
Metabolomic Analysis of the Takifugu Obscurus Gill under Acute Hypoxic Stress

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Instruments and Reagents
2.3. Experimental Fish
2.4. Experimental Design and Sample Collection
2.5. Metabolite Extraction
2.6. Data Processing and Metabolite Identification
3. Results
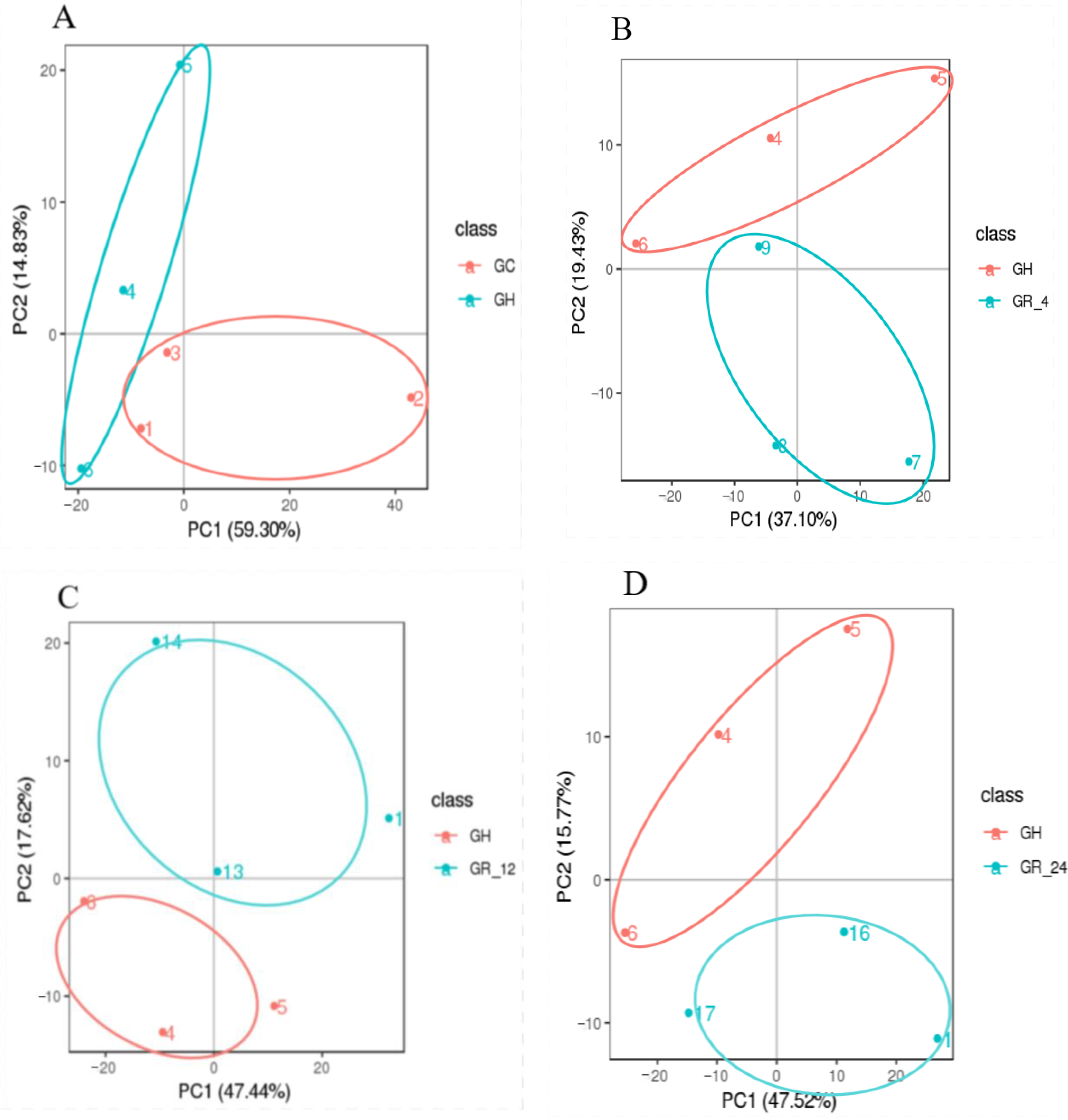
3.1. Data Quality Analyses
3.2. Identification of Differentially Abundant Metabolites
3.3. Metabolic Pathways Responsive to Hypoxic Stress
4. Discussion
4.1. Lipid Metabolism
4.2. Catabolism of Amino Acids
4.3. Purine Metabolism
4.4. Hypoxia-related Signaling Pathways
4.4.1. The FoxO Signaling Pathway
4.4.2. The mTOR Signaling Pathway
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.R. Effects of Hypoxic Stress on Energy Metabolism, Blood Indexes and Gene Expression of Takifugu obscurus; Nanjing Normal University: Nanjing, China, 2018. [Google Scholar]
- Rosenberger, A.E.; Chapman, L.J. Hypoxic wetland tributaries as faunal refugiafrom an introduced predator. Ecol. Freshw. Fish 1999, 8, 22–34. [Google Scholar] [CrossRef]
- Richards, J.G. Physiological, behavioral and biochemical adaptations of intertidal fishes to hypoxia. J. Exp. Biol. 2011, 214, 191–199. [Google Scholar] [CrossRef]
- Xu, H.; Chen, X.M.; Wang, G.Q.; Cong, L.M. Research progress of hypoxia on aquaculture. Feed Ind. 2016, 37, 33–37. [Google Scholar]
- Rasmus, E.; Johansen, L.J.; Rummer, L.J.; Esbaugh, J.A. Effects of hypoxia and ocean acidification on theupper thermal niche boundaries of coral reef fishes. Biol. Lett. 2017, 13, 20170135. [Google Scholar]
- Secor, D.H.; Gunderson, T.E. Effects of hypoxia and temperature on survival, growth, and respiration of juvenile Atlantic sturgeon, Acipenser oxyrinchus. Fish. Bull. 1998, 96, 603–613. [Google Scholar]
- Wu, Y.G.; Lin, F.; Jiang, S.W.; Jia, R.N.; Jiang, C.W.; Xu, Q.H. Analysis of Zebrafish Ovary Transcriptome Characteristics under Long-term Hypoxia Stress. Genom. Appl. Biol. 2021, 40, 1921–1927. [Google Scholar]
- Zhou, B.S. Effects of Hypoxia on Metabolism, Bioenergetics, Growth, and Reproduction of the Common Carp (Cyprinus carpio L.); University of Hong Kong: Hongkong, 2001. [Google Scholar]
- Landry, C.A.; Steele, S.L.; Manning, S.; Cheek, A.O. Long term hypoxia suppresses reproductive capacity in the estuarine fish, Fundulus grandis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 317–323. [Google Scholar] [CrossRef]
- Wang, J.; Li, Z.H.; Chen, Y.F.; Zhou, Y. The combined effect oftemperature and pH on embryonic development of obscure puffer Takifugu obscurus and its ecological implications. Biochem. Syst. Ecol. 2015, 58, 1–6. [Google Scholar] [CrossRef]
