean breed landrace pigs harbor fecal methanogens at higher diversity and density than obese breed erhualian pigs. Archaea 2012, 2012, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Sanabria, E.; Goonewardene, L.A.; Wang, Z.; Zhou, M.; Moore, S.S.; Guan, L.L. Influence of sire breed on the interplay among rumen microbial populations inhabiting the rumen liquid of the progeny in beef cattle. PLoS ONE 2013, 8, e58461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, S.; Bowman, J.P.; Popovski, S.; Pimm, C.; Wright, A.-D.G. Methanobrevibacter millerae sp. nov. and Methanobrevibacter olleyae sp. nov., methanogens from the ovine and bovine rumen that can utilize formate for growth. Int. J. Syst. Evol. Microbiol. 2007, 57, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Murru, F.; Fliegerova, K.; Mura, E.; Mrázek, J.; Kopečný, J.; Moniello, G. A comparison of methanogens of different regions of the equine hindgut. Anaerobe 2018, 54, 104–110. [Google Scholar] [CrossRef]
- Wright, A.-D.G.; Williams, A.J.; Winder, B.; Christophersen, C.T.; Rodgers, S.L.; Smith, K.D. Molecular diversity of rumen methanogens from sheep in western Australia. Appl. Environ. Microbiol. 2004, 70, 1263–1270. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Kumar, S.; Lee, G.-H.; Chang, D.-H.; Rhee, M.-S.; Yoon, M.-H.; Kim, B.-C. Methanobrevibacter boviskoreani sp. nov., isolated from the rumen of Korean native cattle. Int. J. Syst. Evol. Microbiol. 2013, 63, 4196–4201. [Google Scholar] [CrossRef] [Green Version]
- Tun, H.M.; Brar, M.S.; Khin, N.; Jun, L.; Hui, R.K.-H.; Dowd, S.E.; Leung, F.C.-C. Gene-centric metagenomics analysis of feline intestinal microbiome using 454 junior pyrosequencing. J. Microbiol. Methods 2012, 88, 369–376. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Lang, H.; Yang, H. Characterization of the methanogen community in a household anaerobic digester fed with swine manure in China. Appl. Microbiol. Biotechnol. 2013, 97, 8163–8171. [Google Scholar] [CrossRef]
- Saengkerdsub, S.; Ricke, S.C. Ecology and characteristics of methanogenic archaea in animals and humans. Crit. Rev. Microbiol. 2014, 40, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Whitford, M.F.; Teather, R.M.; Forster, R.J. Phylogenetic analysis of methanogens from the bovine rumen. BMC Microbiol. 2001, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Mukumba, P.; Makaka, G.; Mamphweli, S. Anaerobic digestion of donkey dung for biogas production. S. Afr. J. Sci. 2016, 112, 1–4. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Nkamga, V.D.; Huynh, H.T.T.; Aboudharam, G.; Ruimy, R.; Drancourt, M. Diversity of human-associated methanobrevibacter smithii isolates revealed by multispacer sequence typing. Curr. Microbiol. 2015, 70, 810–815. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Dridi, B.; Henry, M.; El Khéchine, A.; Raoult, D.; Drancourt, M. High prevalence of Methanobrevibacter smithii and Methanosphaera stadtmanae detected in the human gut using an improved DNA detection protocol. PLoS ONE 2009, 4, e7063. [Google Scholar] [CrossRef]
- Dridi, B.; Raoult, D.; Drancourt, M. Archaea as emerging organisms in complex human microbiomes. Anaerobe 2011, 17, 56–63. [Google Scholar] [CrossRef]
- Sogodogo, E.; Doumbo, O.; Aboudharam, G.; Kouriba, B.; Diawara, O.; Koita, H.; Togora, S.; Drancourt, M. First characterization of methanogens in oral cavity in Malian patients with oral cavity pathologies. BMC Oral Health 2019, 19, 232. [Google Scholar] [CrossRef]
- Van de Pol, J.A.A.; van Best, N.; Mbakwa, C.A.; Thijs, C.; Savelkoul, P.H.; Arts, I.C.W.; Hornef, M.W.; Mommers, M.; Penders, J. Gut colonization by methanogenic archaea is associated with organic dairy consumption in children. Front. Microbiol. 2017, 8, 35. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Methanogens Species Obtained by Culture | Additional Species Detected by PCR-Based Methods But Not by Culture |
|---|---|
