
Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
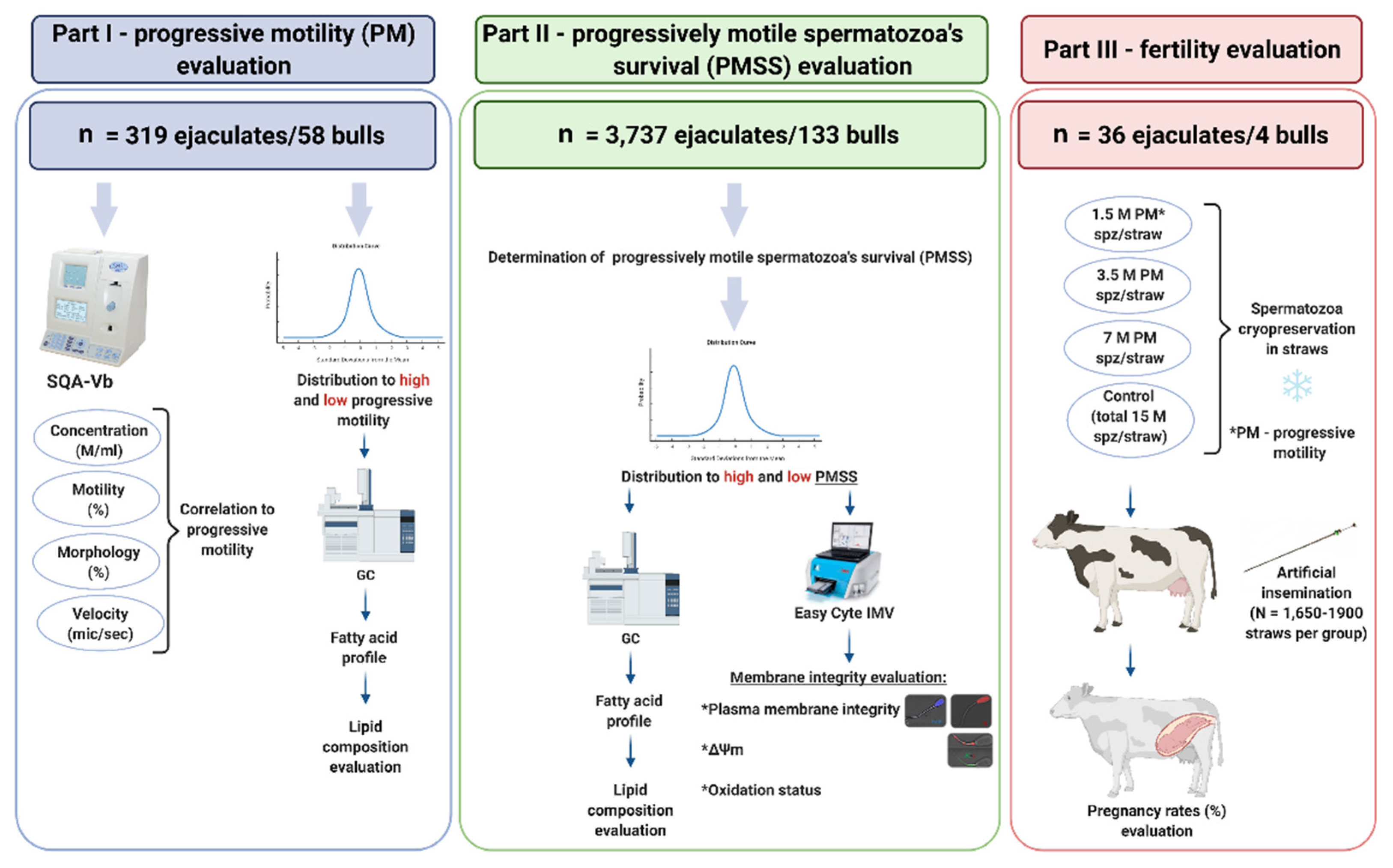
2.1. Experimental Design
2.1.1. Part I: Spermatozoa Progressive Motility
2.1.2. Part II: Progressively Motile Spermatozoa’s Survival
2.1.3. Part III: Fertility Evaluation
2.2. Semen Collection and Processing
2.3. Lipid Extraction and Fatty Acid Analysis
2.4. Flow Cytometry
2.5. Statistical Analysis
3. Results
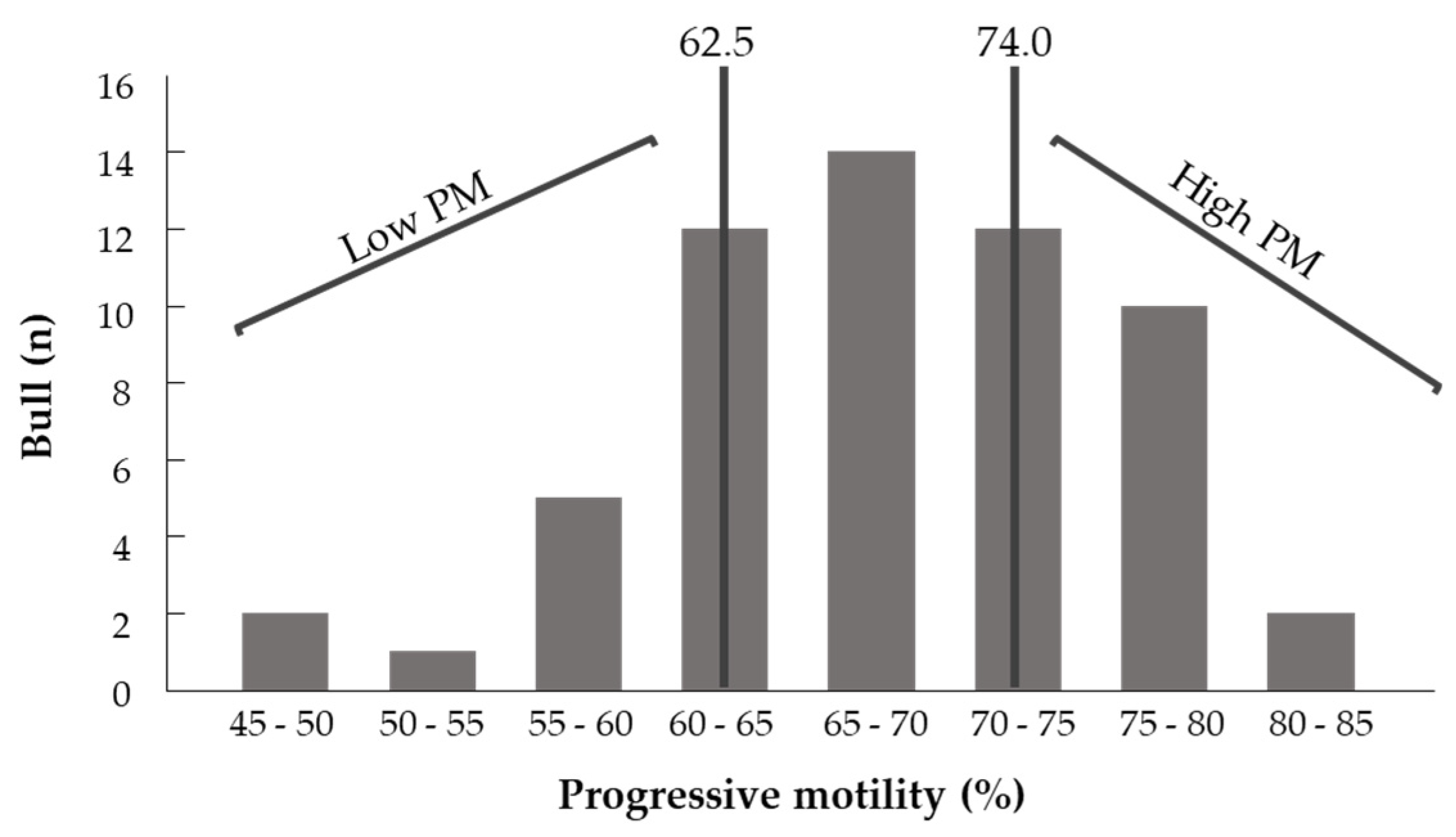
3.1. Part I: Spermatozoa Progressive Motility
3.1.1. Progressive Motility and Physiological Parameters
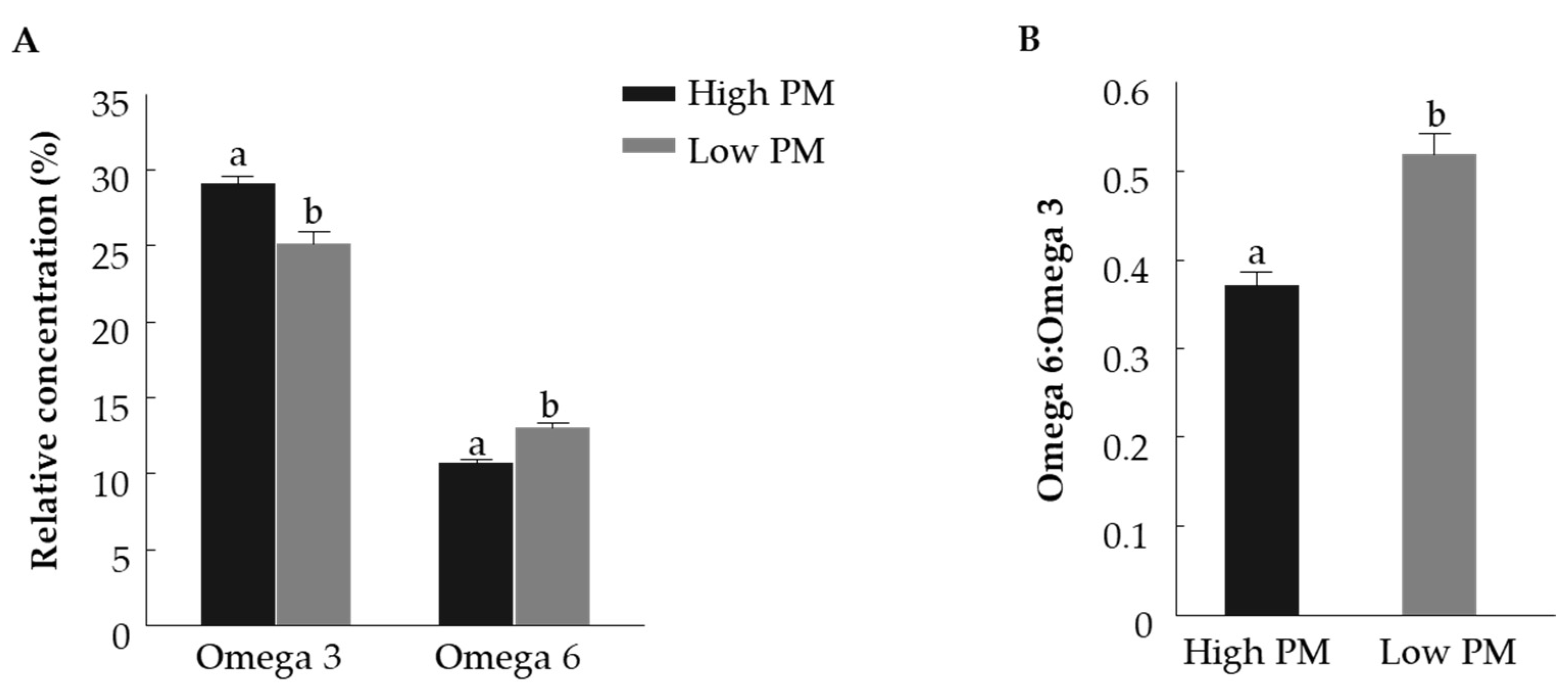
3.1.2. Fatty Acid Composition of Fresh Spermatozoa with High and Low PM
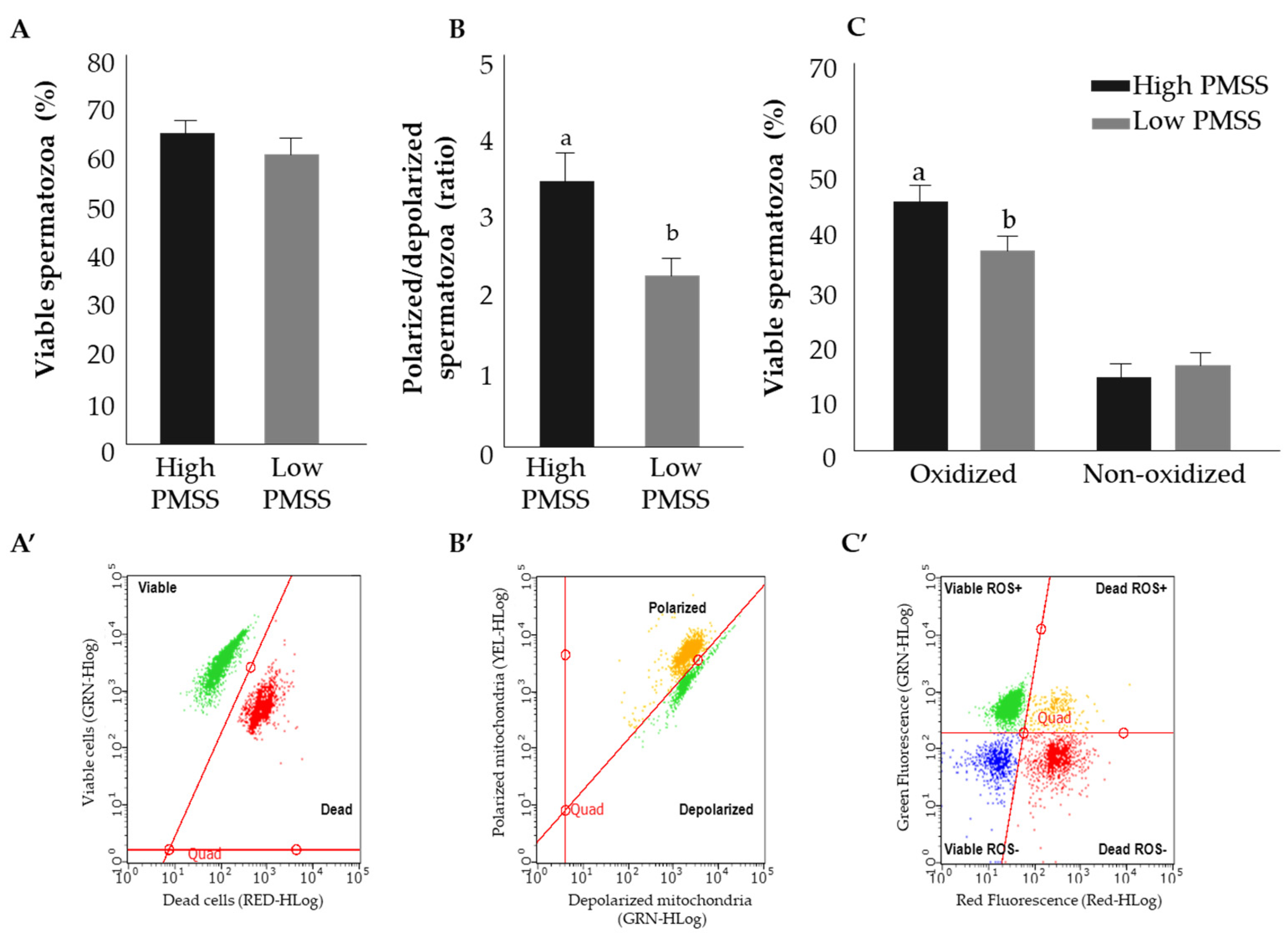
3.2. Part II: Progressively Motile Spermatozoa’s Survival
3.2.1. Progressively Motile Spermatozoa’s Survival and Physiological Parameters
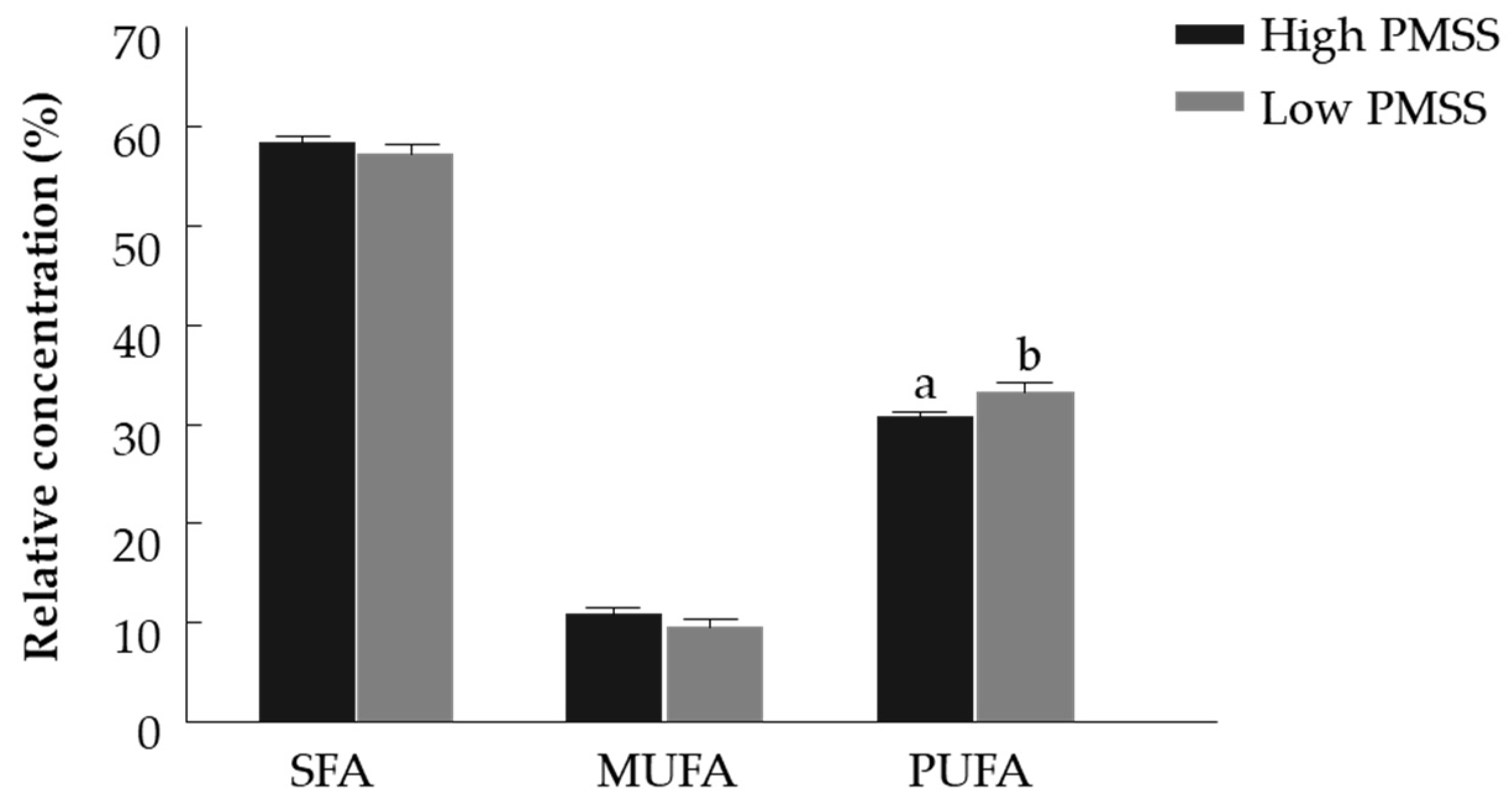
3.2.2. Fatty Acid Composition of High- and Low- PMSS for Fresh Spermatozoa
3.2.3. Structural Characteristics of Spermatozoa in the High- and Low-PMSS Groups
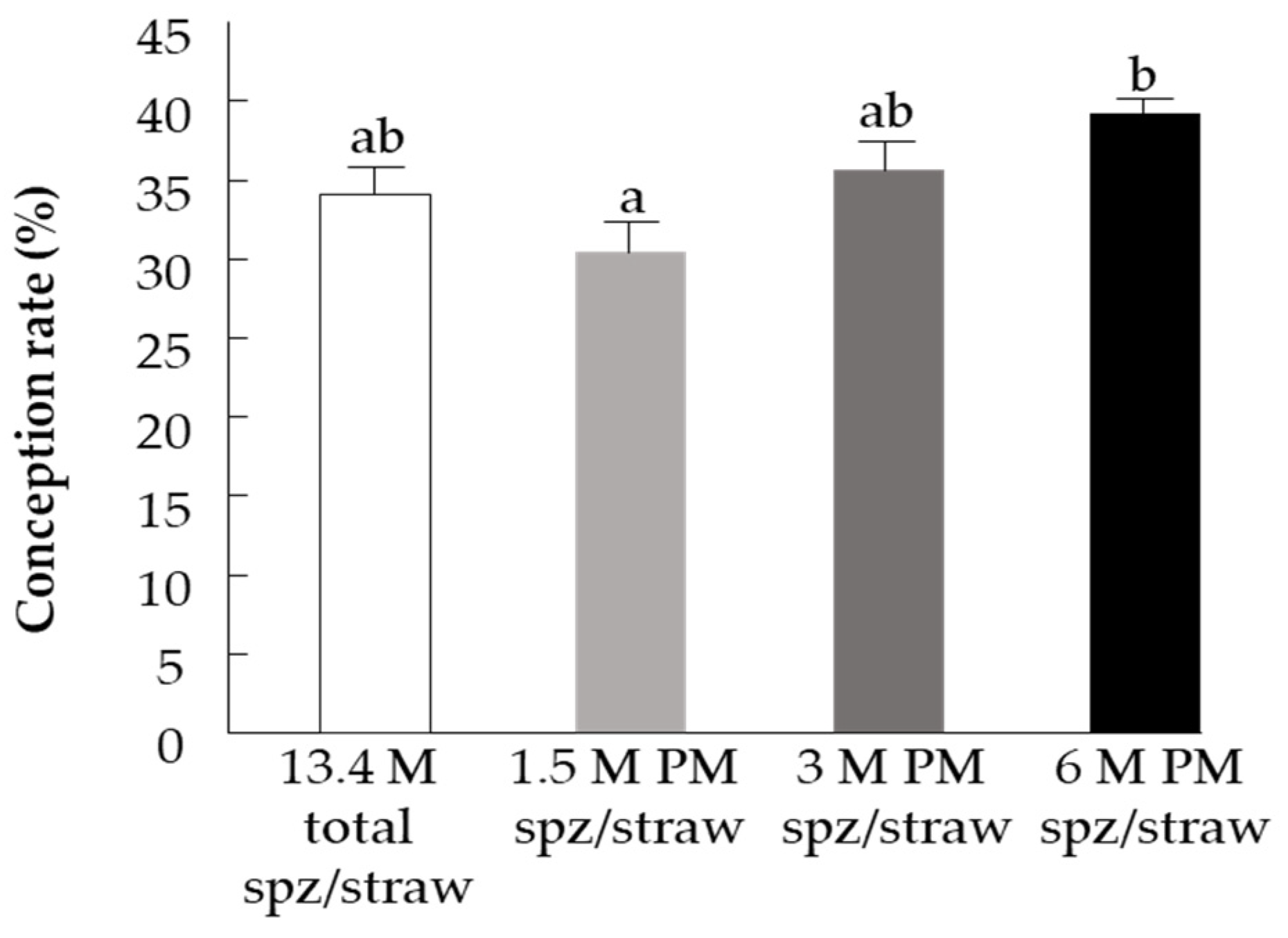
3.3. Part III: Fertility Evaluation–Spermatozoa’s PM and In-Vivo Conception Rate
4. Discussion
4.1. Progressive Motilty, Membrane Integrity and Fatty Acid Composition
4.2. Progressive Motility and Cryopreservation of Spermatozoa
4.3. Progressive Motility and Fertility Competence
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Layek, S.S.; Mohanty, T.K.; Kumaresan, A.; Parks, J.E. Cryopreservation of bull semen: Evolution from egg yolk based to soybean based extenders. Anim. Reprod. Sci. 2016, 172, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M.; Valeanu, A.S.; Lundeheim, N.; Johannisson, A. Sperm quality in frozen beef and dairy bull semen. Acta Vet. Scand. 