Color of Colon Content of Normal and Intrauterine Growth-Restricted Weaned Piglets Is Associated with Specific Microbial Taxa and Physiological Parameters

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Design
2.3. Inspection of Colon Content Color
2.4. Blood and Tissue Sampling
2.5. Flow Cytomtery and Cytokine Analysis
2.6. Sampling of Colon Content
2.7. Extraction of Total Colon Content DNA
2.8. 16S rRNA Gene Library Preparation
2.9. High Throughput Sequencing and Data Integration
2.10. Statistical Analysis
3. Results
3.1. Production Results
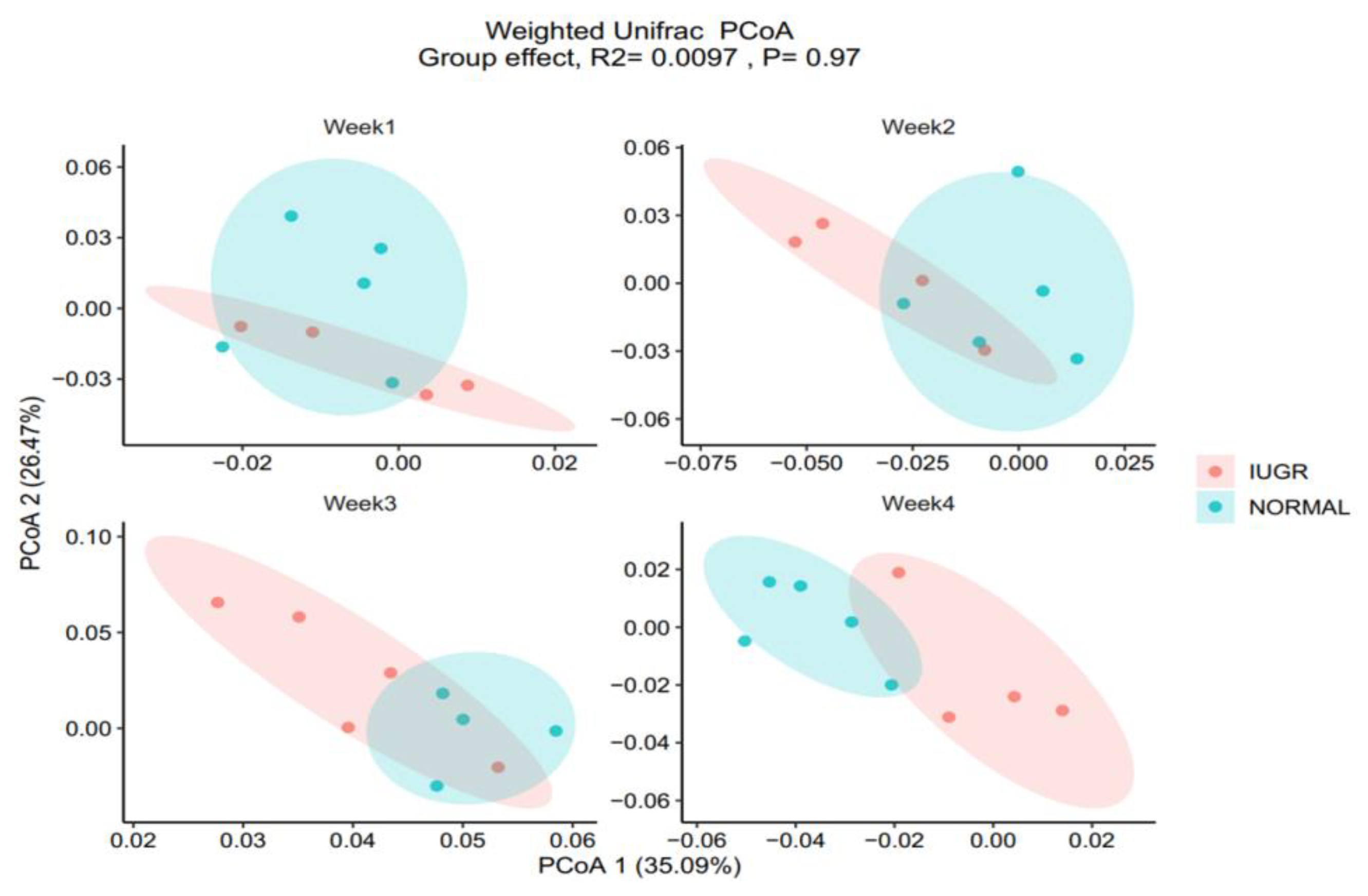
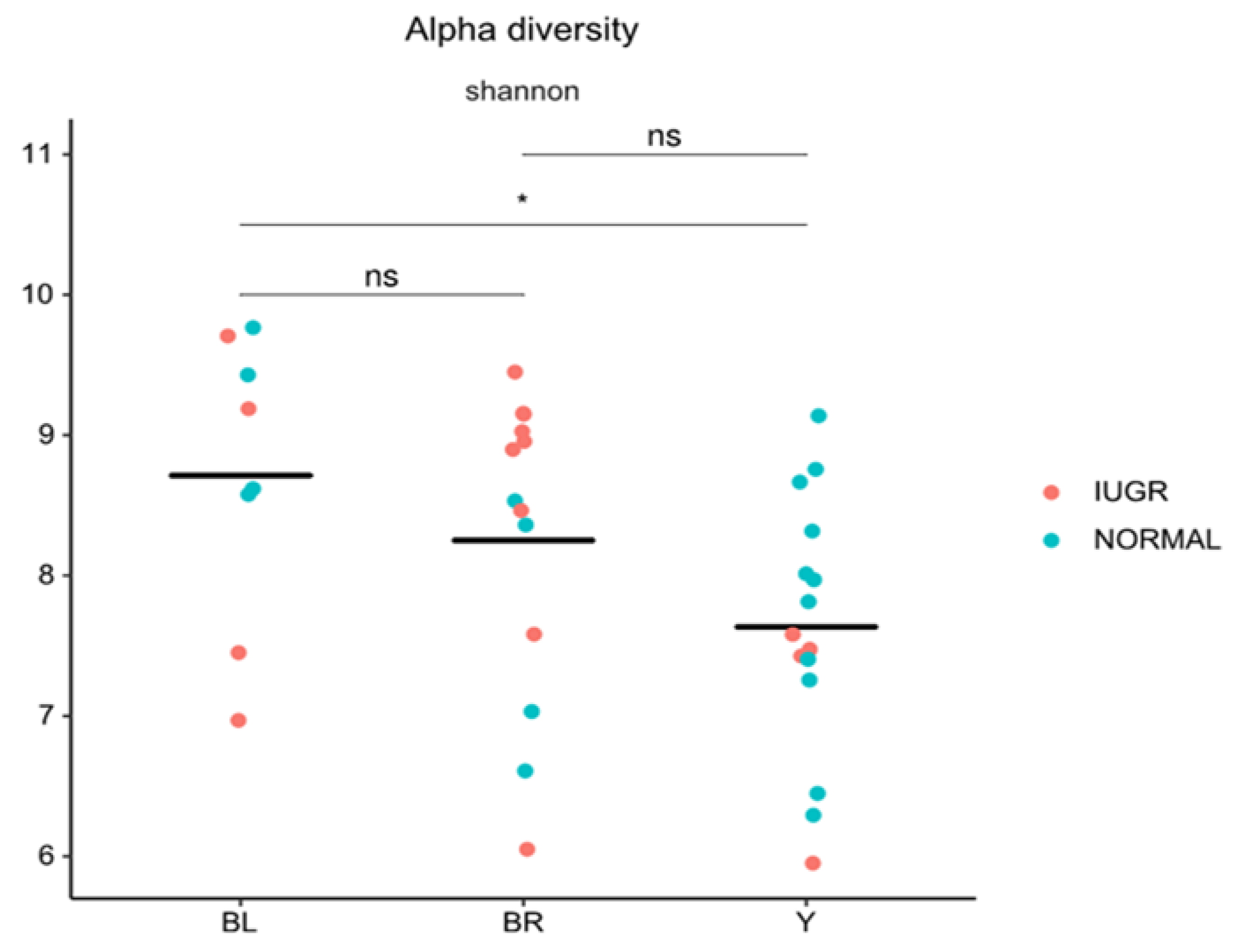
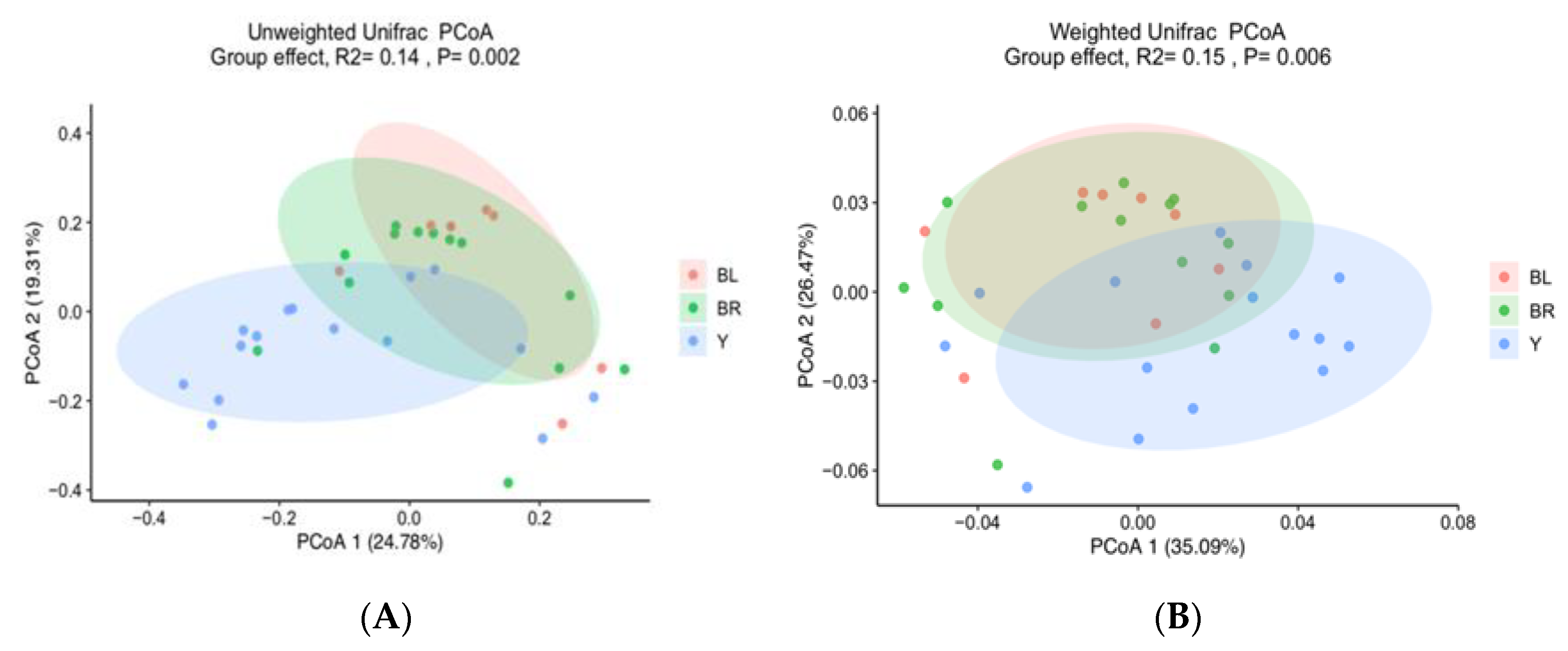
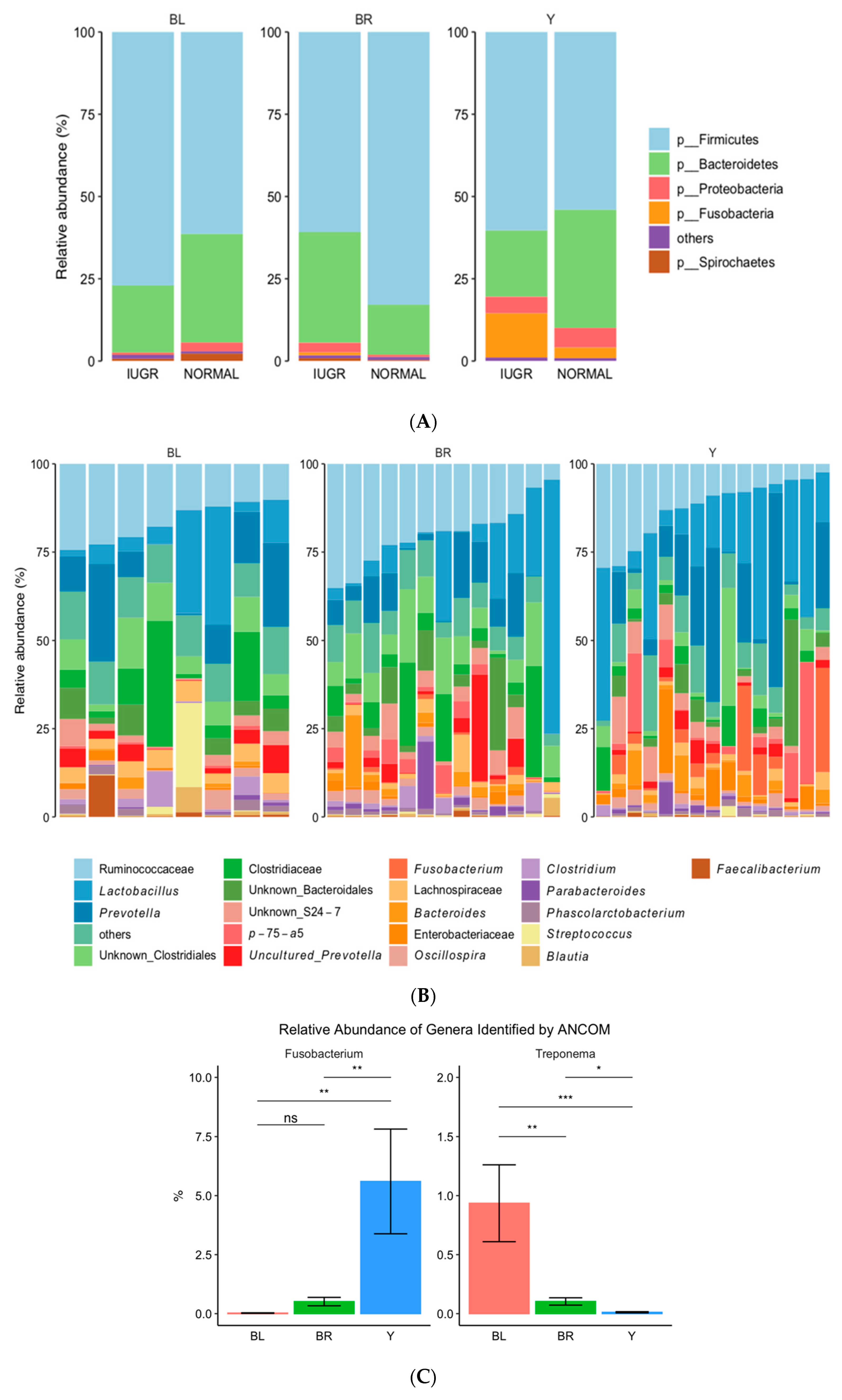
3.2. Gut Microbiota (GM)
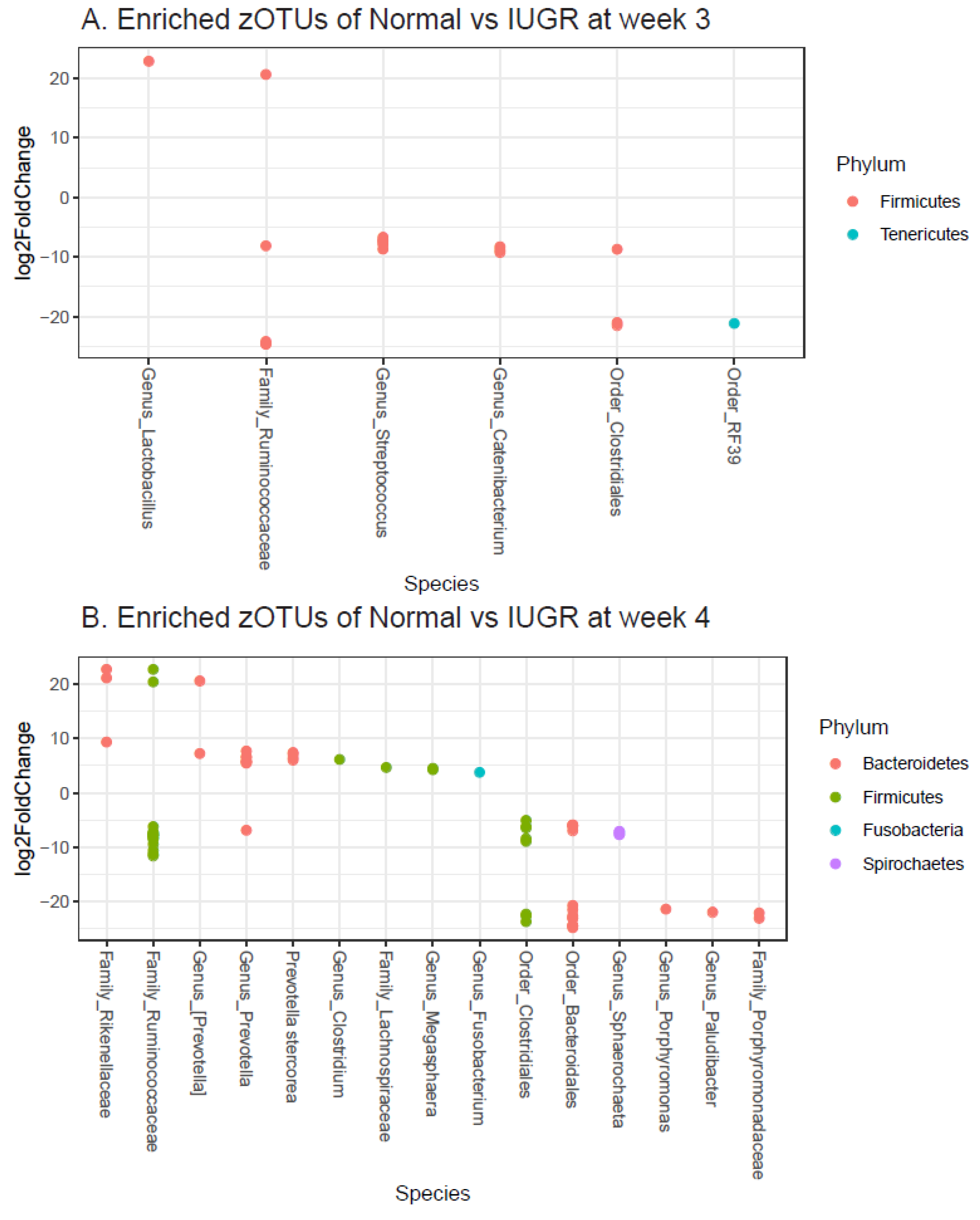
3.3. Colon Color and GM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alexander, B.T. Placental Insufficiency Leads to Development of Hypertension in Growth-Restricted Offspring. Hypertension 2003, 41, 457–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, P.Y.L.; Morris, J.M.; Leslie, G.I.; Kelly, P.J.; Gallery, E.D.M. The Long-Term Effects of Prematurity and Intrauterine Growth Restriction on Cardiovascular, Renal, and Metabolic Function. Int. J. Pediatr. 2010, 2010, 280402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatford, K.L.; Simmons, R.A.; De Blasio, M.J.; Robinson, J.S.; Owens, J.A. Review: Placental Programming of Postnatal Diabetes and Impaired Insulin Action after IUGR. Placenta 2010, 31, S60–S65. [Google Scholar] [CrossRef] [Green Version]