- Cappello, T.; Maisano, M.; Giannetto, A.; Parrino, V.; Fasulo, S. Neurotoxicological effects on marine mussel Mytilus galloprovincialis caged at petro-chemical contaminated areas (eastern Sicily, Italy): 1H NMR and immunohistochemical assays. Comp. Biochem. Physiol. C 2015, 169, 7–15. [Google Scholar]
- Li, X.R.; Wang, T.; Yin, S.W.; Zhang, G.S.; Cao, Q.Q.; Wen, X.; Zhang, H.Y.; Wang, D.; Zhu, W.X. The improved energy metabolism and blood oxygen-carrying capacity for pufferfish, Takifugu fasciatus, against acute hypoxia under the regulation of oxygen sensors. Fish Physiol. Biochem. 2019, 45, 323–340. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, L.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2012, 8, 17–32. [Google Scholar] [CrossRef]
- Kyeong, D.; Kim, J.; Shin, Y.; Subramaniyam, S.; Kang, B.C.; Shin, E.H.; Park, E.H.; Noh, E.S.; Kim, Y.O.; Park, J.Y. Expression of heat shock proteins in thermally challenged Pacific abalone Haliotis discus hannai. Genes 2020, 11, 22. [Google Scholar] [CrossRef]
- Toprak, U. The role of peptide hormones in insect lipid metabolism. Front. Physiol. 2020, 11, 434. [Google Scholar] [CrossRef]
- McGany, J.D.; Brown, N.F. The miochondrial palmitoyiransferase system-From concept to molecular analysis. Eur. J. Biochem. 1997, 244, 1–14. [Google Scholar]
- Yin, J.; Gao, Z.G.; He, Q.; Zhou, D.Q.; Guo, Z.K.; Ye, J.P. Role of hypoxia in obesity-induced disorders of glucose and lipid metabolism in adipose tissue. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E333–E342. [Google Scholar] [CrossRef]
- Mao, S.Z.; Gao, Y.Q.; Chen, J.; Liu, F.Y.; Gao, W.X.; Huang, J.; Liao, W.G.; Cai, M.C. Characteristics of fat metabolism in skeletal musclec of rats after hypobaric hypoxic acclimation. J. Med. Coll. PLA 2008, 5, 258–262. [Google Scholar]
- Zhang, K. Effects of CLA and Hypoxia on Skeletal Muscle Lipid Metabolism in Rats; Hangzhou Normal University: Hangzhou, China, 2013. [Google Scholar]
- Imbrogno, S.; Aiello, D.; Filice, M.; Leo, S.; Mazza, R.; Cerra, M.C.; Napoli, A. MS-based proteomic analysis of cardiac response to hypoxia in the goldfish (Carassius auratus). Sci. Rep. 2019, 9, 18953. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.Q.; Wang, J.; Liao, M.L.; Li, X.X.; Dong, Y.W. Temperature adaptations of the thermophilic snail Echinolittorina malaccana: Insights from metabolomic analysis. J. Exp. Biol. 2021, 224, jeb238659. [Google Scholar] [CrossRef]
