
Karyotype Evolution and Genomic Organization of Repetitive DNAs in the Saffron Finch, Sicalis flaveola (Passeriformes, Aves)

 ,
,  ,
,  ,
,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Chromosome Preparations
2.2. Diploid Number, C and G-Banding
2.3. Fluorescence In Situ Hybridization (FISH) with Simple Short Repeat Probes (SSR) and Bacterial Artificial Chromosomes (BAC) Probes
2.4. Microscopic Analysis and Image Capturing
3. Results
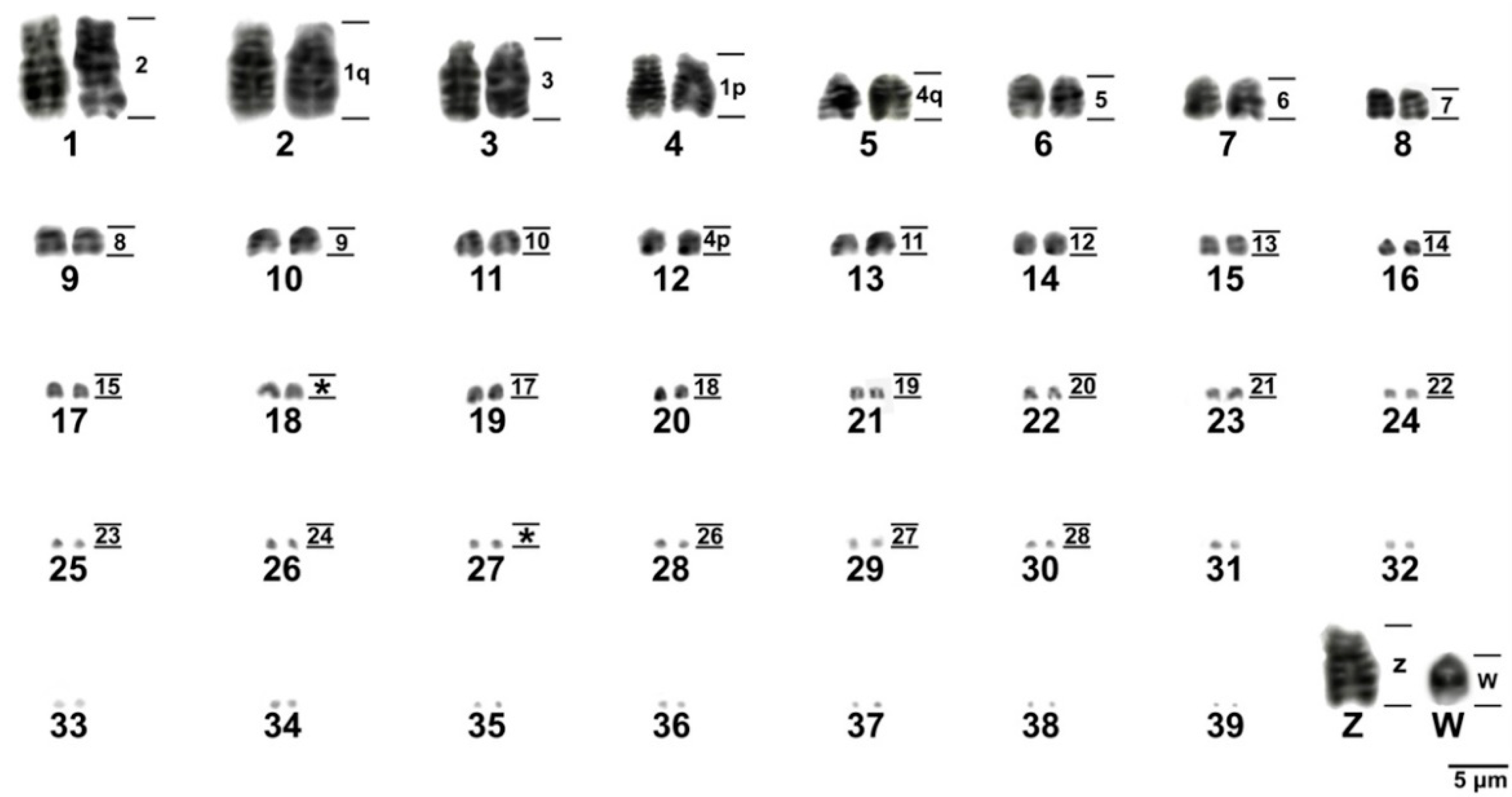
3.1. Karyotype Description, C and G-Banding
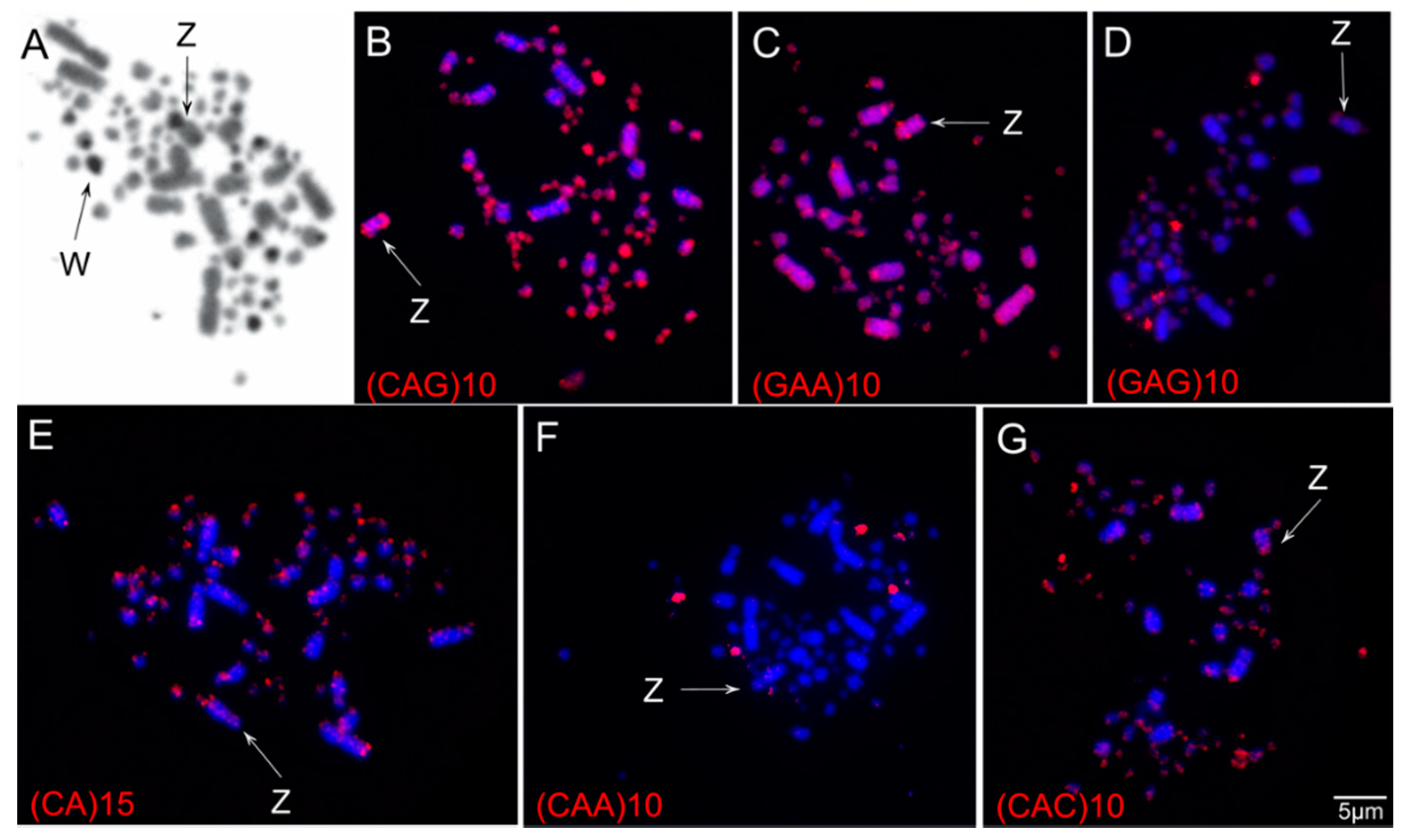
3.2. Chromosomal Distribution of Simple Short Repeats (SSRs)
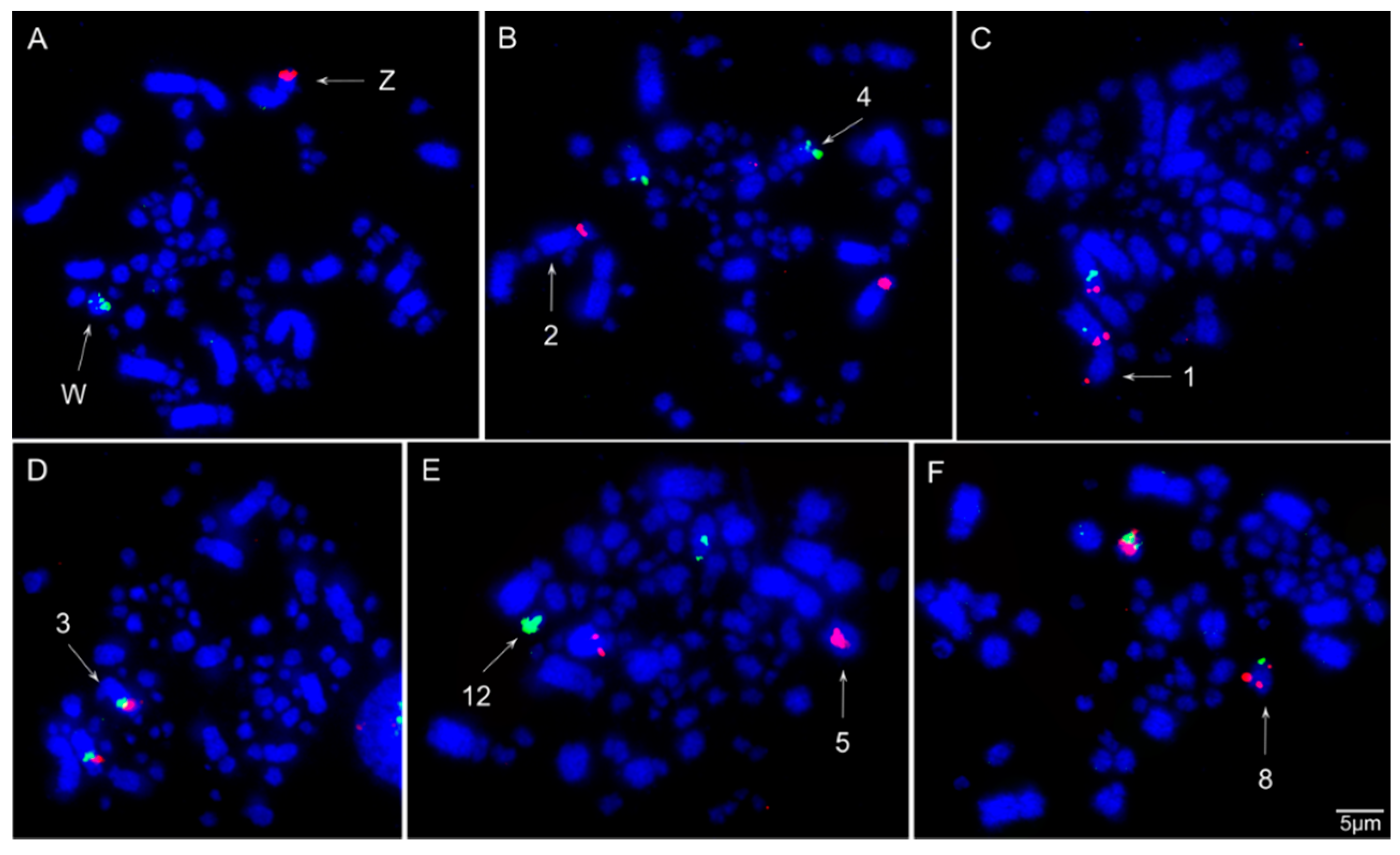
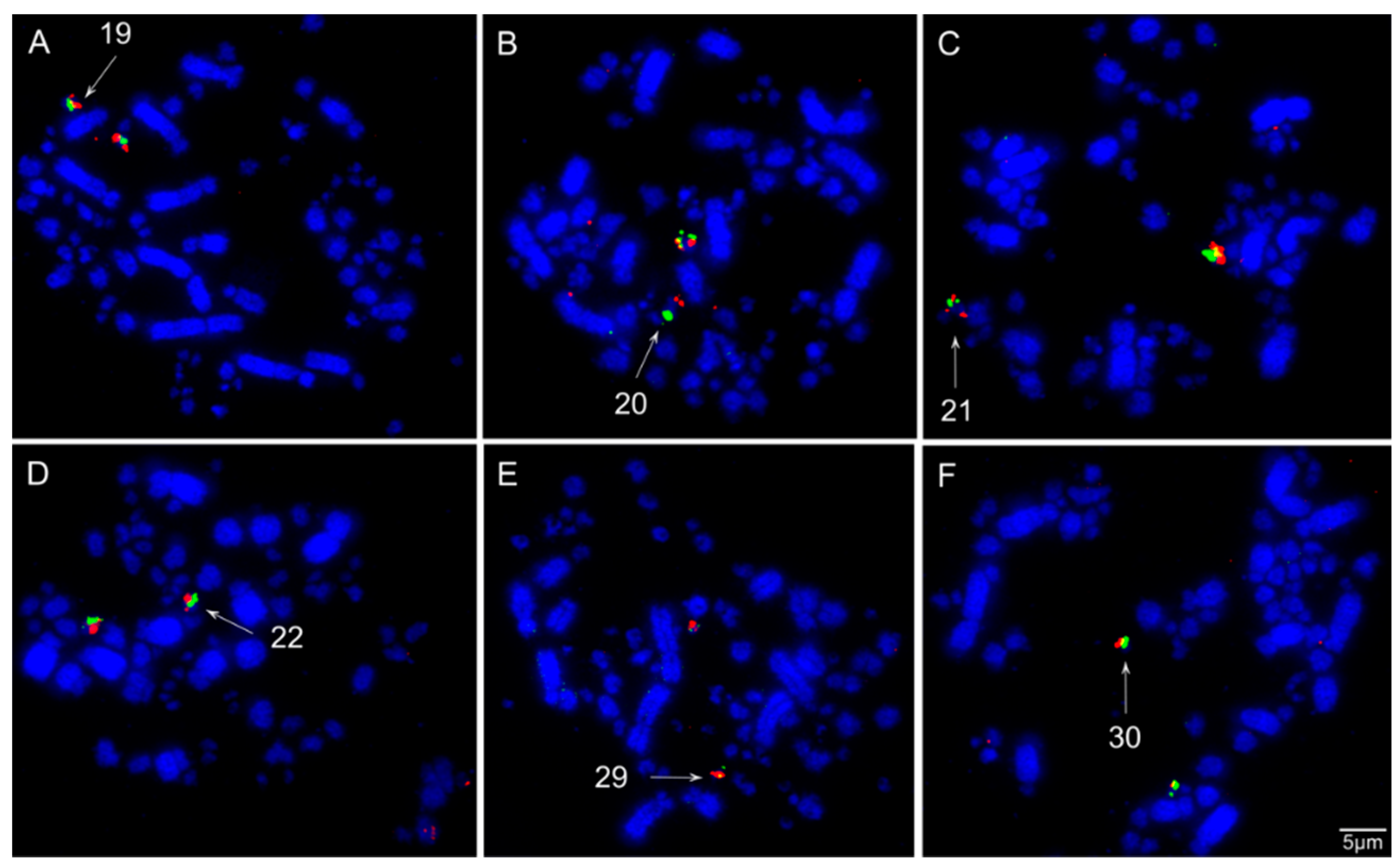
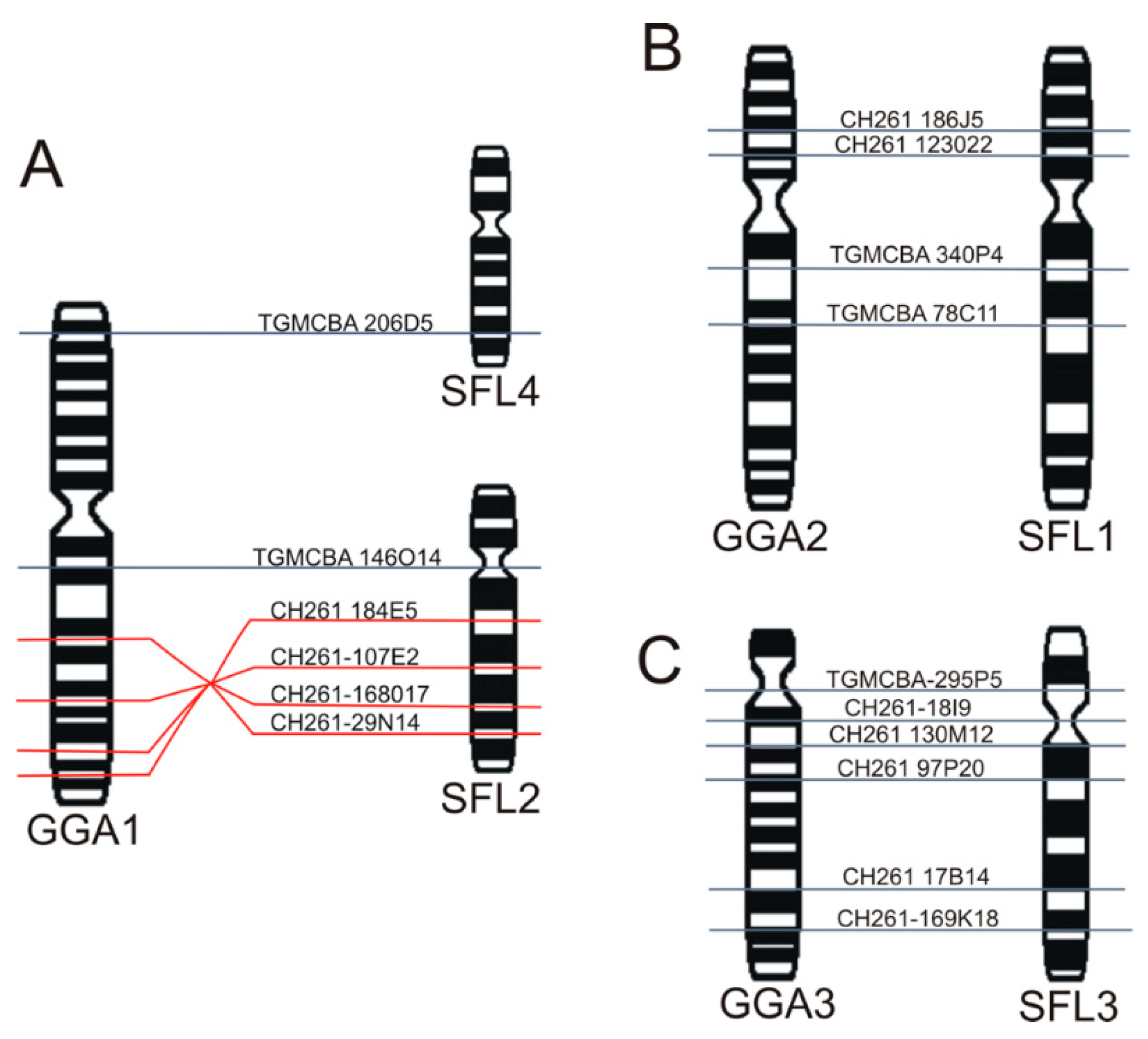
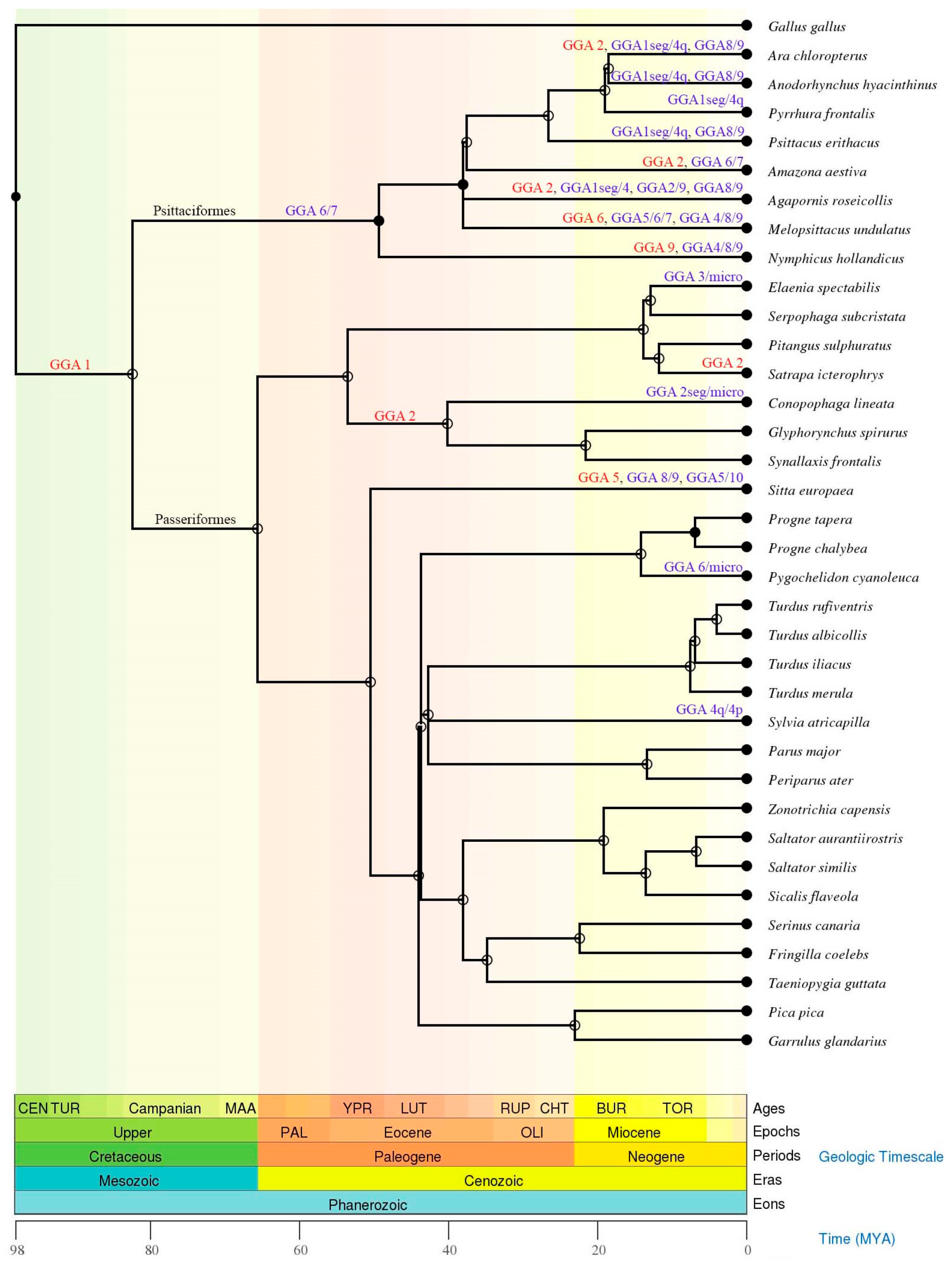
3.3. Chromosomal Homology Between Chicken and Sicalis flaveola
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burns, K.J.; Shultz, A.J.; Title, P.O.; Mason, N.A.; Barker, F.K.; Klicka, J.; Lanyon, S.M.; Lovette, I.J. Phylogenetics and diversification of tanagers (Passeriformes: Thraupidae), the largest radiation of Neotropical songbirds. Mol. Phylogenetics Evol. 2014, 75, 41–77. [Google Scholar] [CrossRef] [PubMed]
- Gill, F. IOC World Bird List; (v11.1); Gill, F., Donsker, D., Rasmussen, P., Eds.; Gill & Wright: London, UK, 2021; Available online: https://www.worldbirdnames.org/ioc-lists/crossref/ (accessed on 21 December 2020).
- Ericson, P.G.P.; Irestedt, M.; Johansson, U.S. Evolution, biogeography, and patterns of diversification in passerine birds. J. Avian Biol. 2003, 34, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Ridgely, R.S.; Tudor, G. Field Guide to the Songbirds of South America: The Passerines; University of Texas Press: Austin, TX, USA, 2009. [Google Scholar]
- Christidis, L.; Boles, W. Systematics and Taxonomy of Australian Birds; Csiro Publishing: Clayton, Australia, 2008. [Google Scholar]
- BirdLife International. Species Factsheet: Sicalis Flaveola. Available online: http://www.birdlife.org (accessed on 21 December 2020).
- Degrandi, T.M.; Barcellos, S.A.; Costa, A.L.; Garnero, A.D.; Hass, I.; Gunski, R.J. Introducing the Bird Chromosome Database: An Overview of Cytogenetic Studies in Birds. Cytogenet. Genome Res. 2020, 160, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, B.; Nogueira, D.M.; Monsores, D.W.; Souza, L.M. Chromosome study in two Aratinga species (A. guarouba and A. acuticaudata) (Psittaciformes). Braz. J. Genet. 1997, 20. [Google Scholar] [CrossRef]
