
RNA Profiles of the Korat Chicken Breast Muscle with Increased Carnosine Content Produced through Dietary Supplementation with β-Alanine or L-Histidine

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. RNA Extraction
2.3. Library Preparation and Sequencing
2.4. Sequencing Quality Assessment and Differential Expression Analysis
2.5. Gene Ontology and Pathway Enrichment Analysis
2.6. Verification of RNA-Seq Results by Quantitative PCR (qPCR)
3. Results
3.1. Quality of RNA-Seq Reads
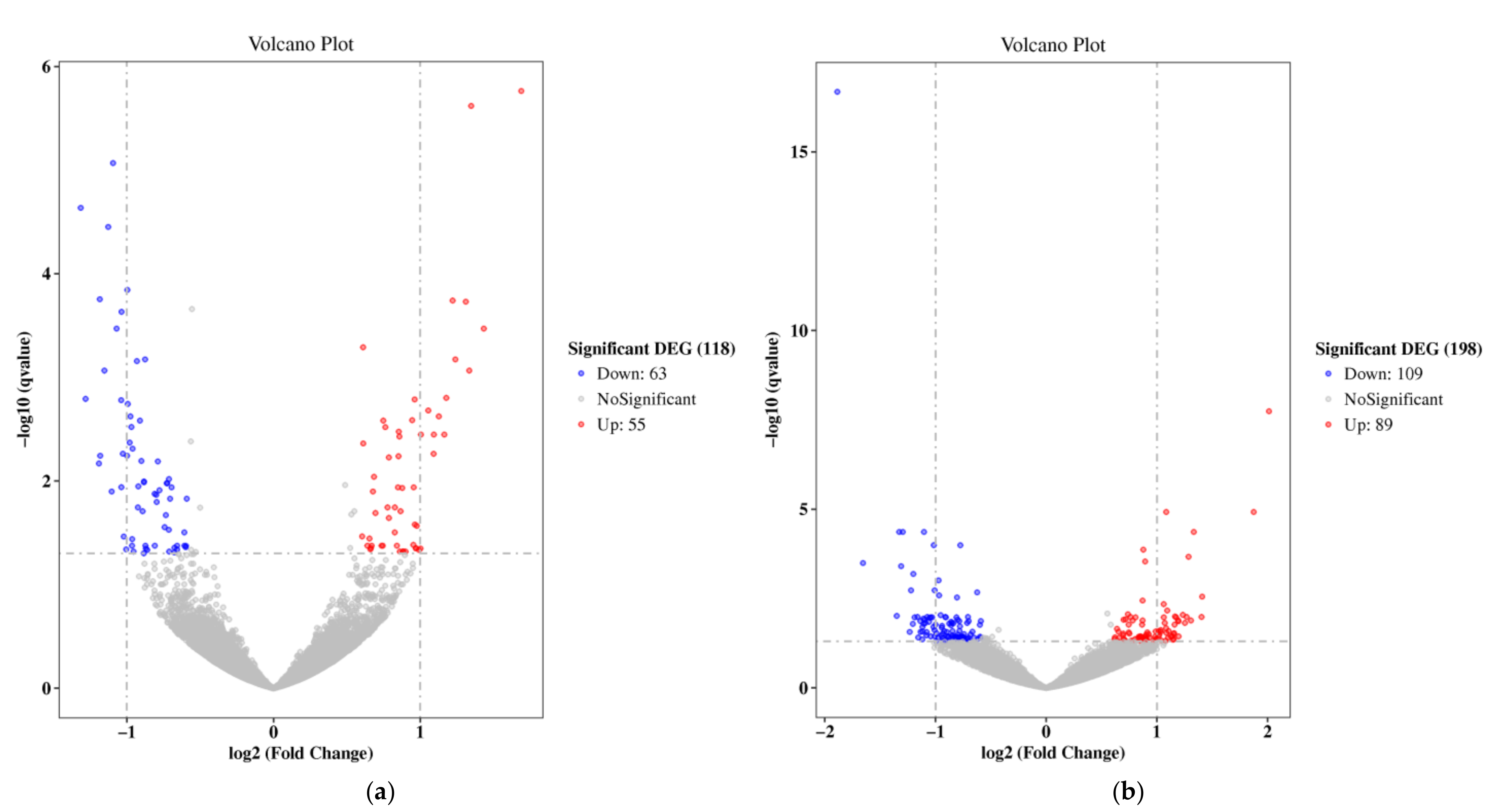
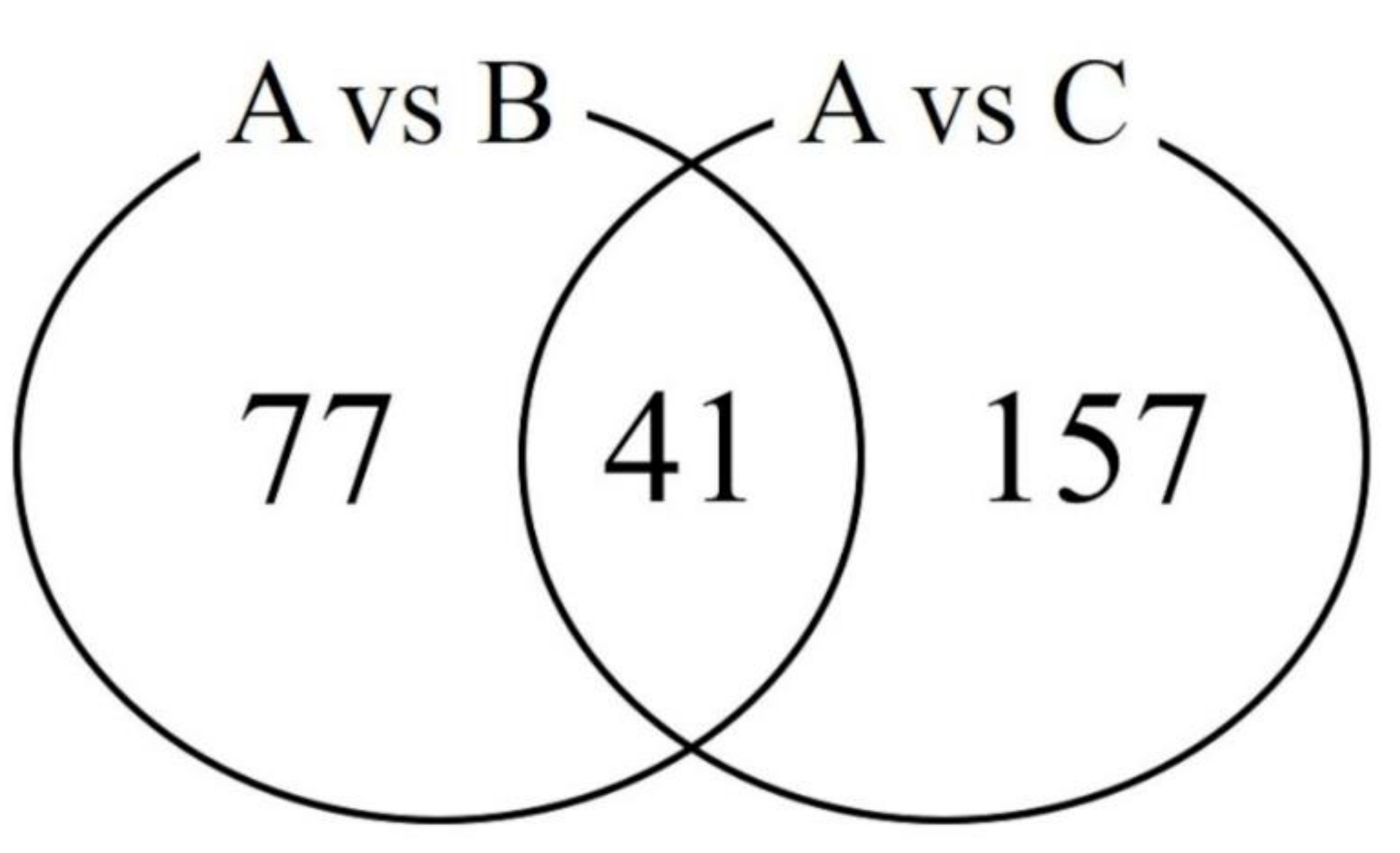
3.2. Significantly Differentially Expressed Transcripts
3.3. GO Annotation and KEGG Pathways Analyses of DEGs
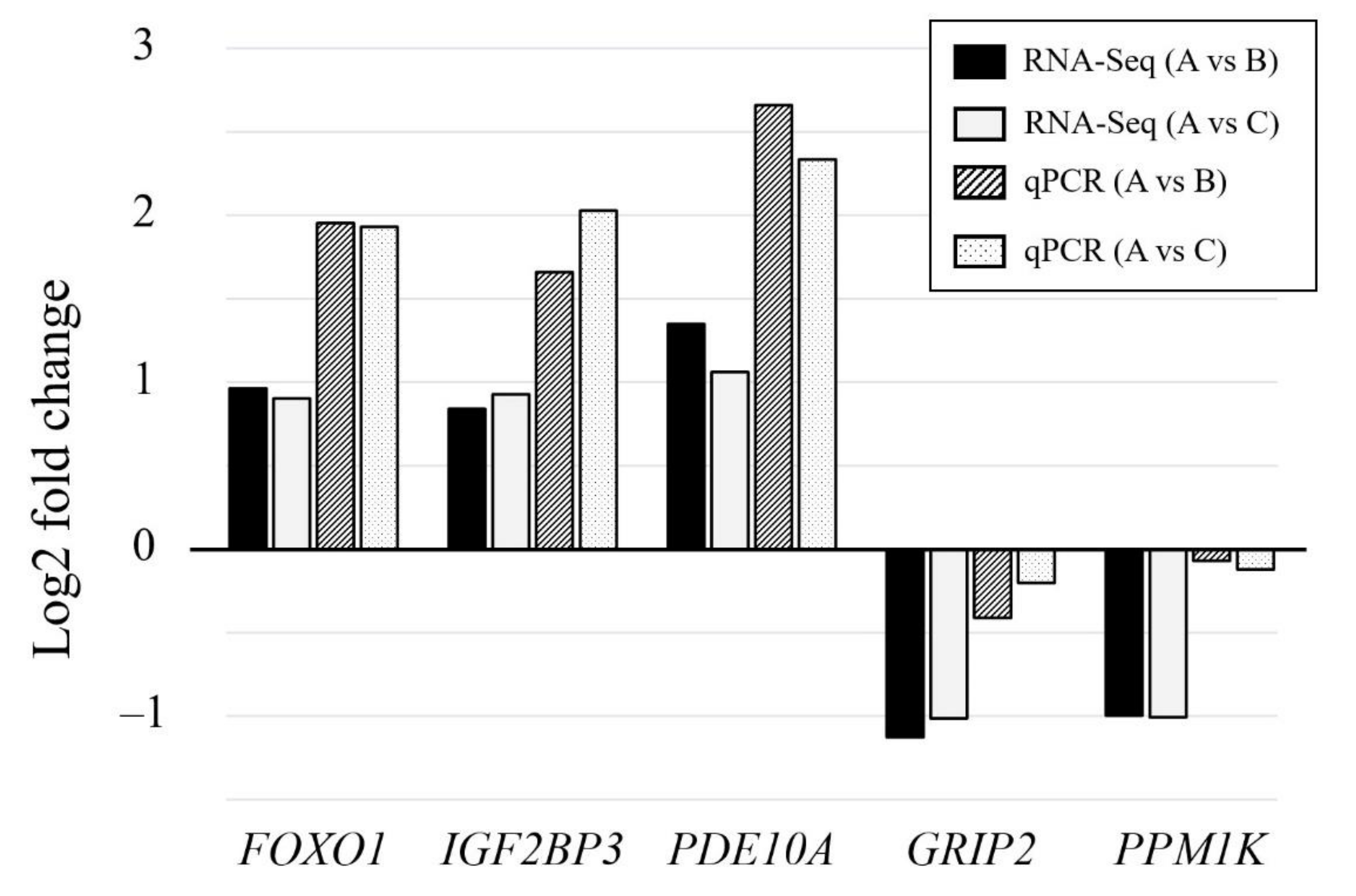
3.4. Validation of DEGs by qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Maliwan, P.; Khempaka, S.; Molee, W. Evaluation of various feeding programmes on growth performance, carcass and meat qualities of Thai indigenous crossbred chickens. S. Afr. J. Anim. Sci. 2017, 47, 16–25. [Google Scholar]
- Intarapichet, K.O.; Maikhunthod, B. Genotype and gender differences in carnosine extracts and antioxidant activities of chicken breast and thigh meats. Meat Sci. 2005, 71, 634–642. [Google Scholar]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar]
- Hipkiss, A.R. Carnosine and its possible roles in nutrition and health. Adv. Food Nutr. Res. 2009, 57, 87–154. [Google Scholar]
- Kai, S.; Watanabe, G.; Kubota, M.; Kadowaki, M.; Fujimura, S. Effect of dietary histidine on contents of carnosine and anserine in muscles of broilers. Anim. Sci. J. 2015, 86, 541–546. [Google Scholar]
- Qi, B.; Wang, J.; Ma, Y.; Wu, S.; Qi, G.; Zhang, H. Effect of dietary β-alanine supplementation on growth performance, meat quality, carnosine content, and gene expression of carnosine-related enzymes in broilers. Poult. Sci. 2018, 97, 1220–1228. [Google Scholar]
- Suwanvichanee, C.; Kubota, S.; Molee, W.; Yongsawatdigul, J.; Molee, A. Effect of β-Alanine and L-Histidine supplementation on growth performance and meat quality in female Korat chicken. In Proceedings of the 2nd International Conference on Tropical Animal Science and Production and the 2nd International Conference on Native Chicken, Nakhon Ratchasima, Thailand, 9–12 July 2019; p. 163. [Google Scholar]
