The Multiple Origins of Roe Deer Populations in Western Iberia and Their Relevance for Conservation

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials & Methods
2.1. Sample Collection
2.2. Laboratory Procedures, Sequencing and Genotyping
3. Data Analysis
3.1. Genetic Structure and Diversity Assessment Using Nuclear Markers
3.2. Phylogenetic Structure and Affinities with Iberian and European Populations
4. Results
4.1. Genetic Structure and Diversity
4.2. Phylogeographic Relationships in Iberia and Europe
5. Conclusions
5.1. Western Iberia Populations—Their Structure and Origins
5.2. Management and Conservation Implications
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Holt, B.G. An Update of Wallace’s Zoogegraphic Regions of the World. Science 2013, 339, 74–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, S.; Ferrand, N. Current Perspectives in Phylogeography and the Significance of South European Refugia in the Creation and Maintenance of European Biodiversity in Phylogeography of Southern European Refugia; Springer: Dordrecht, The Netherlands, 2007; Chapter 14. [Google Scholar]
- Randi, E. Phylogeography of South European Mammals in Phylogeography of Southern European Refugia; Springer: Dordrecht, The Netherlands, 2007; Chapter 3. [Google Scholar]
- Lovari, S.; Herrero, J.; Masseti, M.; Ambarli, H.; Lorenzini, R.; Giannatos, G. Capreolus capreolus, The IUCN Red List of Threatened Species. 2016. Available online: https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T42395A22161386.en (accessed on 27 May 2020).
- Vernesi, C.; Pecchioli, E.; Caramelli, D.; Tiedemann, R.; Randi, E.; Bertorelle, G. The genetic structure of natural and reintroduced roe deer (Capreolus capreolus) populations in the Alps and central Italy, with reference to the mitochondrial DNA phylogeography of Europe. Mol. Ecol. 2002, 11, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Randi, E.; Alves, P.C.; Carranza, J.; Milosevic-Zlatanovic, S.; Sfougaris, A.; Mucci, N. Phylogeography of roe deer (Capreolus capreolus) populations: The effects of historical genetic subdivisions and recent nonequilibrium dynamics. Mol. Ecol. 2004, 13, 3071–3083. [Google Scholar] [CrossRef] [PubMed]
- Royo, L.J.; Pajares, G.; Álvarez, I.; Fernández, I.; Goyache, F. Genetic variability and differentiation in Spanish roe deer (Capreolus capreolus): A phylogeographic reassessment within the European framework. Mol. Phylogenet. Evol. 2007, 42, 47–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, G.; Vernesi, C.; Vicario, S.; Pecchioli, E.; Caccone, A.; Bertorelle, G.; Sbordoni, V. Mitochondrial DNA variation in roe deer (Capreolus capreolus) from Italy: Evidence of admixture in one of the last C. c. italicus pure populations from central-southern Italy. Ital. J. Zool. 2009, 76, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Mucci, N.; Mattucci, F.; Randi, E. Conservation of threatened local gene pools: Landscape genetics of the Italian roe deer (Capreolus c. italicus) populations. Evol. Ecol. Res. 2012, 14, 897–920. [Google Scholar]