2018, 60, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kathiravan, P.; Kalatharan, J.; Karthikeya, G.; Rengarajan, K.; Kadirvel, G. Objective Sperm Motion Analysis to Assess Dairy Bull Fertility Using Computer-Aided System—A Review. Reprod. Domest. Anim. 2011, 46, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Jedrzejczak, P.; Taszarek-Hauke, G.; Hauke, J.; Pawelczyk, L.; Duleba, A.J. Prediction of spontaneous conception based on semen parameters. Int. J. Androl. 2008, 31, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Lewis, S.E.M. Sperm DNA damage or progressive motility: Which one is the better predictor of fertilization in vitro? Syst. Biol. Reprod. Med. 2011, 57, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Brinsko, S.P.; Blanchard, T.L.; Varner, D.D.; Schumacher, J.; Love, C.C.; Hinrichs, K.; Hartman, D.L. Examination of the Stallion for Breeding Soundness. In Manual of Equine Reproduction, 3rd ed.; Elsevier: St. Louis, MO, USA, 2011; pp. 176–206. [Google Scholar] [CrossRef]
- Ahmed, H.; Andrabi, S.M.H.; Jahan, S. Semen quality parameters as fertility predictors of water buffalo bull spermatozoa during low-breeding season. Theriogenology 2016, 86, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Kathiravan, P.; Kalatharan, J.; John Edwin, M.; Veerapandian, C. Computer automated motion analysis of crossbred bull spermatozoa and its relationship with in vitro fertility in zona-free hamster oocytes. Anim. Reprod. Sci. 2008, 104, 9–17. [Google Scholar] [CrossRef]