- Maltin, C.; Balcerzak, D.; Tilley, R.; Delday, M. Determinants of meat quality: Tenderness. Proc. Nutr. Soc. 2003, 62, 337–347. [Google Scholar]
- Ismail, I.; Joo, S.T. Poultry meat quality in relation to muscle growth and muscle fiber characteristics. Korean J. Food Sci. Anim. Resour. 2017, 37, 873–883. [Google Scholar]
- Shu, J.T.; Zhang, M.; Shan, Y.J.; Xu, W.J.; Chen, K.W.; Li, H.F. Analysis of the genetic effects of CAPN1 gene polymorphisms on chicken meat tenderness. Genet. Mol. Res. 2015, 14, 1393–1403. [Google Scholar]
- Piórkowska, K.; Żukowski, K.; Nowak, J.; Połtowicz, K.; Ropka-Molik, K.; Gurgul, A. Genome-wide RNA-Seq analysis of breast muscles of two broiler chicken groups differing in shear force. Anim. Genet. 2016, 47, 68–80. [Google Scholar]
- Malone, J.H.; Oliver, B. Microarrays, deep sequencing and the true measure of the transcriptome. BMC Biol. 2011, 9, 1–9. [Google Scholar]
- Li, W.; Zhang, X.; Du, J.; Li, Y.; Chen, Y.; Cao, Y. RNA-seq-based quantitative transcriptome analysis of meat color and taste from chickens administered by eucalyptus leaf polyphenols extract. J. Food Sci. 2020, 85, 1319–1327. [Google Scholar]
- Li, D.; Wang, X.; Fu, Y.; Zhang, C.; Cao, Y.; Wang, J.; Zhang, Y.; Li, Y.; Chen, Y.; Li, Z.; et al. Transcriptome analysis of the breast muscle of Xichuan black-bone chickens under tyrosine supplementation revealed the mechanism of tyrosine-induced melanin deposition. Front. Genet. 2019, 10, 457. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar]
- Dennis, G.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for annotation, visualization, and integrated discovery. Genome Biol. 2003, 4, 1–11. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar]
- Cong, J.; Zhang, L.; Li, J.; Wang, S.; Gao, F.; Zhou, G. Effects of dietary supplementation with carnosine on growth performance, meat quality, antioxidant capacity and muscle fiber characteristics in broiler chickens. J. Sci. Food Agric. 2017, 97, 3733–3741. [Google Scholar]
- Kralik, G.; Sak-Bosnar, M.; Kralik, Z.; Galović, O. Effect of β-alanine dietary supplementation on concentration of carnosine and quality of broiler muscle tissue. J. Poult. Sci. 2014, 51, 151–156. [Google Scholar]
