
Population Dynamics, Fecundity and Fatty Acid Composition of Oithona nana (Cyclopoida, Copepoda), Fed on Different Diets


 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Copepods Stock Culture
2.2. Regime and Experimental Design
2.3. Tested Parameters
2.3.1. Population Growth, Growth Rate, Composition, and Fecundity
2.3.2. Fatty acid Analysis
2.3.3. Data Analysis
3. Results
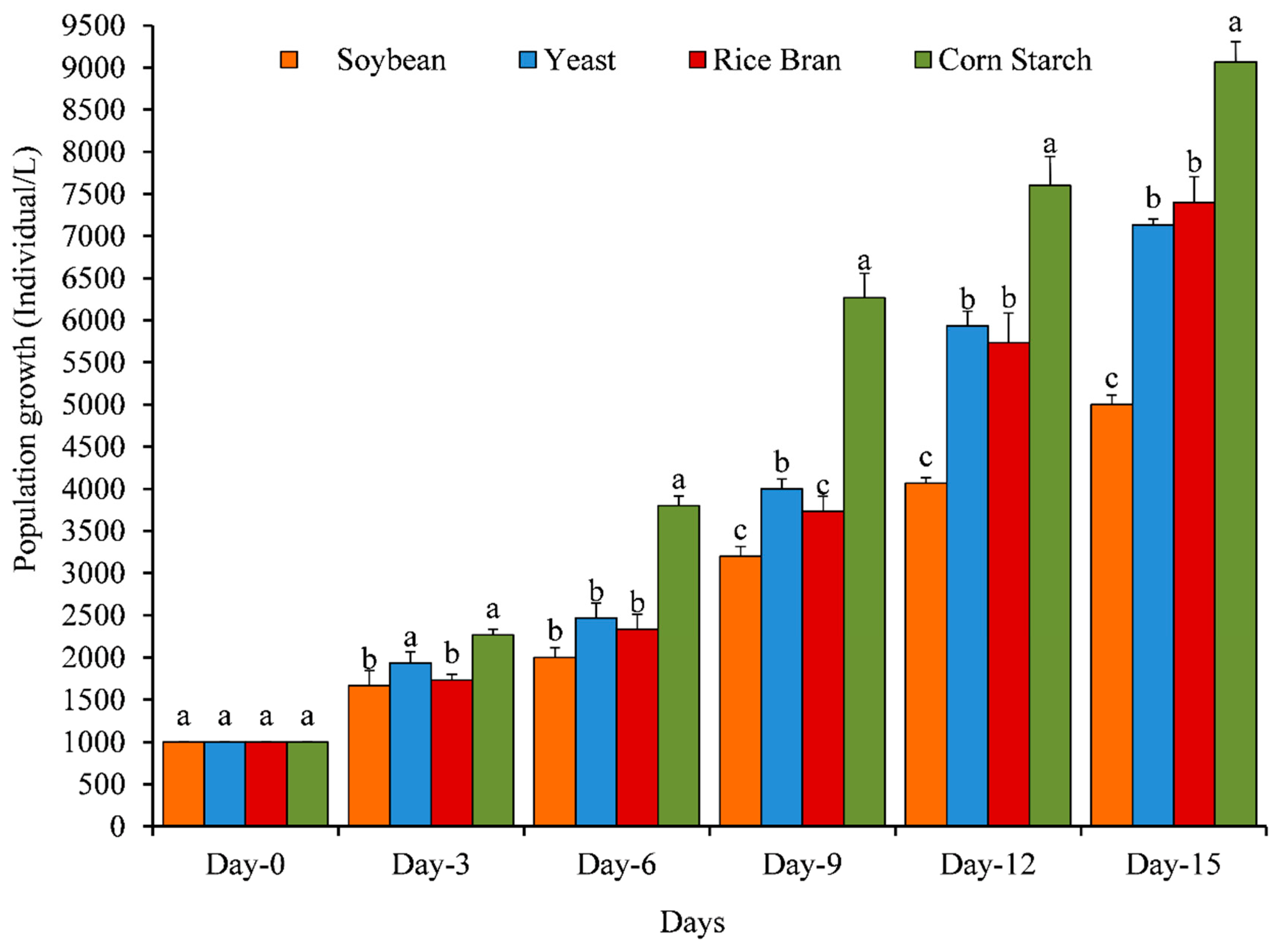
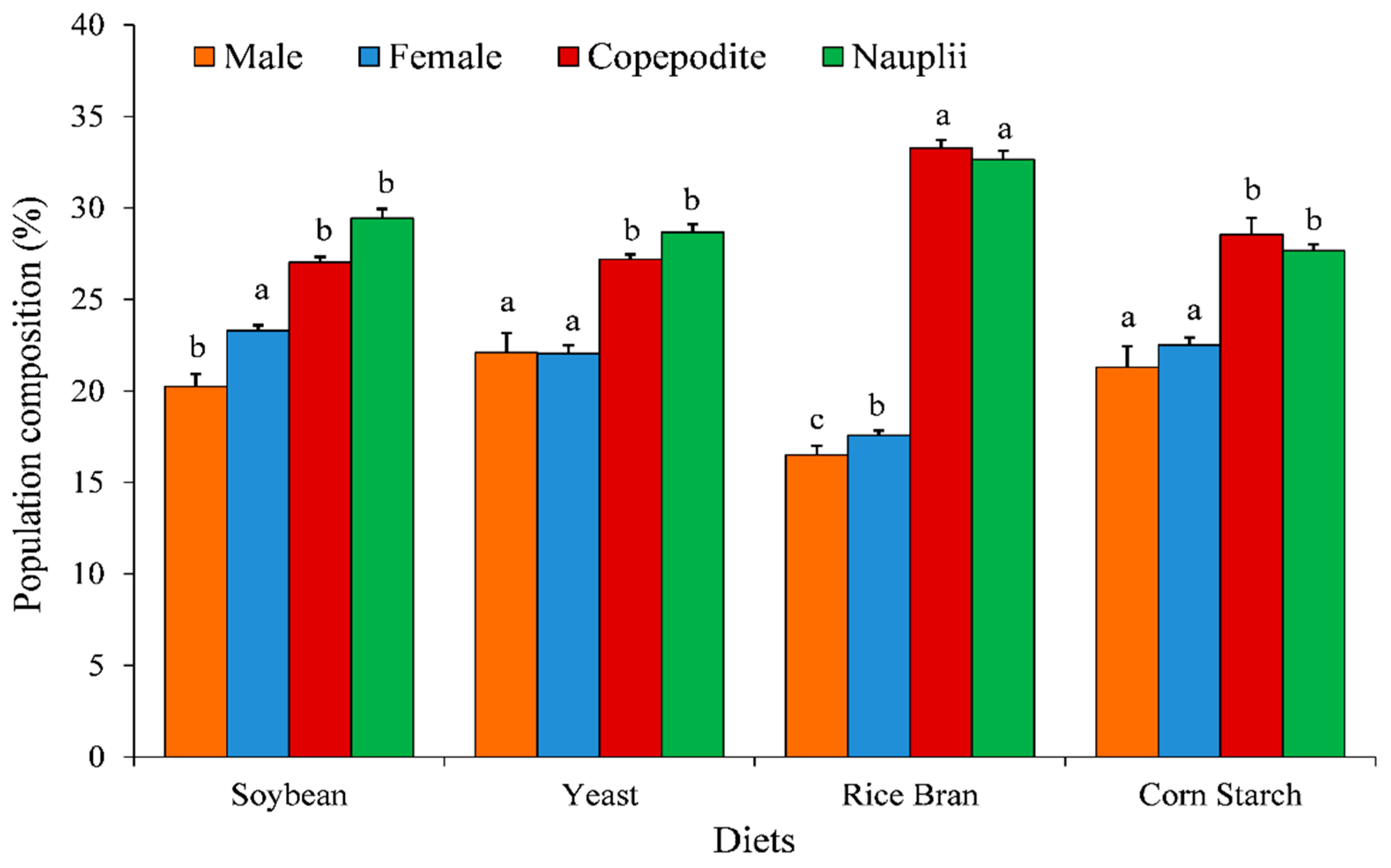
3.1. Population Growth, Growth Rate, Composition and Fecundity
3.2. Fatty Acid Compositions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khairy, H.M.; El-Sayed, H.S. Effect of enriched Brachionus plicatilis and Artemia salina nauplii by microalga Tetraselmis chuii (Bütcher) grown on four different culture media on the growth and survival of Sparus aurata larvae. Afr. J. Biotechnol. 2012, 11, 399–415. [Google Scholar]
- Abdel-Rahman, S.; Abdel Razek, F.; AbouZeid, A.; Ashour, M. Population growth rate, fecundity, filtration and ingestion rate of marine rotifer Brachionus plicatilis fed with motile and immotile microalgae. Egypt. J. Aquat. Res. 2008, 34, 426–439. [Google Scholar]
- Abdel-Rahman, H.; Abdel Razek, A.; AbouZeid, A.; Ashour, M. Optimum growth conditions of three isolated diatoms species Skeletonema costatum, Chaetoceros calcitrans and Detonula confervacea and their utilization as feed for marine penaeid shrimp larvae. Egypt. J. Aquat. Res. 2010, 36, 161–183. [Google Scholar]
- Drillet, G.; Frouël, S.; Sichlau, M.H.; Jepsen, P.M.; Højgaard, J.K.; Joarder, A.K.; Hansen, B.W. Status and recommendations on marine copepod cultivation for use as live feed. Aquaculture 2011, 315, 155–166. [Google Scholar] [CrossRef]
- Sharawy, Z.Z.; Ashour, M.; Abbas, E.; Ashry, O.; Helal, M.; Nazmi, H.; Kelany, M.; Kamel, A.; Hassaan, M.; Rossi Jr, W. Effects of dietary marine microalgae, Tetraselmis suecica, on production, gene expression, protein markers and bacterial count of Pacific white shrimp Litopenaeus vannamei. Aquac. Res. 2020, 51, 2216–2228. [Google Scholar] [CrossRef]
- Abo-Taleb, H.; Ashour, M.; El-Shafei, A.; Alataway, A.; Maaty, M.M. Biodiversity of Calanoida Copepoda in Different Habitats of the North-Western Red Sea (Hurghada Shelf). Water 2020, 12, 656. [Google Scholar] [CrossRef] [Green Version]