- Li, Y.; Kalo, D.; Zeron, Y.; Roth, Z.; Smith, R.H. Progressive motility—A potential predictive parameter for semen fertilization capacity in bovines. Zygote 2016, 24, 70–82. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Hamada, A.; Wasik, M. Sperm Banking: Indications and Regulations. In Infertility: Diagnosis, management and IVF; Jaypee Brothers Medical Publishers Ltd.: New Delhi, India, 2012. [Google Scholar] [CrossRef]
- Castro, L.S.; Hamilton, T.R.S.; Mendes, C.M.; Nichi, M.; Barnabe, V.H.; Visintin, J.A.; Assumpção, M.E.O.A. Sperm cryodamage occurs after rapid freezing phase: Flow cytometry approach and antioxidant enzymes activity at different stages of cryopreservation. J. Anim. Sci. Biotechnol. 2016, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, H.; Andrabi, S.M.H.; Shah, S.A.H.; Jahan, S. Effect of Cryopreservation on Casa Characteristics, Mitochondrial Transmembrane Potential, Plasma and Acrosome Integrities, Morphology and in vivo Fertility of Buffalo Bull Spermatozoa. Cryo Lett. 2019, 40, 173–180. [Google Scholar]
- Hammerstedt, R.H.; Graham, J.K.; Nolan, J.P. Cryopreservation of Mammalian Sperm: What We Ask Them to Survive. J. Androl. 1990, 11, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.L.; Bilodeau, J.F.; Cormier, N. Minireview: Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon. J. Androl. 2000, 21, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Am-in, N.; Kirkwood, R.N.; Techakumphu, M.; Tantasuparuk, W. Lipid profiles of sperm and seminal plasma from boars having normal or low sperm motility. Theriogenology 2011, 75, 897–903. [Google Scholar] [CrossRef]