- Tian, Y.; Xie, M.; Wang, W.; Wu, H.; Fu, Z.; Lin, L. Determination of carnosine in Black-Bone Silky Fowl (Gallus gallus domesticus Brisson) and common chicken by HPLC. Eur. Food Res. Technol. 2007, 226, 311–314. [Google Scholar]
- Lengkidworraphiphat, P.; Wongpoomchai, R.; Taya, S.; Jaturasitha, S. Effect of genotypes on macronutrients and antioxidant capacity of chicken breast meat. Asian-Australas. J. Anim. Sci. 2020, 33, 1817–1823. [Google Scholar]
- Mahootchi, E.; Homaei, S.C.; Kleppe, R.; Winge, I.; Hegvik, T.A.; Megias-Perez, R.; Totland, C.; Mogavero, F.; Baumann, A.; Glennon, J.C.; et al. GADL1 is a multifunctional decarboxylase with tissue-specific roles in β-alanine and carnosine production. Sci. Adv. 2020, 6, eabb3713. [Google Scholar]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How muscle structure and composition influence meat and flesh quality. Sci. World J. 2016, 2016, 1–14. [Google Scholar]
- Pette, D.; Staron, R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000, 50, 500–509. [Google Scholar]
- Muniz, M.M.M.; Fonseca, L.F.S.; dos Santos Silva, D.B.; de Oliveira, H.R.; Baldi, F.; Chardulo, A.L.; Ferro, J.A.; Cánovas, A.; de Albuquerque, L.G. Identification of novel mRNA isoforms associated with meat tenderness using RNA sequencing data in beef cattle. Meat Sci. 2021, 173, 108378. [Google Scholar]
- Vaughan, K.T.; Weber, F.E.; Einheber, S.; Fischman, D.A. Molecular cloning of chicken myosin-binding protein (MyBP) H (86-kDa protein) reveals extensive homology with MyBP-C (C-protein) with conserved immunoglobulin C2 and fibronectin type III motifs. J. Biol. Chem. 1993, 268, 3670–3676. [Google Scholar]
- Liu, A.; Nishimura, T.; Takahasi, K. Relationship between structural properties of intramuscular connective tissue and toughness of various chicken skeletal muscles. Meat Sci. 1996, 43, 43–49. [Google Scholar]
- Ito, S.; Nagata, K. Biology of Hsp47 (Serpin H1), a collagen-specific molecular chaperone. Semin. Cell Dev. Biol. 2017, 62, 142–151. [Google Scholar]
- Zhang, Y.Y.; Zan, L.S.; Wang, H.B. Screening candidate genes related to tenderness trait in Qinchuan cattle by genome array. Mol. Biol. Rep. 2011, 38, 2007–2014. [Google Scholar]
- Cui, H.X.; Liu, R.R.; Zhao, G.P.; Zheng, M.Q.; Chen, J.L.; Wen, J. Identification of differentially expressed genes and pathways for intramuscular fat deposition in pectoralis major tissues of fast- and slow-growing chickens. BMC Genom. 2012, 13, 213. [Google Scholar]
