
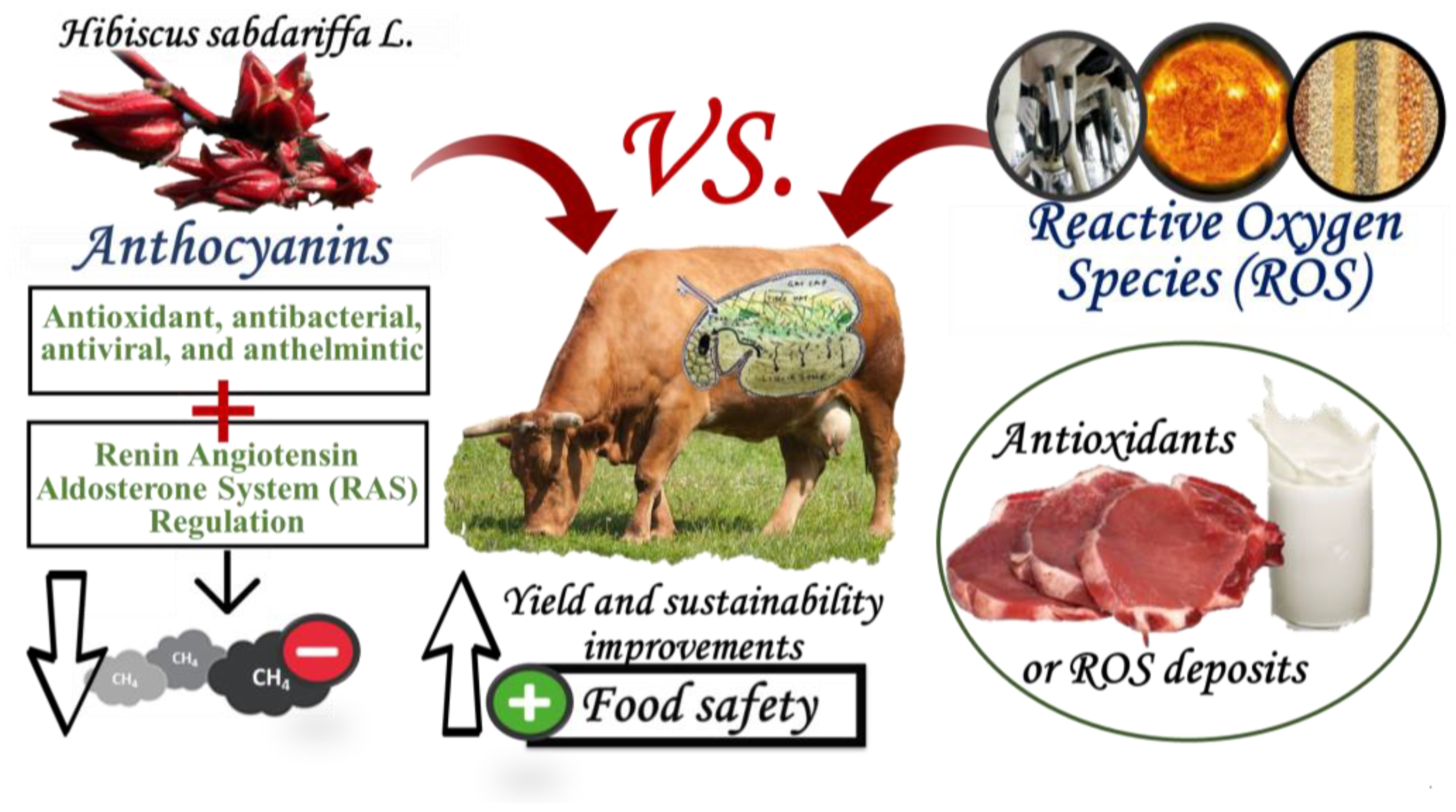
Potential Effects of Delphinidin-3-O-Sambubioside and Cyanidin-3-O-Sambubioside of Hibiscus sabdariffa L. on Ruminant Meat and Milk Quality

 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Hibiscus sabdariffa L. By-Products
3. Oxidative Stress in Ruminants
4. Potential Clinic Effects of Antioxidants
- (1)
- Promote the cytotoxicity to increase the antitumoral activities by producing H2O2 with its pyrogallol moiety or the reduction of Fe(III) to Fe(II), generating -OH ROS (although cysteine N-acetyl protect cells from cytotoxicity of H2O2, it does not avoid cell death process).
- (2)
- Promote apoptosis through mitochondrial damage, membrane depolarization, and cytochrome c release, and protects against mitochondrial damage-related cell death without changes in superoxide dismutase (SOD), glutathione peroxidase, Nrf2, Bcl2, and oxidative stress. Modulates gene expression by inhibiting various transcription factors (including Sp1, NF-κB, AP-1, STAT1, STAT3, and FOXO1) and the expression of NF-κB and AP-1. EGCG inhibits STAT1 to mediate protective effects on myocardial injury.