- Amdi, C.; Jensen, L.L.; Oksbjerg, N.; Hansen, C.F. Supplementing newborn intrauterine growth restricted piglets with a bolus of porcine colostrum raises rectal temperatures one degree Celsius. J. Anim. Sci. 2017, 95, 2968–2976. [Google Scholar] [CrossRef]
- Che, L.; Hu, L.; Zhou, Q.; Peng, X.; Liu, Y.; Luo, Y.; Fang, Z.; Lin, Y.; Xu, S.; Feng, B.; et al. Microbial insight into dietary protein source affects intestinal function of pigs with intrauterine growth retardation. Eur. J. Nutr. 2020, 59, 327–344. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.; Wallace, J.; Spencer, T. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Foxcroft, G.R.; Dixon, W.T.; Novak, S.; Putman, C.T.; Town, S.C.; Vinsky, M.D.A. The biological basis for prenatal programming of postnatal performance in pigs. J. Anim. Sci. 2006, 84, E105–E112. [Google Scholar] [CrossRef]
- Aucott, S.W.; Donohue, P.K.; Northington, F.J. Increased Morbidity in Severe Early Intrauterine Growth Restriction. J. Perinatol. 2004, 24, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Lynegaard, J.C.; Hansen, C.F.; Kristensen, A.R.; Amdi, C. Body composition and organ development of intra-uterine growth restricted pigs at weaning. Animal 2019. [Google Scholar] [CrossRef]
- Bernstein, I.M.; Horbar, J.D.; Badger, G.J.; Ohlsson, A.; Golan, A. Morbidity and mortality among very-low-birth-weight neonates with intrauterine growth restriction. Am. J. Obstet. Gynecol. 2000, 182, 198–206. [Google Scholar] [CrossRef]