- Wen, Y.; Liu, H.; Liu, K.; Cao, H.; Mao, H.; Dong, X.; Yin, Z. Analysis of the physical meat quality in partridge (Alectoris chukar) and its relationship with intramuscular fat. Poult. Sci. 2020, 99, 1225–1231. [Google Scholar]
- Yang, S.H.; He, C.S.; Li, C.H.; Liu, G.Q. RNA-Seq reveals differentially expressed genes and pathways affecting intramuscular fat metabolism in Huangshan black chicken population. J. Agric. Sci. 2020, 12, 117–127. [Google Scholar]
- Girolami, A.; Marsico, I.; D'Andrea, G.; Braghieri, A.; Napolitano, F.; Cifuni, G.F. Fatty acid profile, cholesterol content and tenderness of ostrich meat as influenced by age at slaughter and muscle type. Meat Sci. 2003, 64, 309–315. [Google Scholar]
- Lian, T.; Wang, L.; Liu, Y. A new insight into the role of calpains in post-mortem meat tenderization in domestic animals: A review. Asian-Australas. J. Anim. Sci. 2013, 26, 443–454. [Google Scholar]
- Okumura, F.; Shimogiri, T.; Shinbo, Y.; Yoshizawa, K.; Kawabe, K.; Mannen, H.; Okamoto, S.; Cheng, H.H.; Maeda, Y. Linkage mapping of four chicken calpain genes. Anim. Sci. J. 2005, 76, 121–127. [Google Scholar]
- Rasouli, Z.; Zerehdaran, S.; Azari, M.A.; Shargh, M.S. Genetic polymorphism of the CAPN1 gene is associated with meat quality traits in Japanese quail. Br. Poult. Sci. 2013, 54, 171–175. [Google Scholar]
- Piórkowska, K.; Nowak, J.; Połtowicz, K. The normalisation of CAPN gene expression in M. pectoralis superficialis in broiler lines differing in growth rate and their relationship to breast muscle tenderness. Br. Poult. Sci. 2015, 56, 452–458. [Google Scholar]
- Sorimachi, H.; Hata, S.; Ono, Y. Impact of genetic insights into calpain biology. J. Biochem. 2011, 150, 23–37. [Google Scholar]
- Fernández-Barroso, M.Á.; Silió, L.; Rodríguez, C.; Palma-Granados, P.; López, A.; Caraballo, C.; Sánchez-Esquiliche, F.; Gómez-Carballar, F.; García-Casco, J.M.; Muñoz, M. Genetic parameter estimation and gene association analyses for meat quality traits in open-air free-range Iberian pigs. J. Anim. Breed Genet. 2020, 137, 581–598. [Google Scholar]
- El-Senousey, H.K.; Fouad, A.M.; Yao, J.H.; Zhang, Z.G.; Shen, Q.W. Dietary alpha lipoic acid improves body composition, meat quality and decreases collagen content in muscle of broiler chickens. Asian-Australas. J. Anim. Sci. 2013, 26, 394–400. [Google Scholar]