- Lucio, C.F.; Brito, M.M.; Angrimani, D.S.R.; Belaz, K.R.A.; Morais, D.; Zampieri, D.; Losano, J.D.A.; Assumpção, M.E.O.A.; Nichi, M.; Eberlin, M.N.; et al. Lipid composition of the canine sperm plasma membrane as markers of sperm motility. Reprod. Domest. Anim. 2017, 52, 208–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argov-Argaman, N.; Mahgrefthe, K.; Zeron, Y.; Roth, Z. Season-induced variation in lipid composition is associated with semen quality in Holstein bulls. Reproduction 2013, 145, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Shalit, U.; Rabinovitch, L.; Greenberg, S.; Zeron, Y. Performance evaluation of the SQA-Vb automated sperm quality analyzer for bulls. Theriogenology 2008, 70, 585–586. [Google Scholar] [CrossRef]
- Argov-Argaman, N.; Mahgrefthe, K.; Zeron, Y.; Roth, Z. Variation in lipid profiles within semen compartments-the bovine model of aging. Theriogenology 2013, 80, 712–721. [Google Scholar] [CrossRef]
- Sellem, E.; Broekhuijse, M.L.W.J.; Chevrier, L.; Camugli, S.; Schmitt, E.; Schibler, L.; Koenen, E.P.C. Use of combinations of in vitro quality assessments to predict fertility of bovine semen. Theriogenology 2015, 84, 1447–1454.e5. [Google Scholar] [CrossRef] [PubMed]
- Komsky-Elbaz, A.; Roth, Z. Fluorimetric techniques for the assessment of sperm membranes. J. Vis. Exp. 2018, 2018, e58622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane lipid composition: Effect on membrane and organelle structure, function and compartmentalization and therapeutic avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalata, A.A.; Christophe, A.B.; Depuydt, C.E.; Schoonjans, F.; Comhaire, F.H. The fatty acid composition of phospholipids of spermatozoa from infertile patients. Mol. Hum. Reprod. 