- Manogura, A.C.; Turan, O.; Kush, M.L.; Berg, C.; Bhide, A.; Turan, S.; Moyano, D.; Bower, S.; Nicolaides, K.H.; Galan, H.L.; et al. Predictors of necrotizing enterocolitis in preterm growth-restricted neonates. Am. J. Obstet. Gynecol. 2008, 198, 638.e631–638.e635. [Google Scholar] [CrossRef] [PubMed]
- Che, L.; Thymann, T.; Bering, S.B.; Le Huërou-Luron, I.; D’Inca, R.; Zhang, K.; Sangild, P.T. IUGR does not predispose to necrotizing enterocolitis or compromise postnatal intestinal adaptation in preterm pigs. Pediatric Res. 2010, 67, 54–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fança-Berthon, P.; Hoebler, C.; Mouzet, E.; David, A.; Michel, C. Intrauterine Growth Restriction Not Only Modifies the Cecocolonic Microbiota in Neonatal Rats But Also Affects Its Activity in Young Adult Rats. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ma, C.; Xie, P.; Zhu, Q.; Wang, X.; Yin, Y.; Kong, X. Gut Microbiota of Newborn Piglets with Intrauterine Growth Restriction have Lower Diversity and Different Taxonomic Abundances. J. Appl. Microbiol. 2019, 127, 354–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Li, N.; Liu, C.; Li, T.; Wang, W.; Jiang, L.; Li, Z.; Han, D.; Tao, S.; Wang, J. Characteristics of the gut microbiota colonization, inflammatory profile, and plasma metabolome in intrauterine growth restricted piglets during the first 12 hours after birth. J. Microbiol. 