- (3)
- Increase second messengers, such as Ca2+, cAMP, and cGMP. EGCG elevates cytosolic Ca2+ without electrical stimulation by inhibition of sarcoplasmic/endoplasmic reticulum Ca2+-ATPase activity (SERCA), which affects the activities of Ca2+-requiring enzymes, such as calmodulin (CAM)-dependent protein kinase II and CAMKKβ (CAMKKβ is an upstream regulator of AMP-dependent kinase (AMPK), which plays crucial roles in energy metabolism and cardiovascular functions). If it stimulates vasorelaxation by increasing cAMP and cGMP in the aorta, then it may stimulate the production of cyclic nucleotides with beneficial biological effects in cardiovascular physiology.
- (4)
- Inhibit the transcription of FOXO1 to lead to the suppression of basal levels of endothelin-1 and differentiation of adipocytes. In mitochondria, EGCG enhances fat utilization, reducing the expression of leptin and stearyl-CoA desaturase while increasing fat oxidation. Moreover, EGCG regulates activities of cell surface growth factor receptors, especially receptor tyrosine kinases (RTK), including epidermal growth factor receptor (EGFR), vascular endothelial growth factor receptor (VEGFR), insulin-like growth factor receptor (IGFR), and the insulin receptor (InsR).
- (5)
- Inhibit DNA methyltransferase, which reverses methylation-induced gene silencing.
- (6)
- Inhibit autophagy, leading to apoptosis in macrophage cell lines.
5. Effect of Anthocyanins on Diet Nutritive Value and Productive Behavior in Ruminants
5.1. Effects on Ruminal Digestibility, Volatile Fatty Acids, and Methane Emission
5.2. Post-Ruminal Effects of Anthocyanins
6. Antioxidants Effect on Milk and Meat Quality as well as Shelf-Life
6.1. Anthocyanins and Polyphenols in Milk and Meat Fatty Acids Composition
6.2. Anthocyanins and Polyphenols in Milk and Meat Shelf-Life
7. Limitations and Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sáyago, A.S.G.; Arranz, S.; Serrano, J.; Goñi, I. Dietary fiber content and associated antioxidant compounds in Roselle flower (Hibiscus sabdariffa L.) beverage. J. Agric. Food Chem. 2007, 55, 7886–7890. [Google Scholar] [CrossRef] [PubMed]
- Fagbenhro, O.A. Soybean meal substitution with Roselle (Hibiscus sabdariffa L.) seed meal in dry practical diets for fingerlings of the African Catfish, Clarias gariepinus (Burchell 1822). J. Anim. Vet. Adv. 2005, 4, 473–477. [Google Scholar]
- SAGARPA—Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Plan Rector Nacional Sistema Producto Jamaica. Comité Nacional Sistema Producto Jamaica. 2012. Available online: https://docplayer.es/8913342-Plan.-rector-del-sistema-producto-jamaica.html. (accessed on 30 January 2021).
- Ariza, F.R.; Serrano, A.V.; Navarro, G.S.; Ovando, C.M.; Vázquez, G.E.; Barrios, A.A.; Michel-Aceves, A.C.; Guzman-Maldonado, S.H.; Otero-Sanchez, M.A. Variedades mexicanas de jamaica (Hibiscus sabdariffa L.) ‘alma blanca’ y ‘rosalíz’ de color claro, y ‘cotzaltzin’ y ‘tecoanapa’ de color rojo. Rev. Fitotec. Mex. 2014, 37, 181–185. [Google Scholar]
- Aragón, G.A.; Torija, T.; Avelleira, C.R.; Tapia, R.; Contreras, M.I.R.; López, O.J.F. Control de plagas de la jamaica (Hibiscus sabdariffa L.) con Gliricidia sepium (Jacq.) en Chiautla de Tapia, Puebla. Rev. AIA 2008, 12, 33–42. [Google Scholar]
- SIAP—Servicio de Información Agroalimentaria y Pesquera. Producción Agrícola. 2018. Available online: http://nube.siap.gob.mx/cierre_agricola/ (accessed on 30 January 2021).
- FAO. Food and Agriculture Organization of the United Nations. Hibiscus: Post-harvest Operations. INPhO-Post-harvet Compendium. Roma, Italia. 2004. Available online: http://www.fao.org/fileadmin/user_upload/inpho/docs/Post_Harvest_Compendium_-_Hibiscus.pdf (accessed on 30 January 2021).