- Katemala, S.; Molee, A.; Thumanu, K.; Yongsawatdigul, J. Meat quality and Raman spectroscopic characterization of Korat hybrid chicken obtained from various rearing periods. Poult. Sci. 2021, 100, 1248–1261. [Google Scholar]
- Beauclercq, S.; Hennequet-Antier, C.; Praud, C.; Godet, E.; Collin, A.; Tesseraud, S.; Metayer-Coustard, S.; Bourin, M.; Moroldo, M.; Martins, F.; et al. Muscle transcriptome analysis reveals molecular pathways and biomarkers involved in extreme ultimate pH and meat defect occurrence in chicken. Sci. Rep. 2017, 7, 6447. [Google Scholar]
- Teltathum, T.; Mekchay, S. Relationships between Pectoralis muscle proteomes and shear force in Thai indigenous chicken meat. Agric. Nat. Resour. 2010, 44, 53–60. [Google Scholar]
- Doherty, M.K.; McLean, L.; Hayter, J.R.; Pratt, J.M.; Robertson, D.H.L.; EI-Shafei, A.; Gaskell, S.J.; Beynon, R.J. The proteome of chicken skeletal muscle: Changes in soluble protein expression during a growth in layer strain. Proteomics 2004, 4, 2082–2093. [Google Scholar]
- Hausman, G.J.; Dodson, M.V.; Ajuwon, K.; Azain, M.; Barnes, K.M.; Guan, L.L.; Jiang, Z.; Poulos, S.P.; Sainz, R.D.; Smith, S.; et al. Board-invited review: The biology and regulation of preadipocyte and adipocytes in meat animals. J. Anim. Sci. 2009, 87, 1218–1246. [Google Scholar]
- Dimitriadis, G.; Mitrou, P.; Lambadiari, V.; Maratou, E.; Raptis, S.A. Insulin effects in muscle and adipose tissue. Diabetes Res. Clin. Pract. 2011, 93, S52–S59. [Google Scholar]
- Zhang, M.; Zheng, D.; Peng, Z.; Zhu, Y.; Li, R.; Wu, Q.; Li, Y.; Li, H.; Xu, W.; Zhang, M.; et al. Identification of differentially expressed genes and lipid metabolism signaling pathways between muscle and fat tissues in broiler chickens. J. Poult. Sci. 2021, 58, 131–137. [Google Scholar]
- Huang, H.; Tindall, D.J. Dynamic FoxO transcription factors. J. Cell Sci. 2007, 120, 2479–2487. [Google Scholar]
- Eijkelenboom, A.; Burgering, B.M.T. FOXOs: Signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 2013, 14, 83–97. [Google Scholar]
- Xie, L.; Luo, C.; Zhang, C.; Zhang, R.; Tang, J.; Nie, Q.; Ma, L.; Hu, X.; Li, N.; Da, Y.; et al. Genome-wide association study identified a narrow chromosome 1 region associated with chicken growth traits. PLoS ONE 2012, 7, e30910. [Google Scholar]