2019, 57, 748–758. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Tito, R.Y.; Joossens, M.; Raes, J. Stool consistency is strongly associated with gut microbiota richness and composition, enterotypes and bacterial growth rates. Gut 2016, 65, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-M.; Chang, M.-H.; Du, J.-C.; Lin, C.-C.; Chen, A.-C.; Lee, H.-C.; Lau, B.-H.; Yang, Y.-J.; Wu, T.-C.; Chu, C.-H.; et al. Screening for biliary atresia by infant stool color card in Taiwan. Pediatrics 2006, 117, 1147–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.-H.; Yokoyama, K.; Mizuta, K.; Tsuchioka, T.; Kudo, T.; Sasaki, H.; Nio, M.; Tang, J.; Ohkubo, T.; Matsui, A. Stool Color Card Screening for Early Detection of Biliary Atresia and Long-Term Native Liver Survival: A 19-Year Cohort Study in Japan. J. Pediatr. 2015, 166, 897–902.e891. [Google Scholar] [CrossRef] [PubMed]
- Lien, T.-H.; Chang, M.-H.; Wu, J.-F.; Chen, H.-L.; Lee, H.-C.; Chen, A.-C.; Tiao, M.-M.; Wu, T.-C.; Yang, Y.-J.; Lin, C.-C.; et al. Effects of the infant stool color card screening program on 5-year outcome of biliary atresia in taiwan. Hepatology 2011, 53, 202–208. [Google Scholar] [CrossRef]
- Hales, J.; Moustsen, V.A.; Nielsen, M.B.F.; Hansen, C.F. Individual physical characteristics of neonatal piglets affect preweaning survival of piglets born in a noncrated system. J. Anim. Sci. 2013, 91, 4991–5003. [Google Scholar] [CrossRef] [Green Version]