- Cid, O.S.; Guerrero, B.J.A. Propiedades funcionales de la Jamaica (Hibiscus sabdariffa L.). Rev. TSIA 2012, 6, 47–63. [Google Scholar]
- Da-Costa, R.I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L. A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustan, A.C.; Devron, C. Fatty acids: Structures and properties. In Encyclopedia of Life Sciences; John Wiley and Sons: New York, NY, USA, 2005; pp. 1–7. [Google Scholar]
- Jabeur, I.; Pereira, E.; Barros, L.; Calhelha, R.C.; Sokovic, M.; Oliveira, M.B.; Ferreira, I.C.F.R. Hibiscus sabdariffa L. as a source of nutrients, bioactive compounds and colouring agents. Food Res. Int. 2017, 100, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formagio, A.S.N.; Ramos, D.D.; Vieira, M.C.; Ramalho, S.R.; Silva, M.M.; Zárate, N.A.H.; Foglio, M.A.; Calvalho, J.E. Phenolic compounds of Hibiscus sabdariffa and influence of organic residues on its antioxidant and antitumoral properties. Braz. J. Biol. 2015, 75, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Ali, B.H.; Wabel, N.A.; Bluden, G. Phytochemical, pharmacological and toxicological aspects of Hibiscus sabdariffa L.: A review. Phytother Res. 2005, 19, 369–375. [Google Scholar] [CrossRef]
- Bergmeier, D.; Berres, P.H.D.; Filippi, D.; Bilibio, D.; Bettiol, V.R.; Priamo, W.L. Extraction of total polyphenols from hibiscus (Hibiscus sabdariffa L.) and waxweed/“sete-sangrias” (Cuphea carthagenensis) and evaluation of their antioxidant potential. Acta Sci. Technol. 2014, 36, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Borrás-Linares, I.; Fernández-Arroyo, S.; Arráez-Roman, D.; Palmeros-Suárez, P.A.; del Val-Díaz, R.; Andrade-Gonzáles, I.; Fernández-Gutiérrez, A.; Gómez-Leyva, J.F.; Segura-Carretero, A. Characterization of phenolic compounds, anthocyanidin, antioxidant and antimicrobial activity of 25 varieties of Mexican Rosselle (Hibiscus sabdariffa). Ind. Crop. Prod. 2015, 69, 385–394. [Google Scholar] [CrossRef]
- Pour, P.M.; Fakhri, S.; Asgary, S.; Farzaei, M.H.; Echeverría, J. The signaling pathways, and therapeutical targets of antiviral agents: Focusing on the antiviral approaches and clinical perspectives of anthocyanin in the management of viral diseases. Front. Pharmacol. 2019, 10, 1207. [Google Scholar] [CrossRef] [Green Version]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsable for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Guardiola, S.; Mach, N. Potencial terapéutico del Hibiscus sabdariffa: Una revisión de las evidencias científicas. Endocrinol. Nutr. 2014, 61, 274–295. [Google Scholar] [CrossRef]
- Hygreeva, D.; Pandey, M.C.; Radhakrishna, K. Potential applications of plant-based derivatives as fat replacers, antioxidants and antimicrobials in fresh and processed meat products. Meat Sci. 2014, 98, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Agüero, M.; Segura, C.; Parra, J. Análisis comparativo de compuestos fenólicos totales y actividad antioxidante de cuatro marcas de tisanas de Hibiscus sabdariffa (Malvaceae) comercializadas en Costa Rica. Uniciencia 2014, 28, 34–42. [Google Scholar]
- Satúe-Gracia, M.T.; Heinonen, M.; Frankel, E.N. Anthocyanins as Antioxidants on Human Low-Density Lipoprotein and Lecithin−Liposome Systems. J. Agric. Food Chem. 1997, 45, 3362–3367. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant Activity of Plant Extracts Containing Phenolic Compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Flores, L.A.; Salinas-Moreno, Y.; Espinoza-García, B.M.; Sánchez-Feria, C. Caracterización fisicoquímica y actividad antioxidante de extractos de jamaica (Hibiscus Sabdariffa L.) Nacional e importada. Rev. Chapingo Ser. Hortic. 2008, 14, 121–129. [Google Scholar]
- Sindi, H.A.; Marshall, L.J.; Morgan, M.R.A. Comparative chemical andbiochemical analysis of extracts of Hibiscus sabdariffa. Food Chem. 2014, 164, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.; Deng, J.; Man, Y.; Qu, Y. Green Tea Extracts Epigallocatechin-3-gallate for Different Treatments. Biomed. Res. Int. 2017, 5615647. [Google Scholar] [CrossRef] [Green Version]