- Biosa, D.; Scandura, M.; Tagliavini, J.; Luccarini, S.; Mattioli, L.; Apollonio, M. Patterns of genetic admixture between roe deer of different origin in central Italy. J. Mammal. 2015, 96, 827–838. [Google Scholar] [CrossRef] [Green Version]
- Tsaparis, D.; Sotiropoulos, K.; Legakis, A.; Kotoulas, G.; Kasapidis, P. New phylogeographic insights support the distinctiveness and conservation value of the little-known Greek roe deer populations. Mamm. Biol. 2019, 96, 23–27. [Google Scholar] [CrossRef]
- Hewitt, G.M. Post-glacial recolonization of Europe. Biol. J. Linn. Soc. Lond. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Hewitt, G.M. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Lorenzini, R.; Lovari, S. Genetic diversity and phylogeography of the European roe deer: The refuge area theory revisited. Biol. J. Linn. Soc. Lond. 2006, 88, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Meiri, M.; Lister, A.M.; Higham, T.F.G.; Stewart, J.R.; Straus, L.G.; Obermaier, H.; Morales, M.R.G.; Marín-Arroyo, A.B.; Barnes, I. Late-glacial recolonization and phylogeography of European red deer (Cervus elaphus L.). Mol. Ecol. 2013, 22, 4711–4722. [Google Scholar] [CrossRef]
- Scandura, M.; Iacolina, L.; Apollonio, M. Genetic diversity in the European wild boar Sus scrofa: Phylogeography, population structure and wild × domestic hybridization. Mamm. Rev. 2011, 41, 125–137. [Google Scholar] [CrossRef]
- Veličković, N.; Ferreira, E.; Djan, M.; Ernst, M.; Vidaković, D.O.; Monaco, A.; Fonseca, C. Demographic history, current expansion and future management challenges of wild boar populations in the Balkans and Europe. Heredity 2016, 117, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Taberlet, P.; Fumagalli, L.; Hausser, J. Chromosomal versus mitochondrial DNA evolution: Tracking the evolutionary history of the southwestern European populations of the Sorex araneus group (Mammalia, Insectivora). Evolution 1994, 48, 623–636. [Google Scholar] [CrossRef]
- Branco, M.; Ferrand, N.; Monnerot, M. Phylogeography of the European rabbit (Oryctolagus cuniculus) in the Iberian Peninsula inferred from RFLP analysis of the cytochrome b gene. Heredity 2000, 85, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Frati, F.; Hartl, G.B.; Lovari, S.; Delibes, M.; Markov, G. Quaternary radiation and genetic structure of the red fox Vulpes vulpes in the Mediterranean Basin, as revealed by allozymes and mitochondrial DNA. J. Zool. 1998, 245, 43–51. [Google Scholar] [CrossRef]
- Carranza, J.; Salinas, M.; de Andrés, D.; Pérez-González, J. Iberian red deer: Paraphyletic nature at mtDNA but nuclear markers support its genetic identity. Ecol. Evol. 2016, 6, 905–922. [Google Scholar] [CrossRef] [Green Version]
- Lorenzini, R.; San José, C.; Braza, F.; Aragón, S. Genetic differentiation and phylogeography of roe deer in Spain, as suggested by mitochondrial DNA and microsatellite analysis. Ital. J. Zool. 2003, 70, 89–99. [Google Scholar] [CrossRef]