- Chevaux, E.; Sacy, A.; Le Treut, Y.; Martineau, G. IntraUterine Growth Retardation (IUGR): Morphological and behavioural description. Proc. 21st Ipvs Congr. Vanc. Can. 2010, 84, 209. [Google Scholar]
- Tybirk, P.; Sloth, N.M.; Jørgensen, L.J.S. Normer for næringsstoffer. In Nutrient Requirement Standards, 24th ed.; SEGES Pig Research Centre: Copenhagen, Denmark, 2016; In Danish. [Google Scholar]
- Bekkali, N.; Hamers, S.L.; Reitsma, J.B.; Van Toledo, L.; Benninga, M.A. Infant Stool Form Scale: Development and Results. J. Pediatr. 2009, 154, 521–526.e521. [Google Scholar] [CrossRef]
- Amdi, C.; Lynegaard, J.C.; Thymann, T.; Williams, A.R. Intrauterine growth restriction in piglets alters blood cell counts and impairs cytokine responses in peripheral mononuclear cells 24 days post-partum. Sci. Rep. 2020, 10, 4683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovreås, L.; Forney, L.; Daae, F.L.; Torsvik, V. Distribution of bacterioplankton in meromictic Lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 1997, 63, 3367–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krych, Ł.; Kot, W.; Bendtsen, K.M.B.; Hansen, A.K.; Vogensen, F.K.; Nielsen, D.S. Have you tried spermine? A rapid and cost-effective method to eliminate dextran sodium sulfate inhibition of PCR and RT-PCR. J. Microbiol. Methods 2018, 144, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016, 081257. [Google Scholar]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ—J. Life Environ. Sci. 2016, 4, e2584. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nature 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Fischer, S.; Kumar, N.; Clavel, T. Rhea: A transparent and modular R pipeline for microbial profiling based on 16S rRNA gene amplicons. PeerJ—J. Life Environ. Sci. 2017, 5, e2836. [Google Scholar] [CrossRef] [Green Version]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.-R.; Shin, J.; Lee, J.H.; Kang, B.N.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.E. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Amdi, C.; Krogh, U.; Flummer, C.; Oksbjerg, N.; Hansen, C.F.; Theil, P.K. Intrauterine growth restricted piglets defined by their head shape ingest insufficient amounts of colostrum. J. Anim. Sci. 2013, 91, 5605–5613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, G.; Ma, S.; Zhu, Z.; Su, Y.; Zoetendal, E.G.; Mackie, R.; Liu, J.; Mu, C.; Huang, R.; Smidt, H. Age, introduction of solid feed and weaning are more important determinants of gut bacterial succession in piglets than breed and nursing mother as revealed by a reciprocal crossrocaltsnim model. Environ. Microbiol. 2016, 18, 1566–1577. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.A.B.; Chae, J.-P.; Balolong, M.P.; Kim, H.B.; Kang, D.-K. Assessment of fecal bacterial diversity among healthy piglets during the weaning transition. J. Gen. Appl. Microbiol. 2014, 60, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, C.; Hales, J.; Amdi, C.; Moustsen, V. Intrauterine growth-restricted piglets defined by their head shape have impaired survival and growth during the suckling period. Anim. Prod. Sci. 2019, 59, 1056–1062. [Google Scholar] [CrossRef]