- Corredeu, F.; Lunesu, M.L.; Buffa, G.; Atzori, A.S.; Nudda, A.; Battacone, G.; Pulina, G. Can agro-industrial by-products rich in polyphenols be advantageously used in feeding and nutrition of dairy small ruminants? Animals 2020, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Prommachart, R.; Cherdthong, A.; Navanokraw, C.; Pongdontri, P.; Taron, W.; Uriyapongson, J.; Uriyapongson, S. Effect of dietary anthocyanin-extracted residue on meat oxidation and fatty acid profile of male dairy cattle. Animals 2021, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.S.; Celi, P.; Ponnampalam, E.N.; Leury, B.J.; Lui, F.; Dunshea, F.R. Antioxidant dynamics in the live animal and implications for ruminant health and product (milk/meat) quality: Role of vitamin E and selenium. Anim. Prod. Sci. 2014, 54, 1525–1536. [Google Scholar] [CrossRef]
- Cuhna, L.C.M.; Monteiro, M.L.; Lorenzo, J.M.; Munekata, P.E.; Muchenje, V.; Carvalho, F.A.; Conte-Junior, C.A. Natural antioxidants in processing and storage stability of sheep and goat meat products. Food Res. Int. 2018, 111, 379–390. [Google Scholar]
- Lorenzo, J.M.; Pateiro, M.; Domínguez, R.; Barba, F.J.; Putnik, P.; Bursac, K.D.; Shpigelman, A.; Granato, D.; Franco, D. Berries extracts as natural antioxidants in meat products: A review. Food Res. Int. 2018, 106, 1095–1104. [Google Scholar] [CrossRef]
- Hosoda, K.; Sasahara, H.; Matsushita, K.; Tamura, Y.; Miyaji, M.; Matsuyama, H. Anthocyanin and proanthocyanidin contents, antioxidant activity, and in situ degradability of black and red rice grains. Asian-Aust. J. Anim. Sci. 2018, 31, 1213–1220. [Google Scholar] [CrossRef]
- Tian, X.; Paengkoum, P.; Paengkoum, S.; Thongpea, S.; Ban, C. Comparison of forage yield, silage fermentative quality, anthocyanin stability, antioxidant activity, and in vitro rumen fermentation of anthocyanin-rich purple corn (Zea mays L.) stover and sticky corn stover. J. Integr. Agric. 2018, 17, 2082–2095. [Google Scholar] [CrossRef] [Green Version]
- Ianni, A.; Martino, G. Dietary grape pomace supplementation in dairy cows: Effect on nutritional quality of milk and its derived dairy products. Foods 2020, 9, 168. [Google Scholar] [CrossRef] [Green Version]
- Theodorou, M.K.; Kingston-Smith, A.H.; Winters, A.L.; Lee, M.R.F.; Minchin, F.R.; Morris, P.; MacRae, J. Polyphenols and their influence on gut function and health in ruminants: A review. Environ. Chem. Lett. 2006, 4, 121–126. [Google Scholar] [CrossRef]
- Gladine, C.; Rock, E.; Morand, C.; Bauchart, D.; Durand, D. Bioavailability and antioxidant capacity of plant extracts rich in polyphenols, given as a single acute dose, in sheep made highly susceptible to lipoperoxidation. Br. J. Nutr. 2007, 98, 691–701. [Google Scholar] [CrossRef] [Green Version]
- Olagaray, K.E.; Bradford, B.J. Plant flavonoids to improve productivity of ruminants—A review. Anim. Feed Sci. Technol. 2019, 251, 21–36. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Zebeli, Q.; Saleem, F.; Psychogios, N.; Lewis, M.J.; Dunn, S.M.; Xia, J.; Wishart, D.S. Metabolomics reveals unhealthy alterations in rumen metabolism with increased proportion of cereal grain in the diet of dairy cows. Metabolomics 2010, 6, 583–594. [Google Scholar] [CrossRef]
- Celi, P.; Gabal, G. Oxidant/antioxidant balance in animal nutrition and health: The role of protein oxidation. Front. Vet. Sci. 2015, 2, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, M.; Li, Z.; Soladoye, O.P.; van-Hecke, T. Health risks of food oxidation. Adv. Food Nutr Res. 2017, 82, 45–81. [Google Scholar]
- Huang, H.; Szumacher-Strabel, M.; Kumar, P.A.; Slusarcyk, S.; Lechniak, D.; Vazirigohar, M.; Varadyova, Z.; Kozlowska, M.; Cieslak, A. Chemical and phytcochemical composition, in vitro ruminal fermentation, methane production, and nutrient degradability of fresh and ensiled Paulownia hybrid leaves. Anim. Feed Sci. Technol. 2021, 279, 115038. [Google Scholar] [CrossRef]
- Khonkhaeng, B.; Cherdthong, A.; Chantaprasarn, N.; Harvatine, K.J.; Foiklang, S.; Chanjula, P.; Wanapat, M.; So, S.; Polyorach, S. Comparative effect of Volvariella volvacea-trated rice straw and purple corn stover fed at different levels on predicted methane production and milk fatty acid profiles in tropical dairy cows. Livestock Sci. 2021, 251, 104626. [Google Scholar] [CrossRef]
- Nikmaram, N.; Budaraju, S.; Barba, F.J.; Lorenzo, J.M.; Cox, R.B.; Mallikarjunan, K.; Roohinejad, S. Application of plant extracts to improve the shelf-life, nutritional and health-related properties of ready-to-eat meat products. Meat Sci. 2018, 145, 245–255. [Google Scholar] [CrossRef]