- Chen, B.; Xu, J.; He, X.; Xu, H.; Li, G.; Du, H.; Nie, Q.; Zhang, X. A genome-wide mRNA screen and functional analysis reveal FOXO3 as a candidate gene for chicken growth. PLoS ONE 2015, 10, e0137087. [Google Scholar]
- Willson, N.L.; Forder, R.E.A.; Tearle, R.; Williams, J.L.; Hughes, R.J.; Nattrass, G.S.; Hynd, P.I. Transcriptional analysis of liver from chickens with fast (meat bird), moderate (F1 layer x meat bird cross) and low (layer bird) growth potential. BMC Genom. 2018, 19, 309. [Google Scholar]
- Li, Y.; Chen, Y.; Jin, W.; Fu, S.; Li, D.; Zhang, Y.; Sun, G.; Jiang, R.; Han, R.; Li, Z.; et al. Analyses of microRNA and mRNA expression profiles reveal the crucial interaction networks and pathways for regulation of chicken breast muscle development. Front. Genet. 2019, 10, 197. [Google Scholar]
- Sanchez, A.M.J.; Candau, R.B.; Bernardi, H. FoxO transcription factors: Their roles in the maintenance of skeletal muscle homeostasis. Cell Mol. Life Sci. 2014, 71, 1657–1671. [Google Scholar]
- Felício, A.M.; Gaya, L.G.; Ferraz, J.B.S.; Moncau, C.T.; Mattos, E.C.; Santos, N.P.; Filho, T.M.; Balieiro, J.C.C.; Eler, J.P. Heritability and genetic correlation estimates for performance, meat quality and quantitative skeletal muscle fiber traits in broiler. Livest. Sci. 2013, 157, 81–87. [Google Scholar]
- Kubota, S.; Vandee, A.; Keawnakient, P.; Molee, W.; Yongsawatdigul, J.; Molee, A. Effects of the MC4R, CAPN1, and ADSL genes on body weight and purine content in slow-growing chickens. Poult. Sci. 2019, 98, 4327–4337. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sample ID 1 | Raw Reads | Clean Reads | Q30 (%) 2 | GC Content (%) | Total Mapped Read | Uniquely Mapped Reads | Multiple Mapped Reads | Mapping Rage (%) |
|---|---|---|---|---|---|---|---|---|
| A1 | 46,462,620 | 46,288,554 | 94.94 | 55.01 | 37,652,127 | 33,451,212 | 4,200,915 | 81.34 |
| A2 | 47,166,518 | 47,013,356 | 95.04 | 54.01 | 39,871,968 | 35,898,964 | 3,973,004 | 84.81 |
| A3 | 47,881,994 | 47,713,850 | 94.31 | 56.61 | 36,339,766 | 32,824,893 | 3,514,873 | 76.16 |
| A4 | 43,994,624 | 43,820,892 | 94.49 | 56.25 | 33,585,942 | 28,618,863 | 4,967,079 | 76.64 |
| A5 | 48,893,704 | 48,746,180 | 95.02 | 54.87 | 41,053,319 | 35,969,263 | 5,084,056 | 84.21 |