- El-Gamal, M.M.; Othman, S.I.; Abdel-Rahim, M.M.; Mansour, A.T.; Alsaqufi, A.S.; El Atafy, M.M.; Mona, M.H.; Allam, A.A. Palaemon and artemia supplemented diet enhances sea bass, Dicentrarchus labrax, broodstock reproductive performance and egg quality. Aquac. Rep. 2020, 16, 100290. [Google Scholar] [CrossRef]
- Heneash, A.; Ashour, M.; Matar, M. Effect of Un-live Microalgal diet, Nannochloropsis oculata and Arthrospira (Spirulina) platensis, comparing to yeast on population of rotifer, Brachionus plicatilis. Mediterr. Aquac. J. 2015, 7, 48–54. [Google Scholar] [CrossRef]
- Ashour, M.; Elshobary, M.E.; El-Shenody, R.; Kamil, A.-W.; Abomohra, A.E.-F. Evaluation of a native oleaginous marine microalga Nannochloropsis oceanica for dual use in biodiesel production and aquaculture feed. Biomass Bioenergy 2019, 120, 439–447. [Google Scholar] [CrossRef]
- Støttrup, J. The elusive copepods: Their production and suitability in marine aquaculture. Aquac. Res. 2000, 31, 703–711. [Google Scholar] [CrossRef]
- Olivotto, I.; Tokle, N.; Nozzi, V.; Cossignani, L.; Carnevali, O. Preserved copepods as a new technology for the marine ornamental fish aquaculture: A feeding study. Aquaculture 2010, 308, 124–131. [Google Scholar] [CrossRef]
- Abate, T.G.; Nielsen, R.; Nielsen, M.; Jepsen, P.M.; Hansen, B.W. A cost-effectiveness analysis of live feeds in juvenile turbot Scophthalmus maximus (Linnaeus, 1758) farming: Copepods versus A rtemia. Aquac. Nutr. 2016, 22, 899–910. [Google Scholar] [CrossRef]
- Øie, G.; Galloway, T.; Sørøy, M.; Holmvaag Hansen, M.; Norheim, I.; Halseth, C.; Almli, M.; Berg, M.; Gagnat, M.; Wold, P.A. Effect of cultivated copepods (Acartia tonsa) in first-feeding of Atlantic cod (Gadus morhua) and ballan wrasse (Labrus bergylta) larvae. Aquac. Nutr. 2017, 23, 3–17. [Google Scholar] [CrossRef]
- Abdel-Rahim, M.M.; Mansour, A.T.; Mona, M.H.; El-Gamal, M.M.; El Atafy, M.M. To what extent can maternal inherited immunity acquired from a crustacean-enhanced diet improve the performance and vitality of the offspring and enhance profitability of European Sea bass (Dicentrarchus labrax)? J. World Aquac. Soc. 2019, 50, 550–574. [Google Scholar] [CrossRef]
- Blaxter, J.H.; Douglas, B.; Tyler, P.A.; Mauchline, J. The Biology of Calanoid Copepods; Academic Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Støttrup, J. Production and nutritional value of copepods. In Live Feeds in Marine Aquaculture; Støttrup, J., McEvoy, L., Eds.; Blackwell: Oxford, UK, 2003; p. 318. [Google Scholar]