- Maffo, T.; Agbor, E.E.; Mekoudjou, N.H.; Kengne, S.C.N.; Gouado, I. Proximate and mineral composition, protein quality of Hibiscus sabdariffa L. (Roselle) seeds cultivated in two agroecological areas in Cameroon. Int. J. Nutr. Food Sci. 2014, 3, 251–258. [Google Scholar]
- Bryszak, M.; Szumacher-Strabel, M.; El-Sherbiny, M.; Stochmal, A.; Oleszek, W.; Roj, E.; Kumar, O.A.; Cieslak, A. Effects of berry seed residues on ruminal fermentation, methane concentration, milk production, and fatty acid proportions in the rumen and milk of dairy cows. J. Dairy Sci. 2019, 102, 1257–1273. [Google Scholar] [CrossRef] [Green Version]
- Moate, P.J.; Jacobs, J.L.; Hixson, L.J.; Deighton, M.H.; Hanna, M.C.; Morris, G.L.; Ribaux, B.E.; Wales, W.J.; Williams, S.R.O. Effects of feeding either red or white grape marc on milk production and methane emissions from early-lactation dairy cows. Animals 2020, 10, 976. [Google Scholar] [CrossRef]
- Mohd-Esa, N.; Shin, H.F.; Ismail, A.; Lye, Y.C. Antioxidant activity in different parts of roselle (Hibiscus sabdariffa L.) extracts a potential exploration of the seeds. Food Chem. 2010, 122, 1055–1060. [Google Scholar] [CrossRef]
- Sulliman, G.M.; Babiker, S.A.; Eichinger, H.M. Effect of Hibiscus seed-based diet on chemical composition, carcass characteristics and meat quality traits of cattle. Indian J. Anim. Res. 2017, 51, 694–699. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.L.; Morris, B.; Tonnis, B.; Davis, J.; Pederson, G.A. Assessment of oil Content and fatty acid composition variability in two economically important Hibiscus species. J. Agric. Food Chem. 2012, 60, 6620–6626. [Google Scholar] [CrossRef]
- Ismail, A.; Khairul, I.E.; Mohd, N.H. Roselle (Hibiscus sabdariffa L.) seeds-nutritional composition, protein quality and health benefits. Glob. Sci. Book Food 2008, 2, 1–16. [Google Scholar]
- Shaheen, M.A.; El-Nakhlawy, F.S.; Al-Shareef, A.R. Roselle (Hibiscus sabdariffa L.) seeds as unconventional nutritional source. Afr. J. Biotech. 2012, 11, 9821–9824. [Google Scholar]
- Udayasekhara, R.P. Nutrient composition and biological evaluation of mesta (Hibbiscus sabdariffa) seeds. Plant Foods Hum. Nutr. 1996, 49, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Beshir, A.A.; Babiker, S.A. The effect of feeding diet with graded levels of roselle (Hibiscus sabdariffa) seed on carcass characteristics and meat quality of Sudan desert lamb. Res. J. Animal Vet. Sci. 2009, 4, 35–44. [Google Scholar]
- Jínez, M.T.; Cortés, C.A.; Ávila, G.E.; Casaubon, M.A.; Salcedo, E.R. Efecto de niveles elevados de semilla de Jamaica (Hibiscus sabdariffa) en dietas para pollos sobre el comportamiento productivo y funcionamiento hepático. Vet. Mex. 1998, 29, 35–40. [Google Scholar]
- Kwari, I.D.; Igwebuike, J.U.; Mohammed, I.D.; Diarra, S.S. Growth, haematology and serum chemistry of broiler chickens fed raw or differently processed sorrel (Hibiscus sabdariffa) seed meal in a semi-arid environment. IJSN 2011, 2, 22–27. [Google Scholar]
- Mukhtar, A.M. The Effect of Feeding Rosella (Hibiscus sabdariffa) Seed on Broiler Chick’s Performance. Res. J. Anim. Vet. Sci. 2007, 2, 21–23. [Google Scholar]
- Soriano, T.J.; Tejada, H.I. Estudio preliminar del valor nutritivo de la semilla de Jamaica (Hibiscus sabdariffa) para el pollo de engorda. Téc. Pecu. Mex. 1995, 33, 48–52. [Google Scholar]
- Anhwange, B.A.; Ajibola, V.O.; Okibe, F.G. Nutritive value and anti-nutritional factors in Hibiscus sabdariffa. J. Fish. Int. 2006, 1, 73–76. [Google Scholar]
- Tounkara, F.; Amadou, I.; Le, G.; Shi, Y. Effect of boiling on the physicochemical properties of Roselle seeds (Hibiscus sabdariffa L.) cultivated in Mali. Afr. J. Biotech. 2011, 10, 18160–18166. [Google Scholar]
- Bellaloui, N.; Bruns, H.A.; Abbas, H.K.; Mengisty, A.; Fisher, D.K.; Reddy, K.N. Agricutural practices altered soybean protein, oil, fatty acids, sugars, and minerals in the Midsouth USA. Front. Plant. Sci. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Mahmoud, A.A.; Selim, K.A.; Abdel-Baki, M.R. Physico-chemical and oxidative stability characteristics of roselle (Hibiscus sabdariffa L.) seed oil as by-product. Egypt J. Appl. Sci. 2008, 23, 247–257. [Google Scholar]
- Rahman, A.A.; Salih, A.M.; Turki, I.Y.; Abdulatee, S.O.; Elbasheir, O.M.; Magboul, M.M. Determination of dry matter and crude protein degradability of Roselle seed (Hibiscus sabdariffa) using bags nylon technique. Sudan J. Sci. Tech. 2016, 17, 58–65. [Google Scholar]