- Bruininx, E.; Binnendijk, G.; Van der Peet-Schwering, C.; Schrama, J.; Den Hartog, L.; Everts, H.; Beynen, A. Effect of creep feed consumption on individual feed intake characteristics and performance of group-housed weanling pigs. J. Anim. Sci. 2002, 80, 1413–1418. [Google Scholar] [CrossRef] [Green Version]
- Che, C.; Pang, X.; Hua, X.; Zhang, B.; Shen, J.; Zhu, J.; Wei, H.; Sun, L.; Chen, P.; Cui, L. Effects of Human Fecal Flora on Intestinal Morphology and Mucosal Immunity in Human Floraitygy anima Piglet. Scand. J. Immunol. 2009, 69, 223–233. [Google Scholar] [CrossRef]
- Duman, N.; Utkutan, S.; Ozkan, H.; Ozdoğan, S. Are the stool characteristics of preterm infants affected by infant formulas? Turk. J. Pediatr. 2000, 42, 138–144. [Google Scholar]
- Hyams, J.S.; Treem, W.R.; Etienne, N.L.; Weinerman, H.; MacGilpin, D.; Hine, P.; Choy, K.; Burke, G. Effect of infant formula on stool characteristics of young infants. Pediatrics 1995, 95, 50–54. [Google Scholar] [PubMed]
- De Witte, C.; Flahou, B.; Ducatelle, R.; Smet, A.; De Bruyne, E.; Cnockaert, M.; Taminiau, B.; Daube, G.; Vandamme, P.; Haesebrouck, F.; et al. Detection, isolation and characterization of Fusobacterium gastrosuis sp. nov. colonizing the stomach of pigs. Syst. Appl. Microbiol. 2017, 40, 42–50. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Piglet | Yellow | Black | Brown |