| B1 | 46,163,116 | 46,008,446 | 94.76 | 54.55 | 38,295,423 | 34,456,408 | 3,839,015 | 83.24 |
| B2 | 48,282,906 | 48,121,860 | 94.81 | 55.03 | 40,616,995 | 35,710,664 | 4,906,331 | 84.4 |
| B3 | 47,914,044 | 47,776,800 | 94.63 | 55.36 | 39,450,436 | 36,438,554 | 3,011,882 | 82.57 |
| B4 | 47,830,614 | 47,678,704 | 94.66 | 54.1 | 39,195,229 | 35,730,602 | 3,464,627 | 82.21 |
| B5 | 46,867,322 | 46,705,710 | 94.61 | 54.5 | 38,611,389 | 35,737,411 | 2,873,978 | 82.67 |
| C1 | 46,346,778 | 46,172,558 | 94.8 | 53.96 | 38,610,455 | 35,584,466 | 3,025,989 | 83.62 |
| C2 | 46,252,972 | 46,087,264 | 94.89 | 53.78 | 39,318,314 | 36,417,362 | 2,900,952 | 85.31 |
| C3 | 47,149,780 | 47,007,510 | 94.36 | 54.77 | 38,372,185 | 34,911,827 | 3,460,358 | 81.63 |
| C4 | 48,618,940 | 48,452,690 | 95.0 | 54.46 | 40,454,304 | 36,297,707 | 4,156,597 | 83.49 |
| C5 | 41,721,980 | 41,582,684 | 94.67 | 53.52 | 35,046,741 | 32,462,385 | 2,584,356 | 84.28 |
| Average | 46,769,860.8 | 46,611,803.8 | 94.73 | 54.71 | 38,431,639.5 | 34,700,705.4 | 3,730,934.1 | 82.43 |
| Category | Term ID | Term | Count | p-Value | Genes 1 |
|---|---|---|---|---|---|
| Biological process | GO:0003151 | Outflow tract morphogenesis | 4 | 6.75 × 10−4 | BMP4↑, HEYL↓, DHRS3↑, SFRP2↓ |
| GO:0007160 | Cell-matrix adhesion | 3 | 0.031 | OTOA↑, TSC1↑, ITGB1BP1↑ | |
| GO:0002043 | Blood vessel endothelial cell proliferation involved in sprouting angiogenesis | 2 | 0.034 | BMP4↑, ITGB1BP1↑ | |
| Cellular component | GO:0009986 | Cell surface | 8 | 7.13 × 10−4 | ADAMTS15↓, RTN4RL1↓, SDC1↓, CFTR↑, VASN↓, PLA2R1↑, OTOA↑, THBD↓ |
| GO:0005615 | Extracellular space | 12 | 0.001 | PXDNL↓, ADAMTS15↓, TST↓, BMP4↑, PLA2G15↑, VASN↓, TNFSF10↑, CPN1↓, SERPINH1↓, SFRP2↓, PPFIBP2↑, CTSV↑ | |
| GO:0016324 | Apical plasma membrane | 4 | 0.029 | CNTFR↓, CFTR↑, OTOA↑, AMOTL1↑ |
| Category | Term ID | Term | Count | p-Value | Genes 1 |
|---|---|---|---|---|---|
| Biological process | GO:0035556 | Intracellular signal Transduction | 10 | 5.68 × 10−4 | ASB8↓, ASB2↑, SGK1↑, PRKAA2↓, RGS9↑, ASB5↑, NUAK1↑, DAPK1↑, SPSB1↑, ARHGEF7↑ |
| GO:0042752 | Regulation of circadian rhythm | 3 | 0.025 | PRKAA2↓, NOCT↓, PPARA↑ | |
| GO:0006094 | Gluconeogenesis | 3 | 0.032 | PFKFB4↓, PGAM1↓, CRY1↑ | |