- Cabanne, G.S.; Gunski, R.J.; Contreras, J.R. Primeros Resultados de Estudios Citogenéticos en Saltator coerulescens y Saltator similis (Aves: Emberizidae); VI Jornada de Ciências Naturales del Litoral: Corrientes, Argentina, 1997. [Google Scholar]
- Dos Santos, M.S.; Kretschmer, R.; Silva, F.A.O.; Ledesma, M.A.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Garnero, A.D.V.; de Oliveira, E.H.C.; Gunski, R.J. Intrachromosomal rearrangements in two representatives of the genus Saltator (Thraupidae, Passeriformes) and the occurrence of heteromorphic Z chromosomes. Genetica 2014, 143, 535–543. [Google Scholar] [CrossRef]
- Kretschmer, R.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Karyotype evolution in birds: From conventional staining to chromosome painting. Genes 2018, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volker, M.; Backstrom, N.; Skinner, B.M.; Langley, E.J.; Bunzey, S.K.; Ellegren, H.; Griffin, D.K. Copy number variation, chromosome rearrangement, and their association with recombination during avian evolution. Genome Res. 2010, 20, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Warren, W.C.; Clayton, D.F.; Ellegren, H.; Arnold, A.P.; Hillier, L.W.; Künstner, A.; Searle, S.; White, S.; Vilella, A.J.; Fairley, S.; et al. The genome of a songbird. Nature 2010, 464, 757–762. [Google Scholar] [CrossRef]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; Furo, I.O.; O’Brien, P.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Molecular cytogenetic characterization of multiple intrachromosomal rearrangements in two representatives of the genus Turdus (Turdidae, Passeriformes). PLoS ONE 2014, 9, e103338. [Google Scholar] [CrossRef]
- Kretschmer, R.; de Oliveira, E.H.C.; dos Santos, M.S.; Furo, I.O.; O’Brien, P.C.; Ferguson-Smith, M.A.; Garnero, A.D.V.; Gunski, R.J. Chromosome mapping of the large elaenia (Elaenia spectabilis): Evidence for a cytogenetic signature for passeriform birds? Biol. J. Linn. Soc. 2015, 115, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Kretschmer, R.; de Lima, V.L.C.; de Souza, M.S.; Costa, A.L.; O’Brien, P.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; Gunski, R.J.; Garnero, A.D.V. Multidirectional chromosome painting in Synallaxis frontalis (Passeriformes, Furnariidae) reveals high chromosomal reorganization, involving fissions and inversions. Comp. Cytogenet. 