1998, 4, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safarinejad, M.R.; Hosseini, S.Y.; Dadkhah, F.; Asgari, M.A. Relationship of omega-3 and omega-6 fatty acids with semen characteristics, and anti-oxidant status of seminal plasma: A comparison between fertile and infertile men. Clin. Nutr. 2010, 29, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Israelachvili, J.N.; Marcelja, S.; Horn, R.G.; Israelachvili, J.N. Physical principles of membrane organization. Q. Rev. Biophys. 1980, 13, 121–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connor, W.E.; Lin, D.S.; Wolf, D.P.; Alexander, M. Uneven distribution of desmosterol and docosahexaenoic acid in the heads and tails of monkey sperm. J. Lipid Res. 1998, 39, 1404–1411. [Google Scholar] [CrossRef]
- Nissen, H.P.; Kreysel, H.W. Polyunsaturated Fatty Acids in Relation to Sperm Motility. Andrologia 1983, 15, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Mostek, A.; Dietrich, M.A.; Słowińska, M.; Ciereszko, A. Cryopreservation of bull semen is associated with carbonylation of sperm proteins. Theriogenology 2017, 92, 95–102. [Google Scholar] [CrossRef]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Wallach, E.E.; Zamboni, L. The ultrastructural pathology of the spermatozoon as a cause of infertility: The role of electron microscopy in the evaluation of semen quality. Fertil. Steril. 1987, 48, 711–734. [Google Scholar] [CrossRef]
- O’Connell, M.; McClure, N.; Lewis, S.E.M. The effects of cryopreservation on sperm morphology, motility and mitochondrial function. Hum. Reprod. 2002, 17, 704–709. [Google Scholar] [CrossRef]