| GO:0010629 | Negative regulation of gene expression | 4 | 0.038 | CDKN1A↓, CTGF↑, TIPARP↓, NOCT↓ | |
| GO:0009267 | Cellular response to Starvation | 3 | 0.039 | PRKAA2↓, MYOD1↑, PIK3C2B↓ | |
| GO:0042060 | Wound healing | 3 | 0.039 | PECAM1↑, PPARA↑, SLC11A1↓ | |
| GO:0032922 | Circadian regulation of gene expression | 3 | 0.045 | NOCT↓, PPARA↑, CRY1↑ | |
| Molecular function | GO:0004722 | Protein serine/threonine phosphatase activity | 3 | 0.029 | PPM1K↓, PPM1J↓, PDP1↓ |
| Term | Count | p-Value | Genes 1 |
|---|---|---|---|
| β-alanine supplementation | |||
| gga04068: FoxO signaling pathway | 5 | 0.008 | FOXO1↑, CCNG2↑, TNFSF10↑, G6PC3↓, IRS2↑ |
| gga04910: Insulin signaling pathway | 4 | 0.044 | FOXO1↑, G6PC3↓, IRS2↑, TSC1↑ |
| L-histidine supplementation | |||
| gga04068: FoxO signaling pathway | 6 | 0.008 | CDKN1A↓, SGK1↑, PRKAA2↓, FOXO1↑, TNFSF10↑, IRS2↑ |
| gga04931: Insulin resistance | 5 | 0.018 | PRKAA2↓, FOXO1↑, PPARA↑, CREB5↑, IRS2↑ |
| gga04920: Adipocytokine signaling pathway | 4 | 0.029 | ACSBG1↑, PRKAA2↓, PPARA↑, IRS2↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubota, S.; Promkhun, K.; Sinpru, P.; Suwanvichanee, C.; Molee, W.; Molee, A. RNA Profiles of the Korat Chicken Breast Muscle with Increased Carnosine Content Produced through Dietary Supplementation with β-Alanine or L-Histidine. Animals 2021, 11, 2596. https://doi.org/10.3390/ani11092596
Kubota S, Promkhun K, Sinpru P, Suwanvichanee C, Molee W, Molee A. RNA Profiles of the Korat Chicken Breast Muscle with Increased Carnosine Content Produced through Dietary Supplementation with β-Alanine or L-Histidine. Animals. 2021; 11(9):2596. https://doi.org/10.3390/ani11092596
Chicago/Turabian StyleKubota, Satoshi, Kasarat Promkhun, Panpradub Sinpru, Chanadda Suwanvichanee, Wittawat Molee, and Amonrat Molee. 2021. "RNA Profiles of the Korat Chicken Breast Muscle with Increased Carnosine Content Produced through Dietary Supplementation with β-Alanine or L-Histidine" Animals 11, no. 9: 2596. https://doi.org/10.3390/ani11092596
APA StyleKubota, S., Promkhun, K., Sinpru, P., Suwanvichanee, C., Molee, W., & Molee, A. (2021). RNA Profiles of the Korat Chicken Breast Muscle with Increased Carnosine Content Produced through Dietary Supplementation with β-Alanine or L-Histidine. Animals, 11(9), 2596. https://doi.org/10.3390/ani11092596