2018, 12, 97–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, M.S.; Kretschmer, R.; Frankl-Vilches, C.; Bakker, A.; Gahr, M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Comparative cytogenetics between two important songbird, models: The zebra finch and the canary. PLoS ONE 2017, 12, e0170997. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, B.S.; Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.; de Oliveira, E.H.C. Chromosome painting in tyrant flycatchers confirms a set of inversions shared by Oscines and Suboscines (Aves, Passeriformes). Cytogenet. Genome Res. 2017, 153, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Burt, D.W. Parameters of the chicken genome (Gallus gallus). Anim. Genet. 1998, 29, 290–294. [Google Scholar] [CrossRef]
- Smith, J.; Bruley, C.K.; Paton, I.R.; Dunn, I.; Jones, C.T.; Windsor, D.; Morrice, D.R.; Law, A.S.; Masabanda, J.; Sazanov, A.; et al. Differences in gene density on chicken macrochromosomes and microchromosomes. Anim. Genet. 2000, 31, 96–103. [Google Scholar] [CrossRef]
- Guttenbach, M.; Nanda, I.; Feichtinger, W.; Masabanda, J.S.; Griffin, D.K.; Schmid, M. Comparative chromosome painting of chicken autosomal paints 1–9 in nine different bird species. Cytogenet. Genome Res. 2003, 103, 173–184. [Google Scholar] [CrossRef]
- O’Connor, R.E.; Kiazim, L.; Skinner, B.; Fonseka, G.; Joseph, S.; Jennings, R.; Larkin, D.M.; Griffin, D.K. Patterns of microchromosome organization remain highly conserved throughout avian evolution. Chromosoma 2019, 128, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Ribas, T.F.A.; Pieczarka, J.C.; Griffin, D.K.; Kiazim, L.G.; Nagamachi, C.Y.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Aleixo, A.; O’Connor, R.E. Analysis of multiple chromosomal rearrangements in the genome of Willisornis vidua using BAC-FISH and chromosome painting on a supposed conserved karyotype. BMC Ecol. Evol. 2021, 21, 34. [Google Scholar] [CrossRef]
- Nishida, C.; Ishijima, J.; Kosaka, A.; Tanabe, H.; Habermann, F.A.; Griffin, D.K.; Matsuda, Y. Characterization of chromosome structures of Falconinae (Falconidae, Falconiformes, Aves) by chromosome painting and delineation of chromosome rearrangements during their differentiation. Chromosome Res. 2008, 16, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Nanda, I.; Karl, E.; Griffin, D.K.; Schartl, M.; Schmid, M. Chromosome repatterning in three representative parrots (Psittaciformes) inferred from comparative chromosome painting. Cytogenet. Genome Res. 2007, 117, 43–53. [Google Scholar] [CrossRef]