| Methanosarcina sp. Methanobacterium formicicum Methanomicrobium mobile Methanosarcina barkeri Methanobacterium bryantii Methanobrevibacter, ruminantium Methanobrevibacter millerae Methanobrevibacter olleyae Methanoculleus olentangyi Methanobrevibacter woesei Methanobrevibacter gottschalkii Methanobrevibacter thaueri Methanobrevibacter wolinii Methanobrevibacter cuticularis Methanobrevibacter curvatus Methanobrevibacter filiformi | Methanobrevibacter smithii Methanimicrococcus spp. Methanosphaera spp. Methanobacterium spp. |
| Origin of Samples | Species | Collected Number per Sample | Collection Sites |
|---|---|---|---|
| Cat | Felis silvestris catus | 105 | Marseille |
| Dog | Canis lupus | 52 | Marseille |
| Horse | Equus caballus | 89 | Marseille and Carnoux |
| Sheep | Ovis aries | 29 | Bourganeuf |
| Rabbit | Oryctolagus cuniculus | 2 | Allauch |
| Cow | Bos taurus | 57 | Bourganeuf and Allauch |
| Pig | Sus scrofa domesticus | 64 | Avignon |
| Goat | Capra aegagrus hircus | 5 | Allauch |
| Donkey | Equus asinus | 4 | Allauch |
| Animal Species | Number of Samples Analyzed | Number of Positive Samples by RT-PCR | Prevalence [IC 95%] | p-Value |
|---|---|---|---|---|
| Cat | 105 | 105 | 100.0 [96.5–100.0] | 9.9 × 10–8 |
| Dog | 52 | 41 | 78.8 [65.3–88.9] | |
| Horse | 89 | 75 | 84.4 [75.0–91.1] | |
| Sheep | 29 | 28 | 96.6 [82.2–99.9] | |
| Rabbit | 2 | 2 | 100.0 [15.8–100.0] | |
| Cow | 57 | 57 | 100.0 [93.7–100.0] | |
| Pig | 64 | 64 | 100.0 [94.4–100.0] | |
| Goat | 5 | 4 | 80.0 [28.4–99.5] | |
| Donkey | 4 | 4 | 100.0 [39.8–100.0] |
| Animal Species | Number of Samples Analyzed | Number of Positive Samples by PCR-Sequencing | Prevalence [IC 95%] | p-Value |
|---|---|---|---|---|
| Cat | 105 | 50 | 47.6 [37.8–57.6] | 1.4 × 10–12 |
| Dog | 52 | 30 | 57.7 [43.2–71.3] | |
| Horse | 89 | 24 | 27.0 [ 18.1–37.4] | |
| Sheep | 29 | 28 | 96.6 [82.2–99.9] | |
| Rabbit | 2 | 2 | 100.0 [15.8–100.0] | |
| Cow | 57 | 44 | 77.2 [64.2–87.3] | |
| Pig | 64 | 25 | 39.1 [27.1–52.1] | |
| Goat | 5 | 4 | 80.0 [28.4–99.5] | |
| Donkey | 4 | 4 | 100.0 [39.8–100.0] |
| Animal Species | Percentage > 99% | Percentage < 98.7% | Percentage < 95% |
|---|---|---|---|
| Cat | 50 | 0 | 0 |
| Dog | 30 | 0 | 0 |
| Horse | 7 | 15 | 2 |
| Sheep | 18 | 7 | 3 |
| Rabbit | 2 | 0 | 0 |
| Cow | 29 | 12 | 3 |
| Pig | 16 | 4 | 5 |
| Goat | 0 | 2 | 2 |
| Donkey | 1 | 3 | 0 |
| Genotypes | |||||||
|---|---|---|---|---|---|---|---|
| Samples | Origin | Collection Sites | Spacer 1 | Spacer 2 | Spacer 3 | Spacer 4 | Spacer Type * |
| 1 | Sheep | Bouganeuf | × | 1 | |||
| 2 | Horse | Marseille | × | 1 | |||
| 3 | Pig | Avignon | × | × | × | × | 2 |
| 4 | Pig | Avignon | × | × | × | × | 2 |
| 5 | Pig | Avignon | × | × | × | × | 2 |
| 6 | Pig | Avignon | × | × | × | × | 2 |
| 7 | Pig | Avignon | × | × | × | 3 | |
| 8 | Pig | Avignon | × | × | 4 | ||
| 9 | Pig | Avignon | × | × | 5 | ||
| 10 | Pig | Avignon | × | 1 | |||
| 11 | Pig | Avignon | × | × | × | × | 2 |
| 12 | Pig | Avignon | × | × | × | 3 | |
| 13 | Pig | Avignon | × | × | × | × | 2 |
| 14 | Pig | Avignon | × | × | × | × | 2 |
| 15 | Dog | Marseille | × | × | × | × | 2 |
| 16 | Dog | Marseille | × | × | × | × | 2 |
| 17 | Dog | Marseille | × | × | × | 3 | |
| 18 | Dog | Marseille | × | × | × | 3 | |
| 19 | Dog | Marseille | × | × | × | × | 2 |
| 20 | Dog | Marseille | × | 1 | |||
| 21 | Cat | Marseille | × | 1 | |||
| 22 | Cat | Marseille | × | 1 | |||
| 23 | Cat | Marseille | × | 1 | |||
| 24 | Cat | Marseille | × | 1 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guindo, C.O.; Davoust, B.; Drancourt, M.; Grine, G. Diversity of Methanogens in Animals’ Gut. Microorganisms 2021, 9, 13. https://doi.org/10.3390/microorganisms9010013
Guindo CO, Davoust B, Drancourt M, Grine G. Diversity of Methanogens in Animals’ Gut. Microorganisms. 2021; 9(1):13. https://doi.org/10.3390/microorganisms9010013
Chicago/Turabian StyleGuindo, Cheick Oumar, Bernard Davoust, Michel Drancourt, and Ghiles Grine. 2021. "Diversity of Methanogens in Animals’ Gut" Microorganisms 9, no. 1: 13. https://doi.org/10.3390/microorganisms9010013
APA StyleGuindo, C. O., Davoust, B., Drancourt, M., & Grine, G. (2021). Diversity of Methanogens in Animals’ Gut. Microorganisms, 9(1), 13. https://doi.org/10.3390/microorganisms9010013