- Daley, C.A.; Abbot, A.; Doyle, P.; Nader, G.A.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J. 2010, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Tan, D.; Mi, Y.; Zhou, Q.; Ji, S. Epigallocatechin-3-gallet attenuates cerebral cortex damage and promotes brain regeneration in acrylamide-treated rats. Food Funct. 2017, 8, 2275–2282. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, B.; Du, F.; Su, X.; Sun, G.; Zhou, G.; Bian, X.; Liu, N. Epigallocatechin-3-gallate attenuates ureteral obstruction-induced renal interstitial fibrosis in mice. J. Histochem. Cytochem. 2015, 63, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Palabiyik, S.S.; Dincer, B.; Cadirici, E.; Cinar, E.; Gundogdu, C.; Polat, B.; Yayla, M.; Halici, Z. A new update for radiocontrast-induced nephropathy aggravated with glycerol in rats: The protective potential of epigallocatechin-3-gallate. Ren. Fail. 2017, 39, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.L.; Xu, Y. Epigallocateichin gallate attenuates uric acid-induced injury rat renal interstitial fibroblasts NRK-49F by up-regulation of miR-9. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7458–7469. [Google Scholar]
- Ortíz-Romero, P.; Borralleras, C.; Campuzano, V. Epigallocatechin-3-gallate improves cardiac hypertrophy and short-term memory deficits in a Williams-Beuren syndrome mouse model. PLoS ONE 2018, 13, e0194476. [Google Scholar] [CrossRef] [Green Version]
- Klantar, M. The importance of flavonoids in ruminant nutrition. Arch. Anim. Husb. Dairy Sci. 2018, 1, 1–4. [Google Scholar]
- Sandoval-Ramírez, B.A.; Catalán, U.; Fernández-Castillejo, S.; Rubio, L.; Macia, A.; Sola, R. Anthocyanin tissue bioavailability in animals: Possible implications for human health. A systematic review. J. Agric. Food Chem. 2018, 66, 11531–11543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Du, C.; Yan, Z.; Shi, Y.; Duan, H.; Ren, Y. Inhibition of necroptosis attenuates kidney inflammation and interstitial fibrosis induced by unilateral ureteral obstruction. Am. J. Nephrol. 2017, 46, 131–138. [Google Scholar] [CrossRef]
- Kakuta, Y.; Okumi, M.; Isaka, Y.; Tsutahara, K.; Abe, T.; Yazawa, K.; Ichumaru, N.; Matsumura, K.; Hyon, S.; Takahara, S.; et al. Epigallocatechin-3-gallate protects kidneys from ischemia rerperfusion injury by HO-1 upregulation and inhibition of macrophage infiltration. Transpl. Int. 2011, 24, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parichatikanond, W.; Pinthong, D.; Mangmool, S. Blockade of the renin-angiotensin system with delphinidin, cyanin, and quiercetin. Planta Med. 2012, 78, 1626–1632. [Google Scholar]
- Nakamura, K.; Shimizu, T.; Yanagita, T.; Nemoto, T.; Taniuchi, K.; Shimizu, S.; Dimitriadis, F.; Yawata, T.; Higashi, Y.; Ueba, T.; et al. Angiotensin II acting on brain AT1 receptors induces adrenaline secretion and pressor responses in the rat. Sci. Rep. 2014, 4, 7248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, L.; Castillo, J.; Quiñones, M.; Garcia-Vallvé, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of angiotensin-converting enzyme activity by flavonoids: Structure-activity relationship studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurfaradilla, S.A.; Saputri, F.C.; Harahap, Y. Effects of Hibiscus sabdariffa calyces aqueous extract on the antihypertensive potency of captopril in the two-kidney-one-clip rat hypertension model. Evid. Based Complement. Altern. Med. 2019, 2019, 9694212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nileeka, B.W.; Rupasinghe, H.P. Plant flavonoids as angiotensin converting enzyme inhibitors in regulation of hypertension. J. Funct. Food Health Dis. 2011, 1, 172–188. [Google Scholar] [CrossRef]
- Elahi, M.Y.; Rouzbehan, Y.; Rezaee, A. Effects of phenolic compounds in three oak species on in vitro gas production using inoculums of two breeds of indigenous Iranian goats. Anim. Feed Sci. Technol. 2012, 176, 26–31. [Google Scholar] [CrossRef]