- Joseph, S.; O’Connor, R.E.; Al Mutery, A.F.; Watson, M.; Larkin, D.M.; Griffin, D.K. Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation. Diversity 2018, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.M.; Pereira, J.; Garnero, A.D.V.; Gunski, R.J.; O’Connor, R.E.; Griffin, D.K.; Gomes, A.J.B.; Ferguson-Smith, M.A.; et al. Chromosomal evolution in the phylogenetic context in Neotropical Psittacidae with emphasis on a species with high karyotypic reorganization (Myiopsitta monachus). Front. Genet. 2020, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; de Freitas, T.R.O.; Toma, G.A.; Cioffi, M.B.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K. Chromosomal analysis in Crotophaga ani (Aves, Cuculiformes) reveals extensive genomic reorganization and an unusual Z-autosome Robertsonian translocation. Cells 2021, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Souza, M.S.; Furo, I.d.O.; Romanov, M.N.; Gunski, R.J.; Garnero, A.d.V.; de Freitas, T.R.O.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K. Interspecies Chromosome Mapping in Caprimulgiformes, Piciformes, Suliformes, and Trogoniformes (Aves): Cytogenomic Insight into Microchromosome Organization and Karyotype Evolution in Birds. Cells 2021, 10, 826. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Poltronieri, J.; Bertollo, L.A.C.; Artoni, R.F.; Liehr, T.; Cioffi, M.B. Chromosomal Mapping of Repetitive DNAs in Triportheus trifurcatus (Characidae, Characiformes): Insights into the Differentiation of the Z and W Chromosomes. PLoS ONE 2014, 9, e90946. [Google Scholar] [CrossRef]
- George, C.M.; Alani, E. Multiple cellular mechanisms prevent chromosomal rearrangements involving repetitive DNA. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 297–313. [Google Scholar] [CrossRef]
- Dunn, M.J.; Anderson, M.Z. To Repeat or Not to Repeat: Repetitive Sequences Regulate Genome Stability in Candida albicans. Genes 2019, 10, 866. [Google Scholar] [CrossRef] [Green Version]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Matsubara, K.; O’Meally, D.; Azad, B.; Georges, A.; Sarre, S.D.; Graves, J.A.M.; Matsuda, Y.; Ezaz, T. Amplification of microsatellite repeat motifs is associated with the evolutionary differentiation and heterochromatinization of sex chromosomes in Sauropsida. Chromosoma 2016, 125, 111–123. [Google Scholar] [CrossRef]
- de Oliveira, T.D.; Kretschmer, R.; Bertocchi, N.A.; Degrandi, T.M.; de Oliveira, E.H.C.; Cioffi, M.B.; Garnero, A.D.V.; Gunski, R.J. Genomic organization of repetitive DNA in woodpeckers (Aves, Piciformes): Implications for karyotype and ZW sex chromosome differentiation. PLoS ONE 2017, 12, e0169987. [Google Scholar] [CrossRef]