- Sharma, V. Sperm storage for cancer patients in the UK: A review of current practice. Hum. Reprod. 2011, 26, 2935–2943. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, J.F.H.M.; Gadella, B.M. In situ detection and localization of lipid peroxidation in individual bovine sperm cells. Free Radic. Biol. Med. 2003, 35, 1382–1391. [Google Scholar] [CrossRef] [PubMed]
- Celeghini, E.C.C.; de Arruda, R.P.; de Andrade, A.F.C.; Nascimento, J.; Raphael, C.F.; Rodrigues, P.H.M. Effects that bovine sperm cryopreservation using two different extenders has on sperm membranes and chromatin. Anim. Reprod. Sci. 2008, 104, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Vincent, P.; Underwood, S.L.; Dolbec, C.; Bouchard, N.; Kroetsch, T.; Blondin, P. Bovine Semen Quality Control in Artificial Insemination Centers. In Bovine Reproduction; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 685–695. [Google Scholar] [CrossRef]
- Narud, B.; Klinkenberg, G.; Khezri, A.; Zeremichael, T.T.; Stenseth, E.-B.; Nordborg, A.; Haukaas, T.H.; Morrel, J.M.; Heringstad, B.; Myromslien, F.D.; et al. Differences in sperm functionality and intracellular metabolites in Norwegian Red bulls of contrasting fertility. Theriogenology 2020, 157, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, H.; Barth, A.D. In vitro evaluation of sperm quality related to in vivo function and fertility. Soc. Reprod. Fertil. Suppl. 2007, 64, 39–54. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FA | High PM | Low PM | p-Value |
|---|---|---|---|
| C10:0 | 0.072 | 0.041 | 0.12 |