- Farine, L.; Niemann, M.; Schneider, A.; Bütikofer, P. Phosphatidylethanolamine and phosphatidylcholine biosynthesis by the Kennedy pathway occurs at different sites in Trypanosoma brucei. Sci. Rep. 2015, 5, 16787. [Google Scholar] [CrossRef]
- Mukodani, J.; Ishikawa, Y.; Fukuzaki, H. Effects of hypoxia on sterolsynthesis, acyl-CoA: Cholesterol acyltransferase activity, and eflux ofcholesterol in cultured rabbit skin fibroblasts. Arteriosclerosis 1990, 10, 106–110. [Google Scholar] [CrossRef]
- Strasser, B.; Sperner-Unterweger, B.; Fuchs, D.; Gostner, J.M. Mechanisms of Inflammation-Associated Depression: Immune Influences on Tryptophan and Phenylalanine Metabolisms. Curr. Top. Behav. Neurosci. 2016, 31, 95–115. [Google Scholar]
- Yao, M.Z.; Liu, Y.J.; Li, S.W.; Chen, Z.X.; Wang, P.; Sun, Y.C. Metabolomics of gill target organs of Founder’s silver crucian carp under NaHCO3 habitat stress. Chin. Aquat. Sci. 2022, 1–12. [Google Scholar]
- Nemkov, T.; Sun, K.; Reisz, J.A.; Song, A.; Yoshida, T.; Dunham, A.; Wither, M.J.; Francis, R.O.; Roach, R.C.; Dzieciatkowska, M.; et al. Hypoxia modulates the purine salvage pathway and decreases red blood cell and supernatant levels of hypoxanthine during refrigerated storage. Haematologica 2018, 103, 361–372. [Google Scholar] [CrossRef]
- Mustafa, S.A.; Karieb, S.S.; Davies, S.J.; Jha, A.N. Assessment of oxidative damage to DNA, transcriptional expression of key genes, lipid peroxidation and histopathological changes in carp Cyprinus carpio L. following exposure to chronic hypoxic and subsequent recovery in normoxic conditions. Mutagenesis 2015, 30, 107–116. [Google Scholar] [CrossRef]
- Martinez, D.L.; Tsuchiya, Y.; Gout, I. Coenzyme A biosynthetic machinery in mammalian cells. Biochem. Soc. Trans. 2014, 42, 1112–1117. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquaticanimals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, J.Q.; Quan, J.Q.; Wu, S.J.; Wang, X.L.; Pan, Y.C. Effects of acute hypoxic stress and reoxygenation on oxidative stress in crucian carp. Aquac. Sci. 2020, 39, 649–656. [Google Scholar]
- Eijkelenboom, A.; Burgering, B.M. FOXOs: Signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 2013, 14, 83–97. [Google Scholar] [CrossRef]
- Lehtinen, M.K.; Yuan, Z.; Boag, P.R.; Yang, Y.; Villén, J.; Becker, E.; Dibacco, S.; Iglesia, N.; Gygi, S.; Blackwell, T.K. A conserved MST-FOXO signaling pathway mediates oxidative-stress responses and extends life span. Cell 2006, 125, 987–1001. [Google Scholar] [CrossRef]