- Furo, I.O.; Kretschmer, R.; dos Santos, M.S.; Carvalho, C.A.; Gunski, R.J.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Cioffi, M.B.; de Oliveira, E.H.C. Chromosomal Mapping of Repetitive DNAs in Myiopsitta monachus and Amazona aestiva (Psittaciformes, Psittacidae), with emphasis on the sex chromosomes. Cytogenet. Genome Res. 2017, 151, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Oliveira, T.D.; Furo, I.O.; Silva, F.A.O.; Gunski, R.J.; Garnero, A.V.; Cioffi, M.B.; de Oliveira, E.H.C.; de Freitas, T.R.O. Repetitive DNAs and shrink genomes: A chromosomal analysis in nine Columbidae species (Aves, Columbiformes). Genet. Mol. Biol. 2018, 41, 98–106. [Google Scholar] [CrossRef]
- Barcellos, S.; Kretschmer, R.; de Souza, M.S.; Costa, A.L.; Degrandi, T.M.; dos Santos, M.S.; de Oliveira, E.H.C.; Cioffi, M.B.; Gunski, R.J.; Garnero, A.V. Karyotype Evolution and Distinct Evolutionary History of the W Chromosomes in Swallows (Aves, Passeriformes). Cytogenet. Genome Res. 2019, 158, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Gunski, R.J.; Kretschmer, R.; de Souza, M.S.; Furo, I.O.; Barcellos, S.; Costa, A.L.; Cioffi, M.B.; de Oliveira, E.H.C.; Garnero, A.V. Evolution of bird sex chromosomes narrated by repetitive sequences: Unusual W chromosome enlargement in Gallinula melanops (Aves: Gruiformes: Rallidae). Cytogenet. Genome Res. 2019, 158, 152–159. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.S.; Kretschmer, R.; Barcellos, A.S.; Costa, A.L.; Cioffi, M.B.; de Oliveira, E.H.C.; Garnero, A.D.V.; Gunski, R.J. Repeat Sequence Mapping Shows Different W Chromosome Evolutionary Pathways in Two Caprimulgiformes Families. Birds 2020, 1, 19–34. [Google Scholar] [CrossRef]
- De Lucca, E.J. Cariótipo de oito espécies de Aves. Rev. Bras. Biol. 1974, 34, 387–392. [Google Scholar]
- De Lucca, E.J.; Chamma, L. Estudo do complemento cromossômico de 11 espécies de aves das ordens Columbiformes, Passeriformes e Tinamiformes. Braz. J. Med. Biol. Res. 1977, 10, 97–105. [Google Scholar]
- Carvalho, M.V.P. Estudos Citogenéticos na Família Fringillidae (Passeriformess-Aves). MSc Thesis, Programa de pós-graduação em Genética e Biologia Molecular, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil, 1989. [Google Scholar]
- Sasaki, M.; Ikeuchi, T.; Makino, S. A feather pulp culture technique for avian chromosomes, with notes on the chromosomes of the peafowl and the ostrich. Experientia 1968, 24, 1292–1293. [Google Scholar] [CrossRef] [PubMed]