- Lee, C.; O’Bryen, P.; Marcus, N. Copepods in Aquaculture; Wiley-Blackwell: Oxford, UK, 2005. [Google Scholar]
- Ashour, M.; Abo-Taleb, H.; Abou-Mahmoud, M.; El-Feky, M. Effect of the integration between plankton natural productivity and environmental assessment of irrigation water, El-Mahmoudia Canal, on aquaculture potential of Oreochromis niloticus. Turk. J. Fish. Aquatic Sci. 2018, 18, 1163–1175. [Google Scholar]
- Abo-Taleb, H.; Zeina, A.F.; Ashour, M.; Mabrouk, M.M.; Sallam, A.E.; El-feky, M.M. Isolation and cultivation of the freshwater amphipod Gammarus pulex (Linnaeus, 1758), with an evaluation of its chemical and nutritional content. Egypt. J. Aquat. Biol. Fish. 2020, 24, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.T. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Albano, M.; Panarello, G.; Di Paola, D.; D’Angelo, G.; Granata, A.; Savoca, S.; Capillo, G. The mauve stinger Pelagia noctiluca (Cnidaria, Scyphozoa) plastics contamination, the Strait of Messina case. Int. J. Environ. Stud. 2021, 1–6. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Matsuno, K.; Homma, T. Spatial changes in the vertical distribution of calanoid copepods down to great depths in the North Pacific. Zool. Stud. 2015, 54, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kleppel, G.; Hazzard, S.; Burkart, C. Maximizing the nutritional values of copepods in aquaculture: Managed versus balanced nutrition. In Copepods in Aquaculture; Lee, C.S., O’Bryen, P.J., Marcus, N.H., Eds.; Blackwell Publishing: Oxford, UK, 2005; pp. 49–60. [Google Scholar]
- Van der Meeren, T.; Olsen, R.E.; Hamre, K.; Fyhn, H.J. Biochemical composition of copepods for evaluation of feed quality in production of juvenile marine fish. Aquaculture 2008, 274, 375–397. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.-J.; Souissi, A.; Souissi, S.; Hwang, J.-S. Effects of salinity on the reproductive performance of Apocyclops royi (Copepoda, Cyclopoida). J. Exp. Mar. Biol. Ecol. 2016, 475, 108–113. [Google Scholar] [CrossRef]
- Vidhya, K.; Uthayakumar, V.; Muthukumar, S.; Munirasu, S.; Ramasubramanian, V. The effects of mixed algal diets on population growth, egg productivity and nutritional profiles in cyclopoid copepods (Thermocyclops hyalinus and Mesocyclops aspericornis). J. Basic Appl. Zool. 2014, 67, 58–65. [Google Scholar] [CrossRef] [Green Version]