- Putker, M.; Madl, T.; Vos, H.R.; Ruiter, H.D.; Visscher, M.; Berg, V.D.; Kaplan, M.; Korswagen, H.C.; Boelens, R.; Vermeulen, M.; et al. Redox-dependent control of FOXO/DAF-16 by transportin-1. Mol. Cell 2013, 49, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.D.; Wang, Z.H.; Yan, B. Strategies for hypoxia adaptation in fish species: A review. J. Comp. Physiol. B 2013, 183, 1005–1013. [Google Scholar] [CrossRef]
- Brugge, J.; Hung, M.C.; Mills, G.B. A new mutational AKTivation in the PI3K pathway. Cancer Cell 2007, 12, 104–107. [Google Scholar] [CrossRef]
- Faber, A.C.; Coffee, E.M.; Costa, C.; Dastur, A.; Ebi, H.; Hata, A.N.; Yeo, A.T.; Edelman, E.J.; Song, Y.; Tam, A.T.; et al. mTOR inhibition specifically sensitizes colorectal cancers with KRAS or BRAF mutations to BCL-2/BCL-XL inhibition by suppressing MCL-1. Cancer Discov. 2014, 4, 42–52. [Google Scholar] [CrossRef]
- Ossum, C.G.; Wulff, T.; Hoffmann, E.K. Regulation of themitogen-activated protein kinase p44 ERK activity during anoxia/recovery in rainbow trout hypodermal fibroblasts. J. Exp. Biol. 2006, 209, 1765–1776. [Google Scholar] [CrossRef]
- Liu, H.L.; Meng, F.X. Effects and mechanisms of oxidative stress on atresia of luminal follicles in animals. J. Nanjing Agric. Univ. 2019, 42, 6–13. [Google Scholar]
- Shao, D.; Zhai, P.Y.; Delre, D.P.; Sciarretta, S.; Yabuta, N.; Nojima, H.; Lim, D.; Pan, D.J.; Sadoshima, J. A functional interaction between Hippo-YAP signalling and FoxO1 mediates the oxidative stress response. Nat. Commun. 2014, 5, 3315. [Google Scholar] [CrossRef]
- Qi, D.L.; Chao, Y.; Wu, R.R.; Xia, M.Z.; Chen, Q.C.; Zheng, Z.Q. Transcriptome analysis provides insights into the adaptive responses to hypoxia of a schizothoracine Fish (Gymnocypris eckloni). Front. Physiol. 2018, 9, 1326. [Google Scholar] [CrossRef]
- Jibb, L.A.; Richards, J.G. AMP-activated protein kinase activity during metabolic rate depression in the hypoxic goldfish, Carassius auratus. J. Exp. Biol. 2008, 211, 3111–3122. [Google Scholar] [CrossRef]
- Williams, K.J.; Cassidy, A.A.; Verhille, C.E.; Lamarre, S.G.; MacCormack, T.J. Diel cycling hypoxia enhances hypoxia-tolerance in rainbow trout (Oncorhynchus mykiss): Evidence of physiological and metabolic plasticity. J. Exp. Biol. 2019, 222, jeb206045. [Google Scholar] [CrossRef]
- Lizcano, J.M.; Gèoransson, O.; Toth, R.; Deak, M.; Morrice, N.A.; Boudeau, J.; Hawley, S.A.; Udd, L.; Mäkelä, T.P.; Hardie, D.G.; et al. LKB1 is a maste kinase that activates 13 kinases of the AMPK subfamily, including MARK/PAR-1. EMBO J. 2004, 23, 833–843. [Google Scholar] [CrossRef]