- Guerra, M.S. Reviewing the chromosome nomenclature of Levan et al. Brazil. J. Genet. 1986, 9, 741–743. [Google Scholar]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Schnedl, W. Analysis of the human karyotype using a reassociation technique. Chromosoma 1971, 34, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.L.; Lopes, C.F.; de Souza, M.S.; Barcellos, S.A.; Vielmo, P.G.; Gunski, R.J.; Garnero, A.D.V. Comparative cytogenetics in three species of Wood-Warblers (Aves: Passeriformes: Parulidae) reveal divergent banding patterns and chromatic heterogeneity for the W chromosome. Caryologia, in press.
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite accumulation in the Y chromosome in Silene latifolia. Genome 2008, 51, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Ladjali-Mohammedi, K.; Bitgood, J.J.; Tixier-Boichard, M.; de Leon, F.P. International system for standardized avian karyotypes (ISSAK): Standardized banded karyotypes of the domestic fowl (Gallus domesticus). Cytogenet. Genome Res. 1999, 86, 271–276. [Google Scholar] [CrossRef]
- Correia, V.C.; Garnero, A.D.V.; dos Santos, L.P.; da Silva, R.R.; Barbosa, M.O.; Bonifacio, H.L.; Gunski, R.J. Alta similaridade cariotípica na família Emberizidae (Aves: Passeriformes). Biosci. J. 2009, 25, 99–111. [Google Scholar]
- Fulgione, D.; Aprea, G.; Milone, M.; Odierna, G. Chromosomes and heterochromatin of the Italian sparrow (Passer italiae, Vieillot 1817), a taxon of presumed hybrid origins. Folia Zool. 2000, 49, 199–204. [Google Scholar]
- Christid, L. Chromosomal evolution in finches and their allies (families: Ploceidae, Fringillidae, and Emberizidae). Can. J. Genet. Cytol. 1986, 28, 762–769. [Google Scholar] [CrossRef]
- Christid, L. Chromosomal evolution within the family Estrildidae (Aves) II. The Lonchurae. Genetica 1986, 71, 99–113. [Google Scholar] [CrossRef]
- Marchal, J.A.; Acosta, M.J.; Neitzel, H.; Sperling, K.; Bullejos, M.; Díaz de la Guardia, R.; Sanchez, A. X chromosome painting in Microtus: Origin and evolution of the giant sex chromosomes. Chromosome Res. 2004, 12, 767–776. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Moreira-Filho, O.; Almeida-Toledo, L.F.; Bertollo, L.A.C. The contrasting role of heterochromatin in the differentiation of sex chromosomes: An overview from Neotropical fishes. J. Fish Biol. 2012, 80, 2125–2139. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Liehr, T.; Troy, W.P.; Cioffi, M.B. W chromosome dynamics in Triportheus species (Characiformes, Triportheidae)—An ongoing process narrated by repetitive sequences. J. Hered. 2016, 107, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Kejnovsky, E.; Hobza, R.; Cermak, T.; Kubat, Z.; Vyskot, B. The role of repetitive DNA in structure and evolution of sex chromosomes in plants. Heredity 2009, 102, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Ezaz, T.; Sarre, S.D.; O’Meally, D.; Marshall Graves, J.A.; Georges, A. Sex chromosome evolution in lizards: Independent origins and rapid transitions. Cytogenet. Genome Res. 2009, 127, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Castagnone-Sereno, P.; Danchin, E.G.; Deleury, E.; Guillemaud, T.; Malausa, T.; Abad, P. Genome-wide survey and analysis of microsatellites in nematodes, with a focus on the plant-parasitic species Meloidogyne incognita. BMC Genom. 2010, 11, 598. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Furo, I.O.; Cioffi, M.B.; Gunski, R.J.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Freitas, T.R.O.; de Oliveira, E.H.C. Extensive chromosomal fissions and repetitive DNA accumulation shaped the atypical karyotypes of two Ramphastidae (Aves: Piciformes) species. Biol. J. Linn. Soc. 2020, 130, 839–849. [Google Scholar] [CrossRef]
- Johansson, U.S.; Fjeldså, J.; Bowie, R.C.K. Phylogenetic relationships within Passerida (Aves: Passeriformes): A review and a new molecular phylogeny based on three nuclear intron markers. Mol. Phylogenetics Evol. 2008, 48, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Hackett, S.J.; Kimball, R.T.; Reddy, S.; Bowie, R.C.K.; Braun, E.L.; Braun, M.J.; Chojnowski, J.L.; Cox, W.A.; Han, K.L.; Harshman, J.; et al. A phylogenomic study of birds reveals their evolutionary history. Science 2008, 320, 1763–1768. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef] [Green Version]
- Prum, R.O.; Berv, J.S.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef]
- Romanov, M.N.; Farré, M.; Lithgow, P.E.; Fowler, K.E.; Skinner, B.M.; O’Connor, R.; Fonseka, G.; Backström, N.; Matsuda, Y.; Nishida, C.; et al. Reconstruction of gross avian genome structure, organization and evolution suggests that the chicken lineage most closely resembles the dinosaur avian ancestor. BMC Genom. 2014, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1937. [Google Scholar]