- El-khodary, G.M.; Mona, M.M.; El-sayed, H.S.; Ghoneim, A.Z. Phylogenetic identification and assessment of the nutritional value of different diets for a copepod species isolated from Eastern Harbor coastal region. Egypt. J. Aquat. Res. 2020, 46, 173–180. [Google Scholar] [CrossRef]
- Elshobary, M.E.; El-Shenody, R.A.; Ashour, M.; Zabed, H.M.; Qi, X. Antimicrobial and antioxidant characterization of bioactive components from Chlorococcum minutum. Food Biosci. 2020, 35, 100567. [Google Scholar] [CrossRef]
- Zaki, M.A.; Ashour, M.; Heneash, A.M.; Mabrouk, M.M.; Alprol, A.E.; Khairy, H.M.; Nour, A.M.; Mansour, A.T.; Hassanien, H.A.; Gaber, A. Potential Applications of native cyanobacterium isolate (Arthrospira platensis NIOF17/003) for biodiesel production and utilization of its byproduct in marine rotifer (Brachionus plicatilis) production. Sustainability 2021, 13, 1769. [Google Scholar] [CrossRef]
- Ashour, M. Current and future perspectives of microalgae-aquaculture in Egypt, case study: SIMAF-prototype-project. Egypt. J. Anim. Prod. 2020, 57, 163–170. [Google Scholar]
- Lee, K.W.; Park, H.G.; Lee, S.-M.; Kang, H.-K. Effects of diets on the growth of the brackish water cyclopoid copepod Paracyclopina nana Smirnov. Aquaculture 2006, 256, 346–353. [Google Scholar] [CrossRef]
- Payne, M.; Rippingale, R. Evaluation of diets for culture of the calanoid copepod Gladioferens imparipes. Aquaculture 2000, 187, 85–96. [Google Scholar] [CrossRef]
- Farhadian, O.; Yusoff, F.M.; Arshad, A. Population growth and production of Apocyclops dengizicus (Copepoda: Cyclopoida) fed on different diets. J. World Aquac. Soc. 2008, 39, 384–396. [Google Scholar] [CrossRef]
- Ribeiro, A.C.; Souza-Santos, L.P. Mass culture and offspring production of marine harpacticoid copepod Tisbe biminiensis. Aquaculture 2011, 321, 280–288. [Google Scholar] [CrossRef]
- Mubarak, A.S.; Jusadi, D.; Junior, M.Z.; Suprayudi, M.A. The population growth and the nutritional status of Moina macrocopa feed with rice bran and cassava bran suspensions. J. Akuakultur Indones. 2017, 16, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Amian, A.; Etile, R.; Aka, M.; Wandan, E.; Blé, C. Zooplankton diversity and abundance in extensive fish ponds during the rearing of tilapia Oreochromis niloticus juveniles fed with rice bran (West Africa, Côte d’Ivoire). Int. J. Fish. Aquat. Stud. 2018, 6, 131–136. [Google Scholar]