- Wang, Y.M.; Wang, J.H.; Wang, C.; Chen, B.; Liu, J.X.; Cao, H.; Guo, F.C.; Vázquez-Arañón, M. Effect of different rumen-inert fatty acids supplemented with a dietary antioxidant on performance and antioxidative status of early-lactation cows. J. Dairy Sci. 2010, 93, 3738–3745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeoula, L.M.; Machado, E.; Carraro, J.; Aguiar, S.C.; Yoshimura, E.H.; Calvo, A.B.; Seiji, S.F.; Woruby, S.N. Antioxidant action in diets with ground soybeans on ruminal microbial production, digestion, and fermentation in buffaloes. Braz. J. Vet. Res. Anim. Sci. 2019, 48, e20180167. [Google Scholar] [CrossRef] [Green Version]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2016, 101, 605–628. [Google Scholar] [CrossRef]
- Castillo, C.; Hernández, J.; López-Alonso, M.; Miranda, M.; Benedito, J.L. Values of plasma lipid hydroperoxides and total antioxidant status in healthy dairy cows: Preliminary observations. Arch. Tierz. 2003, 46, 227–233. [Google Scholar] [CrossRef]
- Salinas-Rios, T.; Sánchez-Torres-Esqueda, M.T.; Díaz-Cruz, A.; Cordero-Mora, J.L.; Cárdenas-León, M.; Hernández-Bautista, J.; Nava-Cuellar, C.; Nieto-Aquino, R. Oxidative status and fertility of ewes supplemented coffee pulp during estrous synchronization and early pregnancy. Rev. Colomb. Cienc. Pecu. 2016, 29, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, K.; Eruden, B.; Matsuyama, H.; Shioya, S. Effect of anthocyanin-rich corn silage on digestibility, milk production and plasma enzyme activities in lactating dairy cows. Anim. Sci. J. 2012, 83, 453–459. [Google Scholar] [CrossRef]
- Hosoda, K.; Miyaji, M.; Matsuyama, H.; Haga, S.; Ishizaki, H.; Nonaka, K. Effect of supplementation of purple pigment from anthocyanin-rich corn (Zea mays L.) on blood antioxidant activity and oxidation resistance in sheep. Livest. Sci. 2012, 145, 266–270. [Google Scholar] [CrossRef]
- La Rosa, M.; Kechichian, T.; Olson, G.; Saade, G.; Prewit, E.B. Lactation leads to modifications in maternal Renin-Angiotensin System in later life. Obstet. Gynecol. 2020, 27, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Janegrad, P.A.; Kia, H.A.; Moghaddam, G.; Ebrahimi, M. Evaluating caffeine antioxidant properties on Ghezel ram sperm quality after freeze-thawing. Rev. Méd. Vét. 2018, 169, 233–240. [Google Scholar]
- Elamin, K.M.; Hassan, H.E.; Abdalla, H.O.; Arabi, O.H.; Tameem, E.A.A. Effect of feeding crushed roselle seed (Hibiscus sabdariffa L.) on carcass characteristics of sudan desert sheep. Asian J. Anim. Sci. 2012, 6, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Hettiarachchy, N.S.; Horax, R.; Kannan, A.; Praisoody, A.M.D.; Muhundan, A.; Mallangi, C.R. Phytochemicals, antioxidant and antimicrobial activity of Hibiscus sabdariffa, Centella asiatica, Moringa oleifera and Murraya koenigii leaves. J. Med. Plant. Res. 2011, 5, 6672–6680. [Google Scholar]
- Valenzuela, V.C.; Pérez, P.M. Actualización en el uso de antioxidantes naturales derivados de frutas y verduras para prolongar la vida útil de la carne y productos cárneos. Rev. Chil. Nutr. 2016, 43, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Karre, L.; Lopez, K.; Getty, K.J. Natural antioxidants in meat and poultry products. Meat Sci. 2013, 94, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid–protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef]
- Jiang, J.; Xiong, Y.L. Natural antioxidants as food and feed additives to promote health benefits and quality of meat products: A review. Meat Sci. 2016, 120, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Christensen, R.A.; Drackley, J.K.; LaCount, D.W.; Clark, J.H. Infusion of four long-chain fatty acids mixtures into de abomasum of lactating dairy cows. J. Dairy Sci. 1994, 77, 1052–1069. [Google Scholar] [CrossRef]
- Martínez-Borraz, A.; Moya-Camarena, S.Y.; González-Ríos, H.; Hernández, J.; Pinelli-Saavedra, A. Conjugated linoleic acid (CLA) content in milk from confined Holstein cows during summer months in Northwestern Mexico. Rev. Mex. Cienc. Pecu. 2010, 1, 221–235. [Google Scholar]
- Hernández, A.; Kholif, A.E.; Lugo-Coyote, R.; Elghandour, M.M.Y.; Cipriano, M.; Rodríguez, G.B.; Odongo, N.E.; Salem, A.Z.M. The effect of garlic oil, xylanase enzyme and yeast on biomethane and carbon dioxide production from 60-d old Holstein dairy calves fed a high concentrate diet. J. Clean. Prod. 2017, 148, 616–623. [Google Scholar] [CrossRef] [Green Version]
- Taj, K.I.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Hayat, J.M. Antioxidants properties of milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar]
- Castillo, C.; Pereira, V.; Abuelo, A.; Hernández, J. Effect of supplementation with antioxidants on the quality of bovine milk and meat production. Sci. World J. 2013, 616098. [Google Scholar] [CrossRef]