| C11:0 | 0.068 | 0.071 | 0.92 |
| C12:0 | 0.307 | 0.306 | 0.98 |
| C13:0 | 0.053 | 0.041 | 0.27 |
| C14:0 | 17.7 | 18.8 | 0.58 |
| C15:0 | 0.2 | 0.2 | 0.99 |
| C16:0 | 25.74 | 25.68 | 0.96 |
| C17:0 | 0.279 | 0.263 | 0.54 |
| C18:0 | 8.75 | 9.29 | 0.23 |
| C20:0 | 0.169 | 0.211 | 0.06 |
| C21:0 | 0.045 | 0.082 | 0.06 |
| C22:0 | 0.200 | 0.219 | 0.47 |
| C23:0 | 0.103 | 0.106 | 0.73 |
| C24:0 | 0.384 | 0.248 | 0.35 |
| C14:1 | 0.045 | 0.061 | 0.08 |
| C15:1 | 0.218 | 0.255 | 0.07 |
| C16:1n7 cis9 | 0.184 | 0.169 | 0.43 |
| C18:1n7 | 2.79 | 2.98 | 0.23 |
| C18:1n9 t | 0.162 | 0.157 | 0.87 |
| C18:1n9 cis | 2.388 | 2.322 | 0.74 |
| C18:2n6 cis9,12 | 4.28 | 5.726 | <0.001 |
| C18:3n3 cis12,15 | 0.114 | 0.131 | 0.19 |
| C18:3n6 | 0.098 | 0.992 | 0.99 |
| 20:1n9 | 0.137 | 0. 139 | 0.87 |
| C22:1 cis13 | 0.085 | 0.117 | 0.2 |
| 20:2n6 | 0.311 | 0.649 | <0.001 |
| 20:3n6 | 0.757 | 0.730 | 0.69 |
| C20:4n6 | 4.409 | 4.515 | 0.74 |
| C20:5n3 | 0.043 | 0.047 | 0.65 |
| C22:4n6 | 0.871 | 1.310 | 0.002 |
| C22:6n3 | 29.01 | 24.93 | 0.03 |
| SFA | 54.086 | 55.642 | 0.389 |
| MUFA | 6.01 | 6.21 | 0.564 |
| PUFA | 39.9 | 38.1 | 0.346 |
| Item | Fresh (Mean ± SD) | Frozen (Mean ± SD) | ∆ (pp) | |
|---|---|---|---|---|
| Motility (%) | Total | 76.00 ± 9.69 a | 51.00 ± 7.04 b | 25.00 ± 11.19 X |
| Progressive motility (%) | Total | 66.00 ± 8.13 a | 37.81 ± 7.88 b | 28.19 ± 10.74 Y |
| High (>74.0%) | 78.73 ± 3.31 a | 38.70 ± 8.32 b | 40.03 ± 8.96 * | |
| Low (<62.5%) | 57.67 ± 3.75 a | 37.05 ± 7.51 b | 20.62 ± 7.88 ** | |
| FA | High PMSS | Low PMSS | p-Value |
|---|---|---|---|
| C14:0 | 17.18 | 13.44 | 0.02 |