|---|---|---|---|
| IUGR | 5 | 4 | 11 |
| Normal | 11 | 4 | 5 |
| Total number | 16 | 8 | 16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiese, M.; Hui, Y.; Nielsen, D.S.; Williams, A.R.; Lynegaard, J.C.; R. Weber, N.; Amdi, C. Color of Colon Content of Normal and Intrauterine Growth-Restricted Weaned Piglets Is Associated with Specific Microbial Taxa and Physiological Parameters. Animals 2020, 10, 1073. https://doi.org/10.3390/ani10061073
Wiese M, Hui Y, Nielsen DS, Williams AR, Lynegaard JC, R. Weber N, Amdi C. Color of Colon Content of Normal and Intrauterine Growth-Restricted Weaned Piglets Is Associated with Specific Microbial Taxa and Physiological Parameters. Animals. 2020; 10(6):1073. https://doi.org/10.3390/ani10061073
Chicago/Turabian StyleWiese, Maria, Yan Hui, Dennis S. Nielsen, Andrew R. Williams, Julie C. Lynegaard, Nicolai R. Weber, and Charlotte Amdi. 2020. "Color of Colon Content of Normal and Intrauterine Growth-Restricted Weaned Piglets Is Associated with Specific Microbial Taxa and Physiological Parameters" Animals 10, no. 6: 1073. https://doi.org/10.3390/ani10061073
APA StyleWiese, M., Hui, Y., Nielsen, D. S., Williams, A. R., Lynegaard, J. C., R. Weber, N., & Amdi, C. (2020). Color of Colon Content of Normal and Intrauterine Growth-Restricted Weaned Piglets Is Associated with Specific Microbial Taxa and Physiological Parameters. Animals, 10(6), 1073. https://doi.org/10.3390/ani10061073