- Sulehria, A.Q.K.; Younus, I.; Hussain, A. Effect of artificial diets on the growth and survival of rotifers. Biologia 2010, 56, 31–37. [Google Scholar]
- Gyllenberg, G.; Lundqvist, G. Utilization of dissolved glucose by two copepod species. Ann. Zool. Fenn. 1978, 15, 323–327. [Google Scholar]
- Magouz, F.I.M.M.; Essa, M.A.; El-Shafei, A.; Mansour, A.T.; Mahmoud, S.m.; Ashour, M. Effect of extended feeding with live copepods, Oithona nana, and Artemia franciscana on the growth performance, intestine histology, and economic viability of european seabass (Dicentrarchus labrax) postlarvae. Fresenius Environ. Bull. 2021, 7106–7116. [Google Scholar]
- Ashour, M.; Mabrouk, M.M.; Ayoub, H.F.; El-Feky, M.M.; Zaki, S.Z.; Hoseinifar, S.H.; Rossi, W.; Van Doan, H.; El-Haroun, E.; Goda, A.M.-S. Effect of dietary seaweed extract supplementation on growth, feed utilization, hematological indices, and non-specific immunity of Nile Tilapia, Oreochromis niloticus, challenged with, Aeromonas hydrophila. J. Appl. Phycol. 2020, 32, 3467–3479. [Google Scholar] [CrossRef]
- Sorgeloos, P.; Baeza-Mesa, M.; Bossuyt, E.; Bruggeman, E.; Dobbeleir, J.; Versichele, D.; Laviña, E.; Bernardino, A. Culture of Artemia on rice bran: The conversion of a waste-product into highly nutritive animal protein. Aquaculture 1980, 21, 393–396. [Google Scholar] [CrossRef]
- Yamasaki, S.; Canto, J. Culture experiments on the harpacticoid copepod, Tisbintra elongata Mori, and evaluation of that species as a food organism for milkfish larvae. Memoirs Faculty Fish. Kagoshima Univ. 1980, 29, 275–291. [Google Scholar]
- Tseng, L.-C.; Dahms, H.-U.; Chen, Q.-C.; Hwang, J.-S. Copepod feeding study in the upper layer of the tropical South China Sea. Helgol. Mar. Res. 2009, 63, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.W.; Min, W.W.; Lin, H.J.; Chen, W. Population dynamics, protein content, and lipid composition of Brachionus plicatilis fed artificial macroalgal detritus and Nannochloropsis sp. diets. Aquaculture 2013, 380, 62–69. [Google Scholar] [CrossRef]
- El-Shenody, R.A.; Ashour, M.; Ghobara, M.M.E. Evaluating the chemical composition and antioxidant activity of three Egyptian seaweeds: Dictyota dichotoma, Turbinaria decurrens, and Laurencia obtusa. Brazil. J. Food Technol. 2019, 22, e2018203. [Google Scholar] [CrossRef] [Green Version]