- Krimbas, C.B.; Powell, J.R. Drosophila Inversion Polymorphism; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Hoffmann, A.A.; Sgrò, C.M.; Weeks, A.R. Chromosomal inversion polymorphisms and adaptation. Trends Ecol. Evol. 2004, 19, 482–488. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | SSRs | ||
|---|---|---|---|
| (CAA)10 | (CAG)10 | (CA)15 | |
| S. flaveola, 2n = 80 1 | Three pairs of micros | Scattered signals in all chromosomes but strong signals on the telomere regions of macros and micros, and in the Zp | Scattered signals in all chromosomes but strong signals on the telomere regions of macros and in the micros |
| P. cyanoleuca, 2n = 76 2 | Telomere of Wq | Telomere of Wpq | Telomere of Wq |
| P. tapera, 2n = 76 2 | Telomere of 1q | Wq | Telomere of 1q, 2q, Wq; Pericentromeric region of 6, 7, Wq |
| P. chalybea, 2n = 76 2 | Telomere of 1pq, 2q, Wq; pericentromeric region of 1pq | - | Telomere of 1pq, 2q, 4q, Wq; pericentromeric region of 1pq, Wpq |
| M. monachus, 2n = 48 3 | Wq | Telomere region of 1p, 2q, 3q, 4pq, 5q, 6p, Zp; pericentromeric region of 1q, 7q, Wq; centromeric region 1-9; all micros | - |
| A. aestiva, 2n = 70 3 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kretschmer, R.; Rodrigues, B.S.; Barcellos, S.A.; Costa, A.L.; Cioffi, M.d.B.; Garnero, A.d.V.; Gunski, R.J.; de Oliveira, E.H.C.; Griffin, D.K. Karyotype Evolution and Genomic Organization of Repetitive DNAs in the Saffron Finch, Sicalis flaveola (Passeriformes, Aves). Animals 2021, 11, 1456. https://doi.org/10.3390/ani11051456
Kretschmer R, Rodrigues BS, Barcellos SA, Costa AL, Cioffi MdB, Garnero AdV, Gunski RJ, de Oliveira EHC, Griffin DK. Karyotype Evolution and Genomic Organization of Repetitive DNAs in the Saffron Finch, Sicalis flaveola (Passeriformes, Aves). Animals. 2021; 11(5):1456. https://doi.org/10.3390/ani11051456
Chicago/Turabian StyleKretschmer, Rafael, Benilson Silva Rodrigues, Suziane Alves Barcellos, Alice Lemos Costa, Marcelo de Bello Cioffi, Analía del Valle Garnero, Ricardo José Gunski, Edivaldo Herculano Corrêa de Oliveira, and Darren K. Griffin. 2021. "Karyotype Evolution and Genomic Organization of Repetitive DNAs in the Saffron Finch, Sicalis flaveola (Passeriformes, Aves)" Animals 11, no. 5: 1456. https://doi.org/10.3390/ani11051456
APA StyleKretschmer, R., Rodrigues, B. S., Barcellos, S. A., Costa, A. L., Cioffi, M. d. B., Garnero, A. d. V., Gunski, R. J., de Oliveira, E. H. C., & Griffin, D. K. (2021). Karyotype Evolution and Genomic Organization of Repetitive DNAs in the Saffron Finch, Sicalis flaveola (Passeriformes, Aves). Animals, 11(5), 1456. https://doi.org/10.3390/ani11051456