- Ferreira, E.; Souto, L.; Soares, A.M.; Fonseca, C. Genetic structure of the wild boar population in Portugal: Evidence of a recent bottleneck. Mamm. Biol. 2009, 74, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Vingada, J.; Fonseca, C.; Cancela, J.; Ferreira, J.; Eira, C. Ungulates and Their Management in Portugal; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Torres, R.T.; Miranda, J.; Carvalho, J.; Fonseca, C. Expansion and current status of roe deer (Capreolus capreolus) at the edge of its distribution in Portugal. Ann. Zool. Fennici 2015, 52, 339–352. [Google Scholar] [CrossRef]
- Bruford, M.W.; Hanotte, O.; Brookfield, J.F.Y.; Burke, T. Single-Locus and DNA Fingerprinting. In Molecular Genetic Analyses of Populations. A Pratical Approach; IRL Press: Oxford, UK, 1992; pp. 225–269. [Google Scholar]
- Douzery, E.; Randi, E. The mitochondrial control region of Cervidae: Evolutionary patterns and phylogenetic content. Mol. Biol. Evol. 1997, 14, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Gen. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Var Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Slatkin, M. A measure of population subdivision based on microsatellite allele frequencies. Genetics 1995, 139, 457–462. [Google Scholar]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinf. App. Note 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Garza, J.C.; Williamson, E.G. Detection of reduction in population size using data from microsatellite loci. Mol. Ecol. 2001, 10, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Sundqvist, L.; Keenan, K.; Zackrisson, M.; Prodöhl, P.; Kleinhans, D. Directional genetic differentiation and relative migration. Ecol. Evol. 2016, 6, 3461–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcala, N.; Goudet, J.; Vuilleumier, S. On the transition of genetic differentiation from isolation to panmixia: What we can learn from GST and D. Theor. Pop. Biol. 2014, 93, 75–84. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Sommer, R.S.; Fahlke, J.M.; Scmolcke, U.; Benecke, N.; Zachos, F.E. Quaternary history of the European roe deer Capreolus capreolus. Mamm. Rev. 2009, 29, 1–16. [Google Scholar] [CrossRef]
- Cabrera, A. Un Nuevo Cérvido Español. Boletin de la Real Sociedad Española de Historia Natural. Tomo XVI; Museo Nacional de Ciencias Naturales: Madrid, Spain, 1916; pp. 175–176. [Google Scholar]
- Aragón, S.; Braza, F.; San José, C. Caracterítisticas morfológicas de los corzos (Capreolus capreolus) de las sierras de Cádiz- Maálaga. Acta Vertebr. 1995, 22, 51–64. [Google Scholar]
- Coulon, A.; Cosson, J.F.; Morellet, N.; Angibault, J.M.; Cargnelutti, B.; Galan, M.; Aulagnier, S.; Hewison, A.J.M. Dispersal is not female biased in a resource defence mating ungulate, the European roe deer. Proc. R. Soc. B 2006, 273, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnot, N.; Gaillard, J.M.; Coulon, A.; Galan, M.; Cosson, J.F.; Delorme, D.; Klein, F.; Hewison, A.J.M. Difference between the sexes in fine-scale spatial genetic structure of roe deer. PLoS ONE 2010, 5, e14436. [Google Scholar] [CrossRef] [Green Version]