- Schipp, G.R.; Bosmans, J.M.; Marshall, A.J. A method for hatchery culture of tropical calanoid copepods, Acartia spp. Aquaculture 1999, 174, 81–88. [Google Scholar] [CrossRef]
- Peck, M.A.; Holste, L. Effects of salinity, photoperiod and adult stocking density on egg production and egg hatching success in Acartia tonsa (Calanoida: Copepoda): Optimizing intensive cultures. Aquaculture 2006, 255, 341–350. [Google Scholar] [CrossRef]
- Chilmawati, D. The effect of different diet of phytoplankton cells on growth performance of Copepod, Oithona sp. in semi-mass culture. Aquat. Proc. 2016, 7, 39–45. [Google Scholar] [CrossRef]
- Ajiboye, O.; Yakubu, A.; Adams, T.; Olaji, E.; Nwogu, N. A review of the use of copepods in marine fish larviculture. Rev. Fish Biol. Fish. 2011, 21, 225–246. [Google Scholar] [CrossRef]
- Conceição, L.E.; Yúfera, M.; Makridis, P.; Morais, S.; Dinis, M.T. Live feeds for early stages of fish rearing. Aquac. Res. 2010, 41, 613–640. [Google Scholar] [CrossRef]
- Pan, Y.J.; Sadovskaya, I.; Hwang, J.S.; Souissi, S. Assessment of the fecundity, population growth and fatty acid composition of Apocyclops royi (Cyclopoida, Copepoda) fed on different microalgal diets. Aquac. Nutr. 2018, 24, 970–978. [Google Scholar] [CrossRef]
- Booman, M.; Forster, I.; Vederas, J.C.; Groman, D.B.; Jones, S.R. Soybean meal-induced enteritis in Atlantic salmon (Salmo salar) and Chinook salmon (Oncorhynchus tshawytscha) but not in pink salmon (O. gorbuscha). Aquaculture 2018, 483, 238–243. [Google Scholar] [CrossRef]
- FAO. FAOSTAT; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Martins, G.P.; Pezzato, L.E.; Guimaraes, I.G.; Padovani, C.R.; Mazini, B.S.M.; Barros, M.M. Antinutritional factors of raw soybean on growth and haematological responses of Nile tilapia. Boletim do Instituto de Pesca 2017, 43, 322–333. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch-composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Carli, A.; Mariottini, G.; Pane, L. Influence of nutrition on fecundity and survival in Tigriopus fulvus Fischer (Copepoda: Harpacticoida). Aquaculture 1995, 134, 113–119. [Google Scholar] [CrossRef]
- Støttrup, J.G.; Jensen, J. Influence of algal diet on feeding and egg-production of the calanoid copepod Acartia tonsa Dana. J. Exp. Mar. Biol. Ecol. 1990, 141, 87–105. [Google Scholar] [CrossRef]