- Brugarolas, J.; Lei, K.; Hurley, R.L.; Manning, B.D.; Reiling, J.H.; Hafen, E.; Witters, L.A.; Ellisen, L.W.; Kaelin, W.G. Regulation of mTOR function inresponse to hypoxia by REDD1 and the TSC1/TSC2 tumor suppressor complex. Genes 2004, 18, 2893–2904. [Google Scholar] [CrossRef] [Green Version]
- Sofer, A.; Lei, K.; Johannessen, C.M.; Ellisen, L.W. Regulation of mTOR and cell growth in response to energy stress by REDD1. Mol. Cell Biol. 2005, 25, 5834–5845. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 169, 361–371. [Google Scholar] [CrossRef]
- Tian, T.; Li, X.Y.; Zhang, J.H. mTOR Signaling in Cancer and mTOR Inhibitors in Solid Tumor Targeting Therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef]
- Chun, Y.; Kim, J. AMPK-mTOR Signaling and Cellular Adaptations in Hypoxia. Int. J. Mol. Sci. 2021, 22, 9765. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | Differentially Abundant Metabolites | Metabolites More Abundant in GH |
|---|---|---|
| GC vs. GH | 36 | 22 |
| GR_4 vs. GH | 18 | 12 |
| GR_12 vs. GH | 24 | 20 |
| GR_24 vs. GH | 34 | 23 |
| Metabolite | VIP | FC | P | Trend | Metabolic pathway |
|---|---|---|---|---|---|
| Porphobilinogen | 1.61 | 2.92 | 0.049 | ↑ | Porphyrin and chlorophyll metabolism |
| Indole | 1.63 | 1.78 | 0.022 | ↑ | Phenylalanine, tyrosine and tryptophan biosynthesis Tryptophan metabolism |
| 2-Phenylacetamide | 1.59 | 1.76 | 0.027 | ↑ | Phenylalanine metabolism |
| Testosterone | 1.67 | 0.21 | 0.047 | ↑ | Steroid hormone biosynthesis |
| Uric acid | 1.62 | 0.67 | 0.036 | ↑ | Purine metabolism |
| Adenosine 5′-monophosphate | 1.73 | 0.57 | 0.033 | ↑ | FoxO signaling pathway mTOR signaling pathway |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Hu, Z.; Li, R.; Wang, Y.; Zhou, J.; Xu, H.; Wang, G.; Qiu, X.; Wang, X. Metabolomic Analysis of the Takifugu Obscurus Gill under Acute Hypoxic Stress. Animals 2022, 12, 2611. https://doi.org/10.3390/ani12192611
Zhang H, Hu Z, Li R, Wang Y, Zhou J, Xu H, Wang G, Qiu X, Wang X. Metabolomic Analysis of the Takifugu Obscurus Gill under Acute Hypoxic Stress. Animals. 2022; 12(19):2611. https://doi.org/10.3390/ani12192611
Chicago/Turabian StyleZhang, Huakun, Ziwen Hu, Run Li, Yaohui Wang, Jinxu Zhou, Hao Xu, Guan Wang, Xuemei Qiu, and Xiuli Wang. 2022. "Metabolomic Analysis of the Takifugu Obscurus Gill under Acute Hypoxic Stress" Animals 12, no. 19: 2611. https://doi.org/10.3390/ani12192611
APA StyleZhang, H., Hu, Z., Li, R., Wang, Y., Zhou, J., Xu, H., Wang, G., Qiu, X., & Wang, X. (2022). Metabolomic Analysis of the Takifugu Obscurus Gill under Acute Hypoxic Stress. Animals, 12(19), 2611. https://doi.org/10.3390/ani12192611