- Rosell, C.; Carretero, M.A.; Cahil, S.; Pasquina, A. Seguiemento de una reintruccíon de corzo (Capreolus capreolus) em ambiente mediterráneo. Dispersíon y area de campeo. Acta Vertebr. 1996, 23, 109–122. [Google Scholar]
- Veličković, N.; Djan, M.; Ferreira, E.; Stergar, M.; Obreht, D.; Maletić, V.; Fonseca, C. From north to south and back: The role of the Balkans and other southern peninsulas in the recolonization of Europe by wild boar. J. Biogeogr. 2015, 42, 716–728. [Google Scholar] [CrossRef]
- Andersen, R.; Duncan, P.; Linnell, J.D. The European Roe Deer: The Biology of Success; Scandinavian University Press: Oslo, Norway, 1998; Volume 376. [Google Scholar]
- Ducros, D.; Morellet, N.; Patin, R.; Atmeh, K.; Debeffe, L.; Cargnelutti, B.; Chaval, Y.; Lourtet, B.; Coulon, A.; Hewison, A.J.M. Beyond dispersal versus philopatry? Alternative behavioural tactics of juvenile roe deer in a heterogeneous landscape. Oikos 2020, 129, 81–92. [Google Scholar] [CrossRef]
- Hewitt, G.M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. Lond. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Sommer, R.S.; Nadachowski, A. Glacial refugia of mammals in Europe: Evidence from fossil records. Mamm. Rev. 2006, 36, 251–265. [Google Scholar] [CrossRef]
- Gregório, I.; Barros, T.; Pando, D.; Morante, J.; Fonseca, C.; Ferreira, E. Paths for colonization or exodus? New insights from the brown bear (Ursus arctos) population of the Cantabrian Mountains. PLoS ONE 2020, 15, e0227302. [Google Scholar] [CrossRef]
- Sgrò, C.M.; Lowe, A.J.; Hoffmann, A.A. Building evolutionary resilience for conserving biodiversity under climate change. Evol. Appl. 2011, 4, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Randi, E. Management of wild ungulate populations in Italy: Captive-breeding, hybridization and genetic consequences of translocations. Vet. Res. Commun. 2005, 29, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Carranza, J.; Martínez, J.G.; Sánchez-Prieto, C.B.; Fernández-García, J.L.; Sánchez-Fernández, B.; Álvarez-Álvarez, R.; Valencia, J.; Alarcos, S. Game species: Extinction hidden by census numbers. Anim. Biodivers. Conserv. 2003, 26, 81–84. [Google Scholar]
- Laikre, L.; Schwartz, M.K.; Waples, R.S.; Ryman, N.; Ge, M.W.G. Compromising genetic diversity in the wild: Unmonitored large-scale release of plants and animals. Trends Ecol. Evol. 2010, 25, 520–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
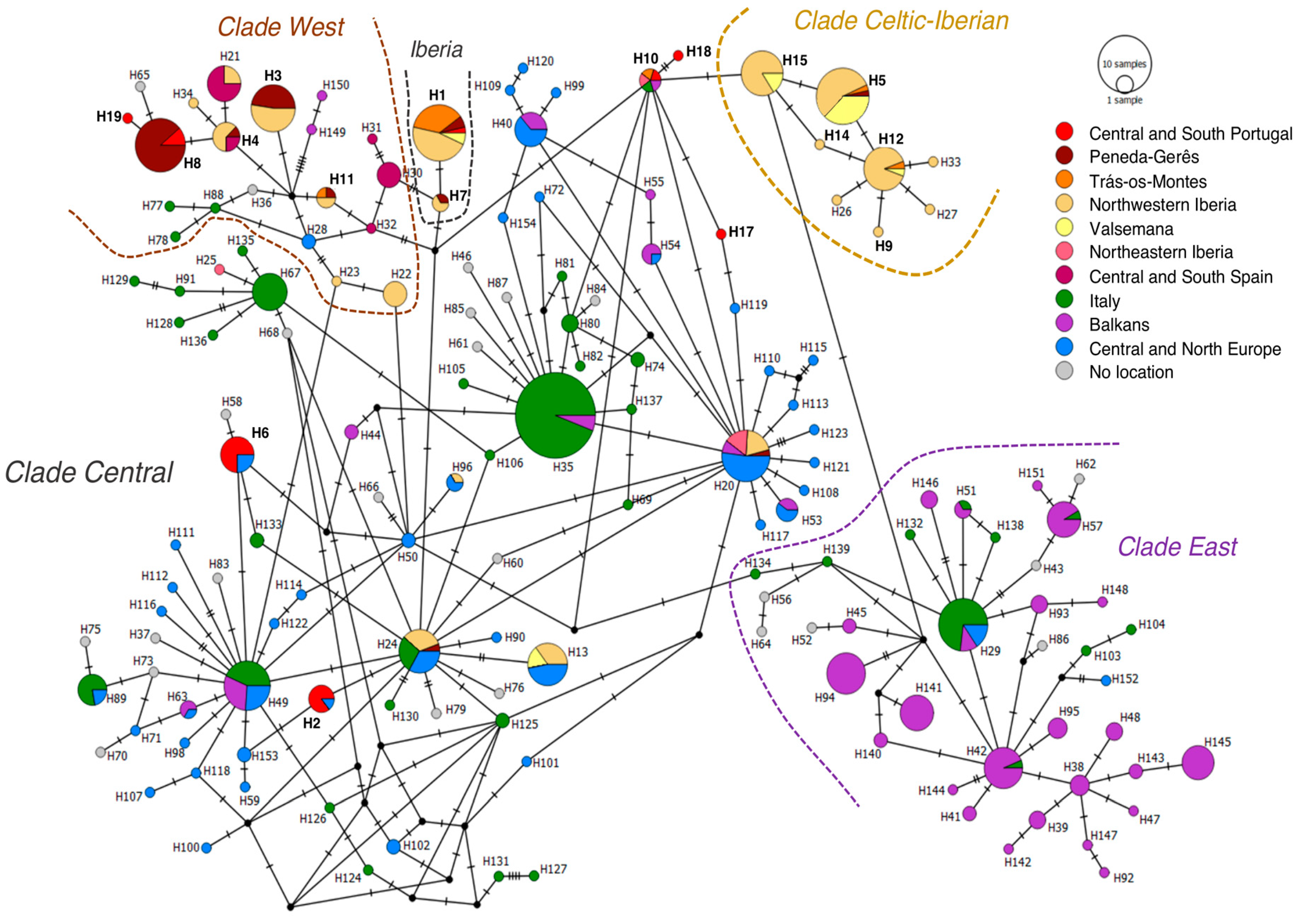{kind=link}
| Central & South Portugal | Peneda Gerês | Trás-os-Montes | NW Spain | Valsemana | |
|---|---|---|---|---|---|
| Central & South Portugal | 0.012 | 0.011 | 0.012 | 0.011 | |
| Peneda Gerês | 0.170 | 0.014 | 0.014 | 0.017 | |