- Bhat, N.A.; Rainaand, R.; Wanganeo, A. Ecological investigation of zooplankton abundance in the Bhoj wetland, Bhopal of central India: Impact of environmental variables. Int. J. Fish. Aquac. 2015, 7, 81–93. [Google Scholar]
- Rayner, T.A.; Jørgensen, N.O.; Blanda, E.; Wu, C.-H.; Huang, C.-C.; Mortensen, J.; Hwang, J.-S.; Hansen, B.W. Biochemical composition of the promising live feed tropical calanoid copepod Pseudodiaptomus annandalei (Sewell 1919) cultured in Taiwanese outdoor aquaculture ponds. Aquaculture 2015, 441, 25–34. [Google Scholar] [CrossRef]
- Dalsgaard, J.; John, M.S.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [PubMed]
- El-Sabaawi, R.; Dower, J.F.; Kainz, M.; Mazumder, A. Characterizing dietary variability and trophic positions of coastal calanoid copepods: Insight from stable isotopes and fatty acids. Mar. Biol. 2009, 156, 225–237. [Google Scholar] [CrossRef]
- Monroig, Ó.; Tocher, D.R.; Navarro, J.C. Biosynthesis of polyunsaturated fatty acids in marine invertebrates: Recent advances in molecular mechanisms. Mar. Drugs 2013, 11, 3998–4018. [Google Scholar] [CrossRef] [Green Version]
- Rasdi, N.W.; Qin, J.G.; Li, Y. Effects of dietary microalgae on fatty acids and digestive enzymes in copepod C yclopina kasignete, a potential live food for fish larvae. Aquac. Res. 2016, 47, 3254–3264. [Google Scholar] [CrossRef]
- Abbas, E.M.; Ali, F.S.; Desouky, M.G.; Ashour, M.; El-Shafei, A.; Maaty, M.M.; Sharawy, Z.Z. Novel comprehensive molecular and ecological study introducing coastal mud shrimp (Solenocera crassicornis) recorded at the Gulf of suez, Egypt. J. Mar. Sci. Eng. 2021, 9, 9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FA | Soybean | Yeast | Rice Bran | Starch |
|---|---|---|---|---|
| SFA | ||||
| C10:0 | 0.31 ± 0.006 d | 0.46 ± 0.17 a | 0.41 ± 0.006 b | 0.373 ± 0.003 c |
| C11:0 | 0.40 ± 0.006 c | 0.81 ± 0.075 a | 0.41 ± 0.023 c | 0.56 ± 0.006 b |
| C12:0 | 0.78 ± 0.003 ab | 0.88 ± 0.058 a | 0.70 ± 0.020 b | 0.52 ± 0.035 c |
| C13:0 | 2.17 ± 0.040 b | 2.87 ± 0.055 a | 2.39 ± 0.205 b | 3.10 ± 0.159 a |
| C14:0 | 10.51 ± 0.309 c | 17.80 ± 0.820 a | 14.15 ± 0.471 b | 16.10 ± 0.393 a |
| C15:0 | 0.91 ± 0.147 d | 2.60 ± 0.101 b | 1.99 ± 0.012 c | 11.80 ± 0.115 a |
| C16:0 | 38.54 ± 0.12 a | 24.78 ± 0.303 c | 30.48 ± 0.029 b | 24.83 ± 0.245 c |