| C15:0 | 0.28 | 0.35 | 0.01 |
| C16:0 | 28.31 | 30.90 | 0.02 |
| C18:0 | 12.35 | 12.10 | 0.61 |
| C20:0 | 0.25 | 0.35 | 0.01 |
| C16:1n7 cis9 | 0.26 | 0.33 | 0.005 |
| C18:1n7 | 2.64 | 2.58 | 0.55 |
| C18:1n9 t | 0.41 | 0.45 | 0.34 |
| C18:1n9 cis | 7.32 | 5.93 | 0.12 |
| C18:2n6 cis9,12 | 4.03 | 4.95 | 0.001 |
| C18:3n3 cis 9,12,15 | 0.20 | 0.25 | 0.03 |
| C18:3n6 | 0.23 | 0.37 | 0.001 |
| 20:1n9 | 0.25 | 0.32 | 0.02 |
| 20:3n6 | 0.59 | 0.55 | 0.84 |
| C20:4n6 | 3.65 | 3.56 | 0.47 |
| C22:4n6 | 0.97 | 1.01 | 0.81 |
| C22:6n3 | 21.08 | 22.57 | 0.14 |
| O3 | 21.28 | 22.82 | 0.118 |
| O6 | 9.46 | 10.44 | 0.007 |
| O6/O3 | 0.45 | 0.47 | 0.335 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kogan, T.; Grossman Dahan, D.; Laor, R.; Argov-Argaman, N.; Zeron, Y.; Komsky-Elbaz, A.; Kalo, D.; Roth, Z. Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa. Animals 2021, 11, 2948. https://doi.org/10.3390/ani11102948
Kogan T, Grossman Dahan D, Laor R, Argov-Argaman N, Zeron Y, Komsky-Elbaz A, Kalo D, Roth Z. Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa. Animals. 2021; 11(10):2948. https://doi.org/10.3390/ani11102948
Chicago/Turabian StyleKogan, Tanya, Dana Grossman Dahan, Ronit Laor, Nurit Argov-Argaman, Yoel Zeron, Alisa Komsky-Elbaz, Dorit Kalo, and Zvi Roth. 2021. "Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa" Animals 11, no. 10: 2948. https://doi.org/10.3390/ani11102948
APA StyleKogan, T., Grossman Dahan, D., Laor, R., Argov-Argaman, N., Zeron, Y., Komsky-Elbaz, A., Kalo, D., & Roth, Z. (2021). Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa. Animals, 11(10), 2948. https://doi.org/10.3390/ani11102948