| Trás-os-Montes | 0.091 | 0.105 | 0.011 | 0.011 | |
| North-western Spain | 0.084 | 0.069 | 0.051 | 0.011 | |
| Valsemana | 0.121 | 0.117 | 0.065 | 0.033 |
| Population | Nuclear DNA (Microsatellites) | Mitochondrial DNA (D-loop) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | HOBS | HEXP | Alleles (Average Values) | M | FIS | Genetic Diversity | N | H | h | Polymorphic Sites | Π | k | ||||
| Na | Neffect | Npriv | Range | |||||||||||||
| CS Portugal | 20 | 0.626 | 0.662 (2) | 6.67 | 4.20 | 0.67 | 18.6 | 0.731 | 0.098 | 0.661 | 23 | 8 | 0.787 | 14 | 0.0081 | 3.91 |
| Peneda Gerês | 33 | 0.545 | 0.580 (3) | 4.56 | 2.74 | 0.44 | 14.7 | 0.622 | 0.067 | 0.547 | 34 | 5 | 0.524 | 10 | 0.0049 | 2.35 |
| Trás-os-Montes | 14 | 0.647 | 0.728 (0) | 5.70 | 3.60 | 0.22 | 15.9 | 0.709 | 0.053 | 0.573 | 15 | 5 | 0.476 | 10 | 0.0055 | 2.63 |
| NW Spain | 22 | 0.625 | 0.671 (0) | 6.00 | 3.66 | 0.44 | 16.9 | 0.707 | 0.032 | 0.632 | 22 | 10 | 0.905 | 17 | 0.0116 | 5.55 |
| Valsemana | 19 | 0.560 | 0.653 (2) | 6.00 | 3.60 | 0.56 | 18.0 | 0.722 | 0.154 | 0.647 | 17 | 5 | 0.743 | 10 | 0.0065 | 3.12 |
| Haplotype | Central & South Portugal | Peneda Gerês | Trás-os-Montes | North-western Iberia | Valsemana | ||
|---|---|---|---|---|---|---|---|
| Alentejo | Beiras | Asturias | Galicia | ||||
| Clade Central | |||||||
| H2 | 1 | 5 | |||||
| H6 | 7 | 2 | |||||
| H10 | 1 | ||||||
| H13 | 1 | 1 | 3 | ||||
| H16 | 1 | ||||||
| H17 | 1 | ||||||
| H18 | 1 | ||||||
| Clade Central (Iberian subclade) | |||||||
| H1 | 1 | 1 | 11 | 1 | 4 | 2 | |
| H7 | 1 | 2 | |||||
| Clade West | |||||||
| H3 | 9 | 1 | 3 | ||||
| H4 | 1 | 1 | |||||
| H8 | 2 | 1 | 22 | ||||
| H11 | 1 | 1 | |||||
| H19 | 1 | ||||||
| Clade Celtic-Iberian | |||||||
| H5 | 1 | 1 | 1 | 8 | |||
| H9 | 1 | ||||||
| H12 | 1 | 2 | 1 | 1 | |||
| H14 | 1 | ||||||
| H15 | 3 | ||||||
| Total | 13 | 10 | 34 | 15 | 7 | 15 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros, T.; Ferreira, E.; Rocha, R.G.; Brotas, G.; Carranza, J.; Fonseca, C.; Torres, R.T. The Multiple Origins of Roe Deer Populations in Western Iberia and Their Relevance for Conservation. Animals 2020, 10, 2419. https://doi.org/10.3390/ani10122419
Barros T, Ferreira E, Rocha RG, Brotas G, Carranza J, Fonseca C, Torres RT. The Multiple Origins of Roe Deer Populations in Western Iberia and Their Relevance for Conservation. Animals. 2020; 10(12):2419. https://doi.org/10.3390/ani10122419
Chicago/Turabian StyleBarros, Tânia, Eduardo Ferreira, Rita Gomes Rocha, Gonçalo Brotas, Juan Carranza, Carlos Fonseca, and Rita Tinoco Torres. 2020. "The Multiple Origins of Roe Deer Populations in Western Iberia and Their Relevance for Conservation" Animals 10, no. 12: 2419. https://doi.org/10.3390/ani10122419
APA StyleBarros, T., Ferreira, E., Rocha, R. G., Brotas, G., Carranza, J., Fonseca, C., & Torres, R. T. (2020). The Multiple Origins of Roe Deer Populations in Western Iberia and Their Relevance for Conservation. Animals, 10(12), 2419. https://doi.org/10.3390/ani10122419