| C17:0 | 0.55 ± 0.003 d | 1.92 ± 0.043 a | 1.16 ± 0.015 b | 0.69 ± 0.003 c |
| C18:0 | 21.88 ± 0.245 a | 11.57 ± 0.205 b | 9.76 ± 0.064 c | 8.12 ± 0.090 d |
| C20:4 | 0.83 ± 0.046 b | 2.65 ± 0.191 a | 2.43 ± 0.300 a | 2.22 ± 0.049 a |
| MUFA | ||||
| C14:1 | 12.35 ± 0.001 b | 15.16 ± 0.823 a | 15.58 ± 0.003 a | 13.93 ± 0.543 ab |
| C15:1 | 0.77 ± 0.015 b | 1.32 ± 0.001 b | 1.16 ± 0.479 b | 4.37 ± 0.150 a |
| C16:1 | 1.89 ± 0.020 b | 4.19 ± 0.592 a | 2.44 ± 0.032 b | 2.33 ± 0.040 b |
| C18:1c | 6.22 ± 0.150 c | 9.07 ± 0.297 b | 13.04 ± 0.069 a | 9.02 ± 0.433 b |
| C18:2c | 1.04 ± 0.104 c | 2.77 ± 0.015 a | 2.46 ± 0.006 b | 1.12 ± 0.012 c |
| PUFA | ||||
| C18:3ω3 | 0.83 ± 0.038 b | 1.18 ± 0.017 a | 1.20 ± 0.061 a | 0.91 ± 0.012 b |
| C20:5ω3 | 0.00 ± 0.000 b | 0.00 ± 0.000 b | 0.24 ± 0.006 a | 0.00 ± 0.000 b |
| ∑ SFA | 76.89 ± 0.297 a | 66.33 ± 0.479 c | 63.87 ± 0.647 d | 68.32 ± 0.084 b |
| ∑ MUFA | 22.27 ± 0.260 d | 32.49 ± 0.514 b | 34.68 ± 5.95 a | 30.77 ± 0.095 c |
| ∑ PUFA | 0.83 ± 0.037 c | 1.18 ± 0.017 b | 1.45 ± 0.055 a | 0.91 ± 0.012 c |
| SFA/UFA | 3.33 ± 0.058 a | 1.97 ± 0.043 c | 1.77 ± 0.052 d | 2.16 ± 0.009 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magouz, F.I.; Essa, M.A.; Matter, M.; Tageldein Mansour, A.; Alkafafy, M.; Ashour, M. Population Dynamics, Fecundity and Fatty Acid Composition of Oithona nana (Cyclopoida, Copepoda), Fed on Different Diets. Animals 2021, 11, 1188. https://doi.org/10.3390/ani11051188
Magouz FI, Essa MA, Matter M, Tageldein Mansour A, Alkafafy M, Ashour M. Population Dynamics, Fecundity and Fatty Acid Composition of Oithona nana (Cyclopoida, Copepoda), Fed on Different Diets. Animals. 2021; 11(5):1188. https://doi.org/10.3390/ani11051188
Chicago/Turabian StyleMagouz, Fawzy I., Mohamed A. Essa, Mustafa Matter, Abdallah Tageldein Mansour, Mohamed Alkafafy, and Mohamed Ashour. 2021. "Population Dynamics, Fecundity and Fatty Acid Composition of Oithona nana (Cyclopoida, Copepoda), Fed on Different Diets" Animals 11, no. 5: 1188. https://doi.org/10.3390/ani11051188
APA StyleMagouz, F. I., Essa, M. A., Matter, M., Tageldein Mansour, A., Alkafafy, M., & Ashour, M. (2021). Population Dynamics, Fecundity and Fatty Acid Composition of Oithona nana (Cyclopoida, Copepoda), Fed on Different Diets. Animals, 11(5), 1188. https://doi.org/10.3390/ani11051188