- Velasco, V.; Williams, P. Improvement meat quality through natural antioxidants. Chil. JAR 2011, 7, 313–322. [Google Scholar]
- Maggiolino, A.; Bragaglio, A.; Rufrano, D.; Claps, S.; Sepe, L.; Damiano, S.; Ciarcia, R.; Dinardo, F.R.; Hopkins, D.L.; Neglia, G.; et al. Dietary supplementation of suckling lambs with anthocyanins: Effects on growth, carcass, oxidative and meat quality traits. Anim. Feed Sci. Tecnol. 2021, 276, 114925. [Google Scholar] [CrossRef]
- Morales, R.; Ungerfeld, E.M. Use of tannins to improve fatty acids profile of meat and milk quality in ruminants: A review. Chil. J. Agric. Res. 2015, 75, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Carmo, T.J.; Peripolli, V.; Goncalves, J.B.C.; Bergmann, T.C.; Soares, F.M.C.; Restle, J.; Kindlein, L.; McManus, C. Carcass characteristics and meat evaluation of Nelore cattle subjected to different antioxidant treatments. Rev. Bras. Zootec. 2017, 46, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Dewanckele, L.; Toral, P.G.; Vlaeminck, B.; Fievez, V. Invited review: Role of rumen biohydrogenation intermediates and rumen microbes in diet-induced milk fat depression: An update. J. Dairy Sci. 2019, 103, 7655–7681. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Authors | DM | CP | EE | CF | Ashes |
|---|---|---|---|---|---|
| g/100 g | g/100 DM | ||||
| Fagbrnhro [2] | 92.6 | 39.4 | 6.1 | 17.7 | 11.4 |
| Maffo et al. [43] | 90.0 | 22.0 | 22.0 | 20.0 | 6.1 |
| Wang et al. [48] | NR | NR | 18.0 | NR | NR |
| Ismail et al. [49] | 90.0 | 33.5 | 22.1 | 18.3 | NR |
| Shaheen and El-Nakhlawy [50] * | NR | 31.4 | 23.2 | 4.29 | 5.5 |
| Udayasekhara [51] ** | 92.4 | 20.6 | 21.0 | 41.1 | 5.4 |
| Beshir and Babikier [52] | 96.6 | 30.3 | 11.1 | 5.1 | 5.6 |
| Jínez et al. [53] | 92.5 | 20.6 | 18.0 | 23.7 | 6.7 |
| Kwari et al. [54] | NR | 38.6 | NR | 13.5 | NR |
| Mukhtar [55] | 91.8 | 21.4 | 17.4 | 12.0 | 5.3 |
| Soriano y Tejeda [56] | 92.7 | 24.8 | 17.8 | 22.9 | 1.6 |
| Anhwange et al. [57] | 94.0 | 19.8 | 28.0 | 6.3 | 5.6 |
| Tounkara et al. [58] | 91.8 | 27.3 | 20.8 | NR | 4.5 |
| Seeds | Calyxes | ||
|---|---|---|---|
| Tounkara et al. [58] | Mahmoud et al. [60] | Jabeur et al. [11] | |
| Saturated fatty acids (%) | |||
| Myristic (C14:0) | 0.21 | 0.26 | 1.24 ± 0.01 |
| Palmitic (C16:0) | 19.21 | 20.52 | 27.73 ± 0.02 |
| Stearic (C18:0) | 5.13 | 5.79 | 4.46 ± 0.01 |
| Arachidonic (C20:0) | 0.67 | 1.02 ± 0.05 | |
| Polyunsaturated fatty acids (%) | |||
| Palmitoleic (C16:1) | 0.36 | 1.32 ± 0.04 | |
| Oleic (C18:1) | 36.9 | 38.46 | 9.1 ± 0.1 |
| Linoleic (C18:2) | 35.02 | 33.25 | 32.65 ± 0.07 |
| α-linoleic (C18:3) | 1.85 | 1.69 | 15.76 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazalde-Cruz, R.; Miranda-Romero, L.A.; Tirado-González, D.N.; Carrillo-Díaz, M.I.; Medina-Cuéllar, S.E.; Mendoza-Martínez, G.D.; Lara-Bueno, A.; Tirado-Estrada, G.; Salem, A.Z.M. Potential Effects of Delphinidin-3-O-Sambubioside and Cyanidin-3-O-Sambubioside of Hibiscus sabdariffa L. on Ruminant Meat and Milk Quality. Animals 2021, 11, 2827. https://doi.org/10.3390/ani11102827
Lazalde-Cruz R, Miranda-Romero LA, Tirado-González DN, Carrillo-Díaz MI, Medina-Cuéllar SE, Mendoza-Martínez GD, Lara-Bueno A, Tirado-Estrada G, Salem AZM. Potential Effects of Delphinidin-3-O-Sambubioside and Cyanidin-3-O-Sambubioside of Hibiscus sabdariffa L. on Ruminant Meat and Milk Quality. Animals. 2021; 11(10):2827. https://doi.org/10.3390/ani11102827
Chicago/Turabian StyleLazalde-Cruz, Rosalba, Luis Alberto Miranda-Romero, Deli Nazmín Tirado-González, María Isabel Carrillo-Díaz, Sergio Ernesto Medina-Cuéllar, Germán David Mendoza-Martínez, Alejandro Lara-Bueno, Gustavo Tirado-Estrada, and Abdelfattah Z. M. Salem. 2021. "Potential Effects of Delphinidin-3-O-Sambubioside and Cyanidin-3-O-Sambubioside of Hibiscus sabdariffa L. on Ruminant Meat and Milk Quality" Animals 11, no. 10: 2827. https://doi.org/10.3390/ani11102827
APA StyleLazalde-Cruz, R., Miranda-Romero, L. A., Tirado-González, D. N., Carrillo-Díaz, M. I., Medina-Cuéllar, S. E., Mendoza-Martínez, G. D., Lara-Bueno, A., Tirado-Estrada, G., & Salem, A. Z. M. (2021). Potential Effects of Delphinidin-3-O-Sambubioside and Cyanidin-3-O-Sambubioside of Hibiscus sabdariffa L. on Ruminant Meat and Milk Quality. Animals, 11(10), 2827. https://doi.org/10.3390/ani11102827